
Herbaceous and Woody Root Biomass, Seasonal Changes in Root Turnover, and Arbuscular Mycorrhizal and Ectomycorrhizal Colonization during Primary Succession in Post-Mining Sites


Abstract
:1. Introduction
2. Material and Methods
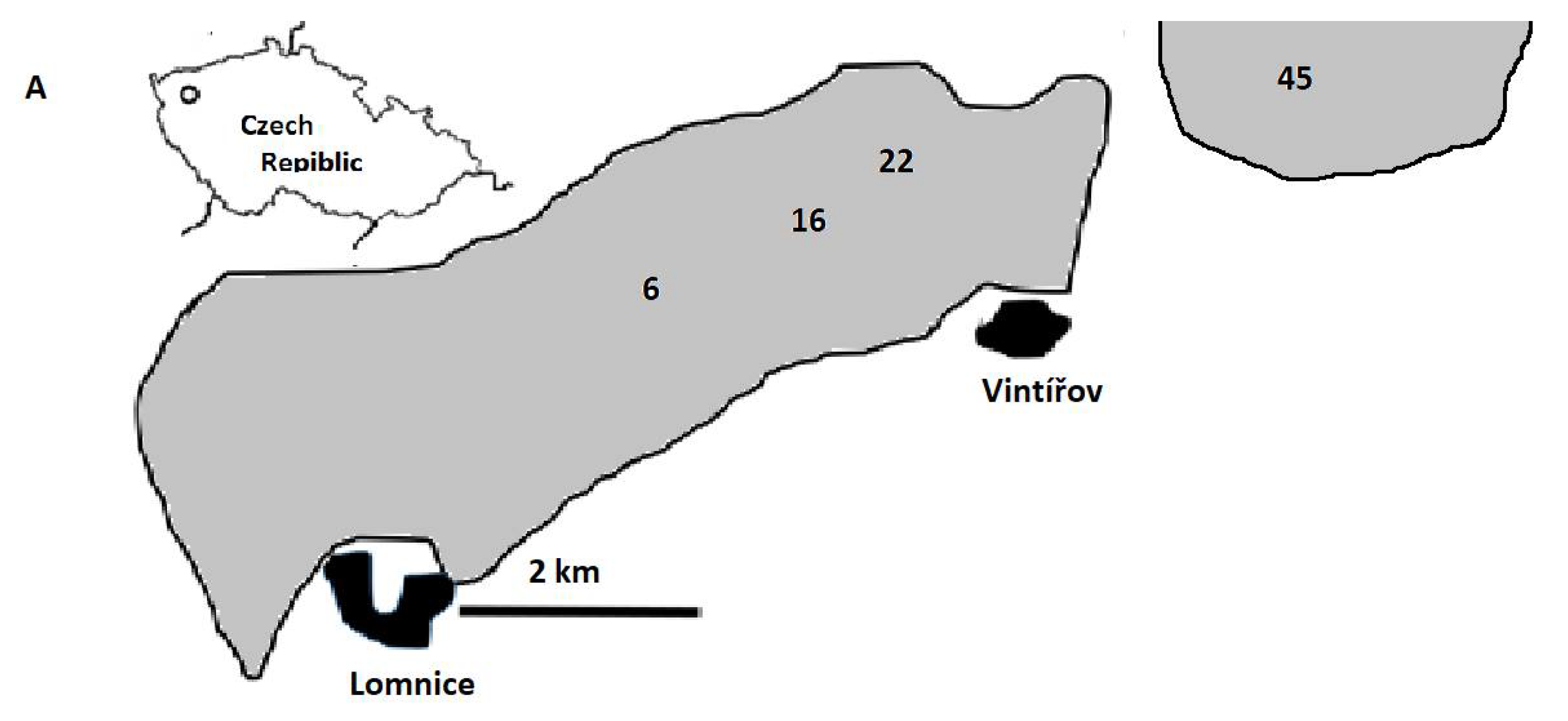
2.1. Study Sites
2.2. Root Sampling and Ingrowth Cores
2.3. Root Processing
2.4. Mycorrhizal Colonization Potential
2.5. Data Evaluation and Statistics
3. Results
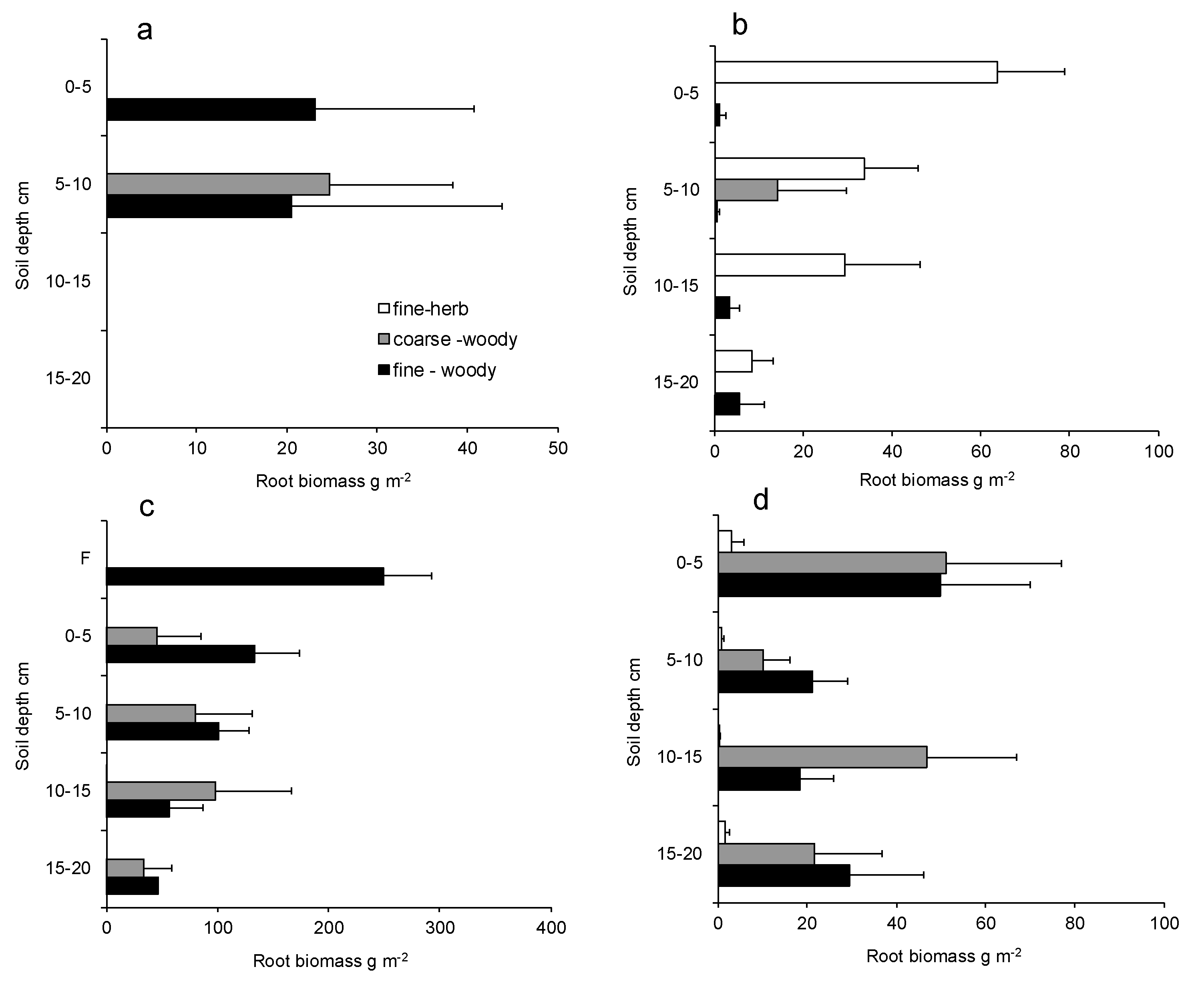
3.1. Depth Distribution of Roots
3.2. Root Biomass and Length Variation among Succession Age and Season
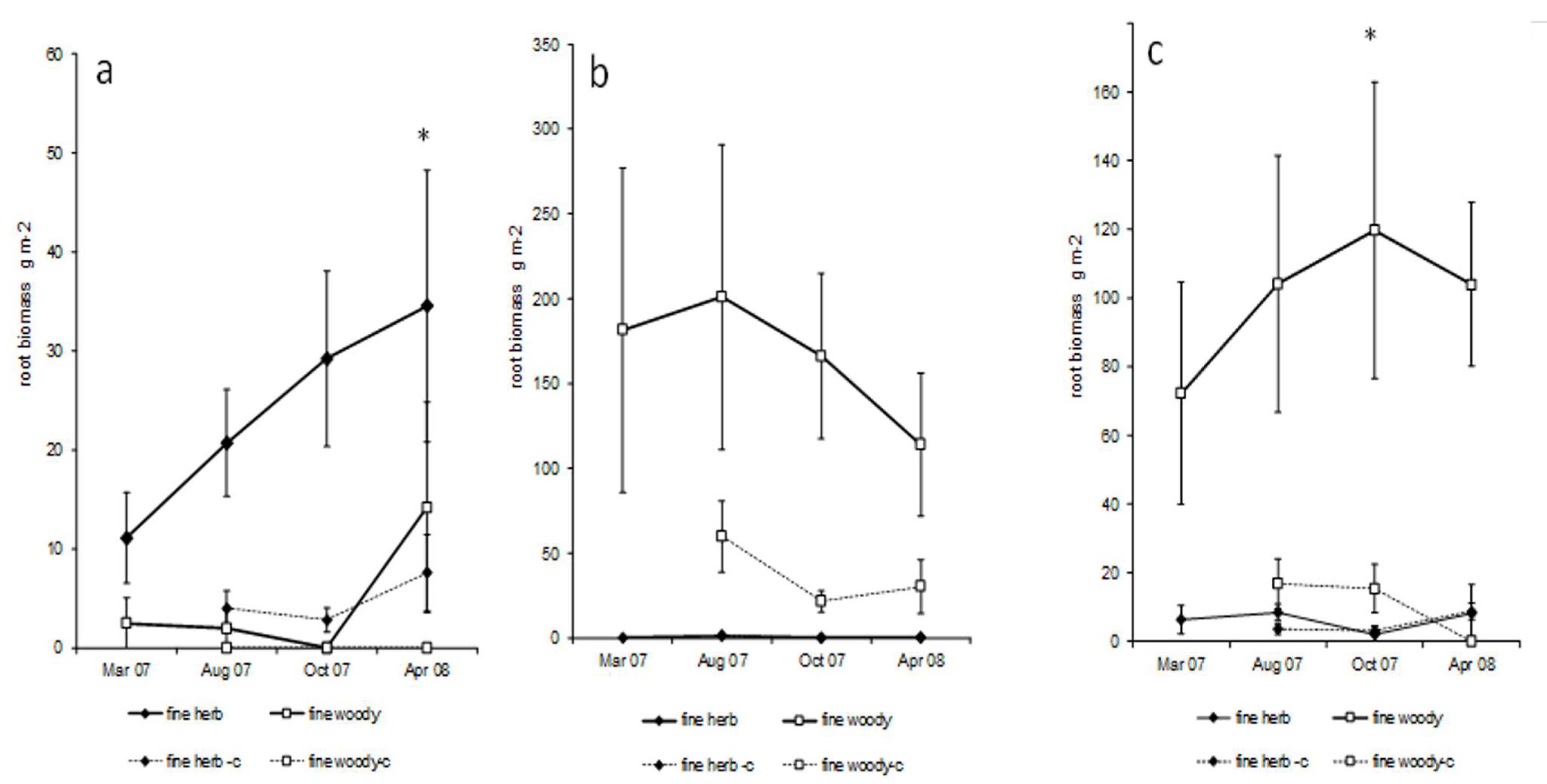
3.3. Ingrowth Cores
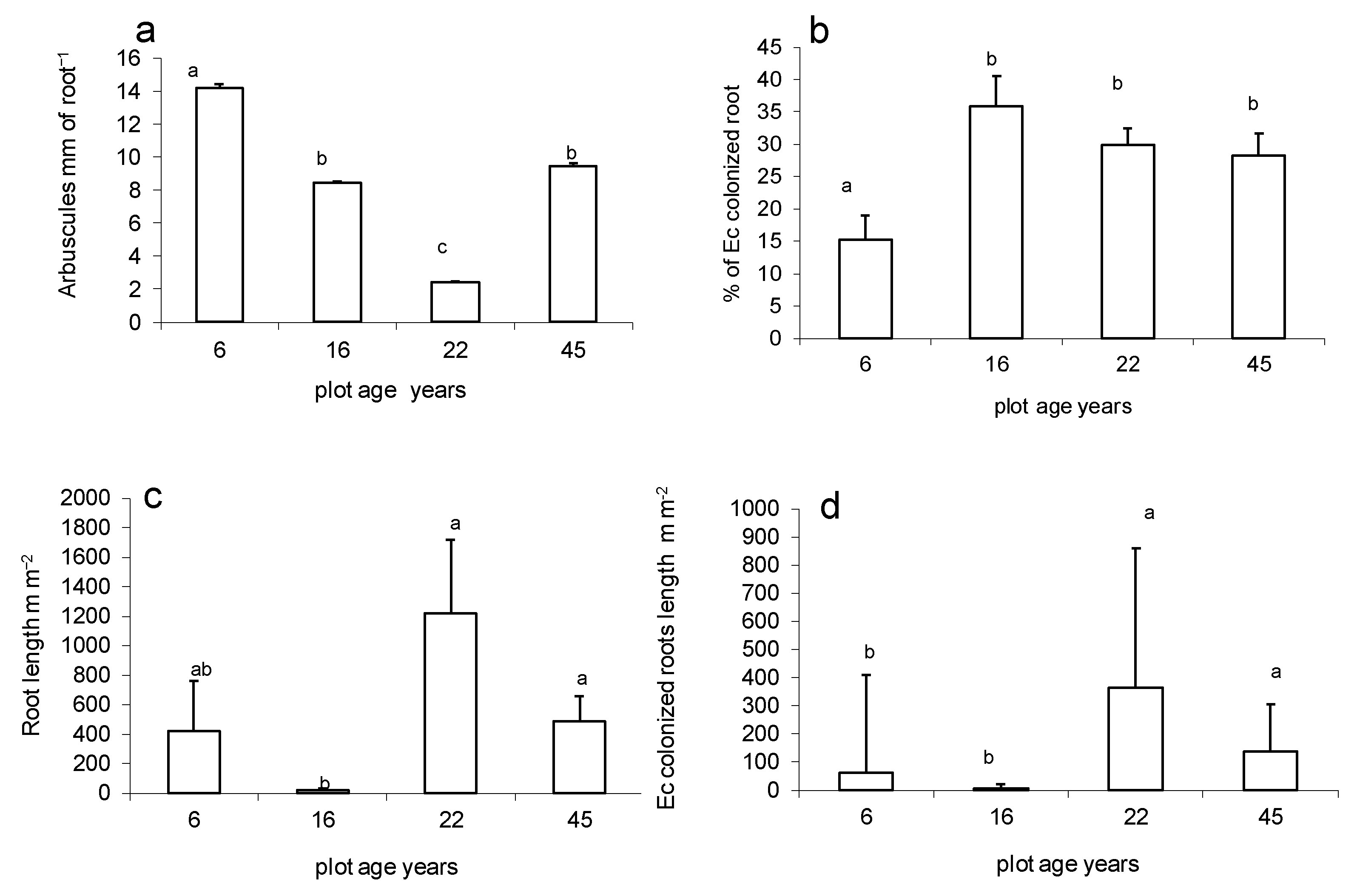
3.4. Mycorrhizal Colonization Potential
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bradshaw, A. Restoration of mined lands—Using natural processes. Ecol. Eng. 1997, 8, 255–269. [Google Scholar] [CrossRef]
- Frouz, J. Soil Biota and Ecosystem Development in Post Mining Sites, 1st ed.; CRC Press: Boca Raton, FL, USA, 2014. [Google Scholar]
- Skousen, J.G.; Johnson, C.D.; Garbutt, K. Natural revegetation of 15 abandoned mine land sites in West Virginia. J. Environ. Qual. 1994, 23, 1224–1230. [Google Scholar] [CrossRef]
- Pietrzykowski, M.; Krzaklewski, W. An assessment of energy efficiency in reclamation to forest. Ecol. Eng. 2007, 30, 341–348. [Google Scholar] [CrossRef]
- Frouz, J.; Prach, K.; Pižl, V.; Háněl, L.; Starý, J.; Tajovský, K.; Materna, J.; Balík, V.; Kalčík, J.; Řehounková, K. Interactions between soil development, vegetation and soil fauna during spontaneous succession in post mining sites. Eur. J. Soil Biol. 2008, 44, 109–121. [Google Scholar] [CrossRef]
- Frouz, J.; Dvorščík, P.; Vávrová, A.; Doušová, O.; Kadochová, Š.; Matějíček, L. Development of canopy cover and woody vegetation biomass on reclaimed and unreclaimed post-mining sites. Ecol. Eng. 2015, 84, 233–239. [Google Scholar] [CrossRef]
- Frouz, J.; Vobořilová, V.; Janoušová, I.; Kadochová, Š.; Matějíček, L. Spontaneous establishment of late successional tree species English oak (Quercus robur) and European beech (Fagus sylvatica) at reclaimed alder plantation and unreclaimed post mining sites. Ecol. Eng. 2015, 77, 1–8. [Google Scholar] [CrossRef]
- Macdonald, S.E.; Landhaeusser, S.M.; Skousen, J.; Franklin, J.; Frouz, J.; Hall, S.; Jacobs, D.F.; Quideau, S. Forest restoration following surface mining disturbance: Challenges and solutions. New For. 2015, 46, 703–732. [Google Scholar] [CrossRef]
- Mudrák, O.; Doležal, J.; Frouz, J. Initial species composition predicts the progress in the spontaneous succession on post-mining sites. Ecol. Eng. 2016, 95, 665–670. [Google Scholar] [CrossRef]
- Prach, K.; Bartha, S.; Joyce, C.B.; Pyšek, P.; Van Diggelen, R.; Wiegleb, G. The role of spontaneous vegetation succession in ecosystem restoration: A perspective. Appl. Veg. Sci. 2001, 4, 111–114. [Google Scholar] [CrossRef]
- Prach, K.; Hobbs, R. Spontaneous succession versus technical reclamation in the restoration of disturbed sites. Restor. Ecol. 2008, 16, 363–366. [Google Scholar] [CrossRef]
- de León, D.G.; Moora, M.; Öpik, M.; Neuenkamp, L.; Gerz, M.; Jairus, T.; Vasar, M.; Bueno, C.G.; Davison, J.; Zobel, M. Symbiont dynamics during ecosystem succession: Co-occurring plant and arbuscular mycorrhizal fungal communities. FEMS Microbiol. Ecol. 2016, 92, fiw097. [Google Scholar] [CrossRef] [PubMed]
- Hüttl, R.F.; Weber, E. Forest ecosystem development in post-mining landscapes: A case study of the Lusatian lignite district. Naturwissenschaften 2001, 88, 322–329. [Google Scholar] [CrossRef] [PubMed]
- Markowicz, A.; Woźniak, G.; Borymski, S.; Piotrowska-Seget, Z.; Chmura, D. Links in the functional diversity between soil microorganisms and plant communities during natural succession in coal mine spoil heaps. Ecol. Res. 2015, 30, 1005–1014. [Google Scholar] [CrossRef]
- Rydlová, J.; Püschel, D.; Dostálová, M.; Janouskova, M.; Frouz, J. Nutrient limitation drives response of Calamagrostis epigejos to arbuscular mycorrhiza in primary succession. Mycorrhiza 2016, 26, 757–767. [Google Scholar] [CrossRef] [PubMed]
- Krüger, C.; Kohout, P.; Janoušková, M.; Püschel, D.; Frouz, J.; Rydlová, J. Plant Communities Rather than Soil Properties Structure Arbuscular Mycorrhizal Fungal Communities along Primary Succession on a Mine Spoil. Front. Microbiol. 2017, 8, 719. [Google Scholar] [CrossRef]
- Holdaway, R.J.; Richardson, S.J.; Dickie, I.A.; Peltzer, D.A.; Coomes, D.A. Species- and community-level patterns in fine root traits along a 120,000-year soil chronosequence in temperate rain forest. J. Ecol. 2011, 99, 954–963. [Google Scholar] [CrossRef]
- Erktan, A.; Roumet, C.; Bouchet, D.; Stokes, A.; Pailler, F.; Munoz, F. Two dimensions define the variation of fine root traits across plant communities under the joint influence of ecological succession and annual mowing. J. Ecol. 2018, 106, 2031–2042. [Google Scholar] [CrossRef]
- Zangaro, W.; De Almeida Alves, R.; De Souza, P.; Rostirola, L.; Lescano, L.; Rondina, A.; Nogueira, M. Succession and environmental variation influence soil exploration potential by fine roots and mycorrhizal fungi in an Atlantic ecosystem in southern Brazil. J. Trop. Ecol. 2014, 30, 237–248. [Google Scholar] [CrossRef]
- Knoblochová, T.; Kohout, P.; Püschel, D.; Doubková, P.; Frouz, J.; Cajthaml, T.; Kukla, J.; Vosátka, M.; Rydlová, J. Asymmetric response of root-associated fungal communities of an arbuscular mycorrhizal grass and an ectomycorrhizal tree to their coexistence in primary succession. Mycorrhiza 2017, 27, 775–789. [Google Scholar] [CrossRef]
- Mudrák, O.; Hermová, M.; Tesnerová, C.; Rydlová, J.; Frouz, J. Above-ground and below-ground competition between the willow Salix caprea and its understorey. J. Veg. Sci. 2015, 27, 156–164. [Google Scholar] [CrossRef]
- Smith, S.E.; Read, D.J.D. Mycorrhizal Symbiosis, 3rd ed.; Academic Press: London, UK, 2008. [Google Scholar]
- Augé, R.M. Water relations, drought and vesicular-arbuscular mycorrhizal symbiosis. Mycorrhiza 2001, 11, 3–42. [Google Scholar] [CrossRef]
- Marulanda, A.; Azcon, R.; Ruiz-Lozano, J.M. Contribution of six arbuscular mycorrhizal fungal isolates to water uptake by Lactuca sativa plants under drought stress. Physiol. Plant. 2003, 119, 526–533. [Google Scholar] [CrossRef]
- Dugassa, G.D.; von Alten, H.; Schönbeck, F. Effects of arbuscular mycorrhiza (AM) on health of Linum usitatissimum L. infected by fungal pathogens. Plant Soil 1996, 185, 173–182. [Google Scholar] [CrossRef]
- Teste, F.P.; Jones, M.D.; Dickie, I.A. Dual-mycorrhizal plants: Their ecology and relevance. New Phytol. 2020, 225, 1835–1851. [Google Scholar] [CrossRef] [PubMed]
- Wang, F. Occurrence of arbuscular mycorrhizal fungi in mining-impacted sites and their contribution to ecological restoration: Mechanisms and applications. Crit. Rev. Environ. Sci. Technol. 2017, 47, 1901–1957. [Google Scholar] [CrossRef]
- Kalucka, I.L.; Jagodzinski, A.M. Successional traits of ectomycorrhizal fungi in forest reclamation after surface mining and agricultural disturbances: A review. Dendrobiology 2016, 76, 91–104. [Google Scholar] [CrossRef]
- Brundrett, M.C.; Abbott, L.K. Roots of Jarrah Forest plants. I. Mycorrhizal associations of shrubs and herbaceous plants. Aust. J. Bot. 1991, 39, 445–457. [Google Scholar] [CrossRef]
- Frouz, J.; Pižl, V.; Tajovský, K. The effect of earthworms and other saprophagous macrofauna on soil microstructure in reclaimed and un-reclaimed post-mining sites in Central Europe. Eur. J. Soil Biol. 2007, 43, 184–189. [Google Scholar] [CrossRef]
- Finzi, A.C.; Abramoff, R.Z.; Spiller, K.S.; Brzostek, E.R.; Darby, B.A.; Kramer, M.A.; Phillips, R.P. Rhizosphere processes are quantitatively important components of terrestrial carbon and nutrient cycles. Glob. Chang. Biol. 2015, 21, 2082–2094. [Google Scholar] [CrossRef]
- Brunner, I.; Bakker, M.R.; Björk, R.G.; Hirano, Y.; Lukac, M.; Aranda, X.; Børja, I.; Eldhuset, T.D.; Helmisaari, H.S.; Jourdan, C.; et al. Fine-root turnover rates of European forests revisited: An analysis of data from sequential coring and ingrowth cores. Plant Soil 2013, 362, 357–372. [Google Scholar] [CrossRef]
- Brunner, I.; Godbold, D.L. Tree roots in a changing world. J. For. Res. 2007, 12, 78–82. [Google Scholar] [CrossRef]
- Gill, R.A.; Jackson, R. Global patterns of root turnover for terrestrial ecosystems. New Phytol. 2000, 147, 13–31. [Google Scholar] [CrossRef]
- Pietsch, S.A.; Hasenauer, H.; Thornton, P.E. BGC-model parameters for tree species growing in central European forests. For. Ecol. Manag. 2005, 211, 264–295. [Google Scholar] [CrossRef]
- Finer, L.; Ohashi, M.; Noguchi, K.; Hirano, Y. Fine root production and turnover in forest ecosystems in relation to stand and environmental characteristics. For. Ecol. Manag. 2011, 262, 2008–2023. [Google Scholar] [CrossRef]
- Gaul, D.; Hertel, D.; Leuschner, C. Estimating fine root longevity in a temperate Norway spruce forest using three independent methods. Funct. Plant Biol. 2009, 36, 11–19. [Google Scholar] [CrossRef] [PubMed]
- Jourdan, C.; Silva, E.V.; Gonçalves, J.L.M.; Ranger, J.; Moreira, R.M.; Laclau, J.P. Fine root production and turnover in Brazilian Eucalyptus plantations under contrasting nitrogen fertilization regimes. For. Ecol. Manag. 2008, 256, 396–404. [Google Scholar] [CrossRef]
- Yuan, Z.Y.; Chen, H.Y.H. Fine Root Biomass, Production, Turnover Rates, and Nutrient Contents in Boreal Forest Ecosystems in Relation to Species, Climate, Fertility, and Stand Age: Literature Review and Meta-Analyses. Crit. Rev. Plant Sci. 2010, 29, 204–221. [Google Scholar] [CrossRef]
- Gaudinski, J.B.; Torn, M.S.; Riley, W.J.; Dawson, T.E.; Joslin, J.D.; Majdi, H. Measuring and modeling the spectrum of fine-root turnover times in three forests using isotopes, minirhizotrons, and the Radix model. Glob. Biogeochem. Cycles 2010, 24, GB3029. [Google Scholar] [CrossRef]
- Matamala, R.; Gonzalez-Meler, M.A.; Jastrow, J.D.; Norby, R.J.; Schlesinger, W.H. Impacts of fine root turnover on forest NPP and soil C sequestration potential. Science 2003, 302, 1385–1387. [Google Scholar] [CrossRef]
- Persson, H. Fine-root production, mortality and decomposition in forest ecosystems. Vegetatio 1980, 41, 101–109. [Google Scholar] [CrossRef]
- Vogt, K.A.; Persson, H. Measuring growth and development of roots. In Techniques and Approaches in Forest Tree Ecophysiology; Lassoie, J.P., Hinkley, T.M., Eds.; CRC Press: Boca Raton, FL, USA, 1991; pp. 477–501. [Google Scholar]
- Mudrák, O.; Frouz, J.; Velichová, V. Understory vegetation in reclaimed and unreclaimed post-mining forest stands. Ecol. Eng. 2010, 36, 783–790. [Google Scholar] [CrossRef]
- Abakumov, E.V.; Frouz, J. Evolution of the soil humus status on the calcareous Neogene clay dumps of the Sokolov quarry complex in the Czech Republic. Eurasian Soil Sci. 2009, 42, 718–724. [Google Scholar] [CrossRef]
- Rojík, P. New stratigraphic subdivision of the Tertiary in Sokolov Basin in Northwestern Bohemia. J. Czech Geol. Soc. 2004, 49, 173–186. [Google Scholar]
- McCormack, M.L.; Crisfield, E.; Raczka, B.M.; Schnekenburger, F.; Eissensat, D.M.; Smithwick, E.A.H. Sensitivity of four ecological models to adjustments in fine root turnover in temperate forests: Does fine root turnover matter? Ecol. Model. 2015, 297, 107–117. [Google Scholar] [CrossRef]
- Püschel, D.; Rydlová, J.; Vosátka, M. Does the sequence of plant dominants affect mycorrhiza development in simulated succession on spoil banks? Plant Soil 2008, 302, 273–282. [Google Scholar] [CrossRef]
- McClaugherty, C.A.; Aber, J.D.; Melillo, J.M. The role of fine roots in the organic-matter and nitrogen budgets of two forested ecosystems. Ecology 1982, 63, 1481–1490. [Google Scholar] [CrossRef]
- Becklin, K.M.; Hertweck, K.L.; Jumpponen, A. Host identity impacts rhizosphere fungal communities associated with three alpine plant species. Microb. Ecol. 2012, 63, 682–693. [Google Scholar] [CrossRef]
- Becklin, K.M.; Pallo, M.L.; Galen, C. Willows indirectly reduce arbuscular mycorrhizal fungal colonization in understorey communities. J. Ecol. 2012, 100, 343–351. [Google Scholar] [CrossRef]
- Kume, A.; Satomura, T.; Tsuboi, N.; Chiwa, M.; Hanba, Y.T.; Nakane, K.; Horikoshi, T.; Sakugawa, H. Effects of understory vegetation on the ecophysiological characteristics of an overstory pine, Pinus densiflora. For. Ecol. Manag. 2003, 176, 195–203. [Google Scholar] [CrossRef]
- McHugh, T.A.; Gehring, C.A. Below-ground interactions with arbuscular mycorrhizal shrubs decrease the performance of pinyon pine and the abundance of its ectomycorrhizas. New Phytol. 2006, 171, 171–178. [Google Scholar] [CrossRef]
- Harvey, E.; Larsen, M.J.; Jurgensen, M.F. Comparative Distribution of Ectomycorrhizae in Soils of Three Western Montana Forest Habitat Types. For. Sci. 1979, 25, 350–358. [Google Scholar] [CrossRef]
- Gale, M.R.; Grigal, D.F. Vertical root distributions of northern tree species in relation to successional status. Can. J. For. Res. 1987, 17, 829–834. [Google Scholar] [CrossRef]
- Frouz, J.; Kalčík, J.; Velichová, V. Factors causing spatial heterogeneity in soil properties, plant cover, and soil fauna in a non-reclaimed post-mining site. Ecol. Eng. 2010, 37, 1910–1913. [Google Scholar] [CrossRef]
- Read, D.J. Mycorrhizas in ecosystems. Experientia 1991, 47, 376–391. [Google Scholar] [CrossRef]
- Hartmann, P.; von Wilpert, K. Fine-root distributions of Central European Forest soils and their interaction with site and soil properties. Can. J. For. Res. 2014, 44, 71–81. [Google Scholar] [CrossRef]
- Lucas, M.; Schlüter, S.; Vogel, H.J.; Vetterlein, D. Roots change the pore structure only if they have to–development of biopores and compaction around roots. In Geophysical Research Abstracts; EGU: Vienna, Austria, 2019; Volume 21. [Google Scholar]
- Chen, Y.-L.; Zhang, Z.-S.; Huang, L.; Zhao, Y.; Hu, Y.-G.; Zhang, P.; Zhang, D.-H.; Zhang, H. Co-variation of fine-root distribution with vegetation and soil properties along a revegetation chronosequence in a desert area in northwestern China. Catena 2017, 151, 16–25. [Google Scholar] [CrossRef]
- Sun, T.; Dong, L.; Mao, Z.; Li, Y. Fine root dynamics of trees and understorey vegetation in a chronosequence of stands. For. Ecol. Manag. 2015, 346, 1–9. [Google Scholar] [CrossRef]
- Ostonen, I.; Püttsepp, Ü.; Biel, C.; Alberton, O.; Bakker, M.; Löhmus, K.; Majdi, H.; Metcalfe, J.D.; Olsthoorn, A.F.M.; Pronk, A.A.; et al. Specific root length as an indicator of environmental change. Plant Biosyst. 2007, 141, 426–442. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Plot Age (Years) | Vegetation | Dominated Species and Their Cover | Soil Development | pH | C% |
|---|---|---|---|---|---|
| 6 | rarely herbs and grasses | Tussilago farfara (20%), Calamagrostis epigeios (5%) seedlings of Salix caprea 10%. | bare overburden | 8.5 | 5 |
| 16 | dense herbs, scattered shrubs | Salix caprea (20%) Calamagrostis epigeios (15%) | herb litter | 7.5 | 8 |
| 22 | mostly shrubs | Salix caprea (50%) | thick fermentation; layer A horizon absent | 6.8 | 10 |
| 45 | mostly trees | Populus tremuloides (50%), Betula pendula (40%). | A horizon 5 to 10 cm | 6.5 | 12 |
| Root | Root Mass g m−2 | |||||
|---|---|---|---|---|---|---|
| Type | 16-Year-Old Plot | 22-Year-Old Plot | 45-Year-Old Plot | |||
| Herb fine | 28.1 | ±3.5 b | 0.3 | ±3.6 a | 6.2 | ±3.6 a |
| Herb coarse | 5.7 | ±3.6 | 5.5 | ±3.7 | 7.1 | ±3.7 |
| Woody fine | 5.4 | ±22.6 a | 160.3 | ±23.2 b | 109.3 | ±23.2 b |
| Woody coarse | 7.5 | ±16.7 a | 38.6 | 1 ± 7.1 a | 97.4 | ±17.1 b |
| Root mass production (maximum minimum method) g m−2 y−1 | ||||||
| Herb fine | 22 | n/a | 5 | |||
| Woody fine | 14 | 97 | 47 | |||
| Root turnover rate y−1 | ||||||
| Herb fine | 2.59 | n/a | 0.80 | |||
| Woody fine | 0.78 | 0.60 | 0.43 | |||
| Root length m m−2 | ||||||
| Herb fine | 1473 | ±202 c | 31 | ±207 a | 785 | ±207 b |
| Herb coarse | 18 | ±29 | 27 | ±29 | 61 | ±29 |
| Woody fine | 11 | ±35.6 a | 2398 | ±364 c | 1274 | ±364 b |
| Woody coarse | 28 | ±19 a | 53 | ±19 ab | 102 | ±19 b |
| Specific root length m g−1 | ||||||
| Herb fine | 52.4 | ±2.2 | 103.3 | ±22.3 | 126.6 | ±105.8 |
| Woody fine | 2.0 | ±1.4 a | 15.0 | ±4.2 b | 11.7 | ±3.8 b |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kaneda, S.; Zedníková, P.; Frouz, J. Herbaceous and Woody Root Biomass, Seasonal Changes in Root Turnover, and Arbuscular Mycorrhizal and Ectomycorrhizal Colonization during Primary Succession in Post-Mining Sites. Diversity 2022, 14, 644. https://doi.org/10.3390/d14080644
Kaneda S, Zedníková P, Frouz J. Herbaceous and Woody Root Biomass, Seasonal Changes in Root Turnover, and Arbuscular Mycorrhizal and Ectomycorrhizal Colonization during Primary Succession in Post-Mining Sites. Diversity. 2022; 14(8):644. https://doi.org/10.3390/d14080644
Chicago/Turabian StyleKaneda, Satoshi, Petra Zedníková, and Jan Frouz. 2022. "Herbaceous and Woody Root Biomass, Seasonal Changes in Root Turnover, and Arbuscular Mycorrhizal and Ectomycorrhizal Colonization during Primary Succession in Post-Mining Sites" Diversity 14, no. 8: 644. https://doi.org/10.3390/d14080644