
Impact of Native Quercus robur and Non-Native Quercus rubra on Soil Properties during Post-Fire Ecosystem Regeneration


Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Site
2.2. Soil Sampling and Analysis
2.3. Data Evaluation
3. Results
3.1. Basic Soil Parameters
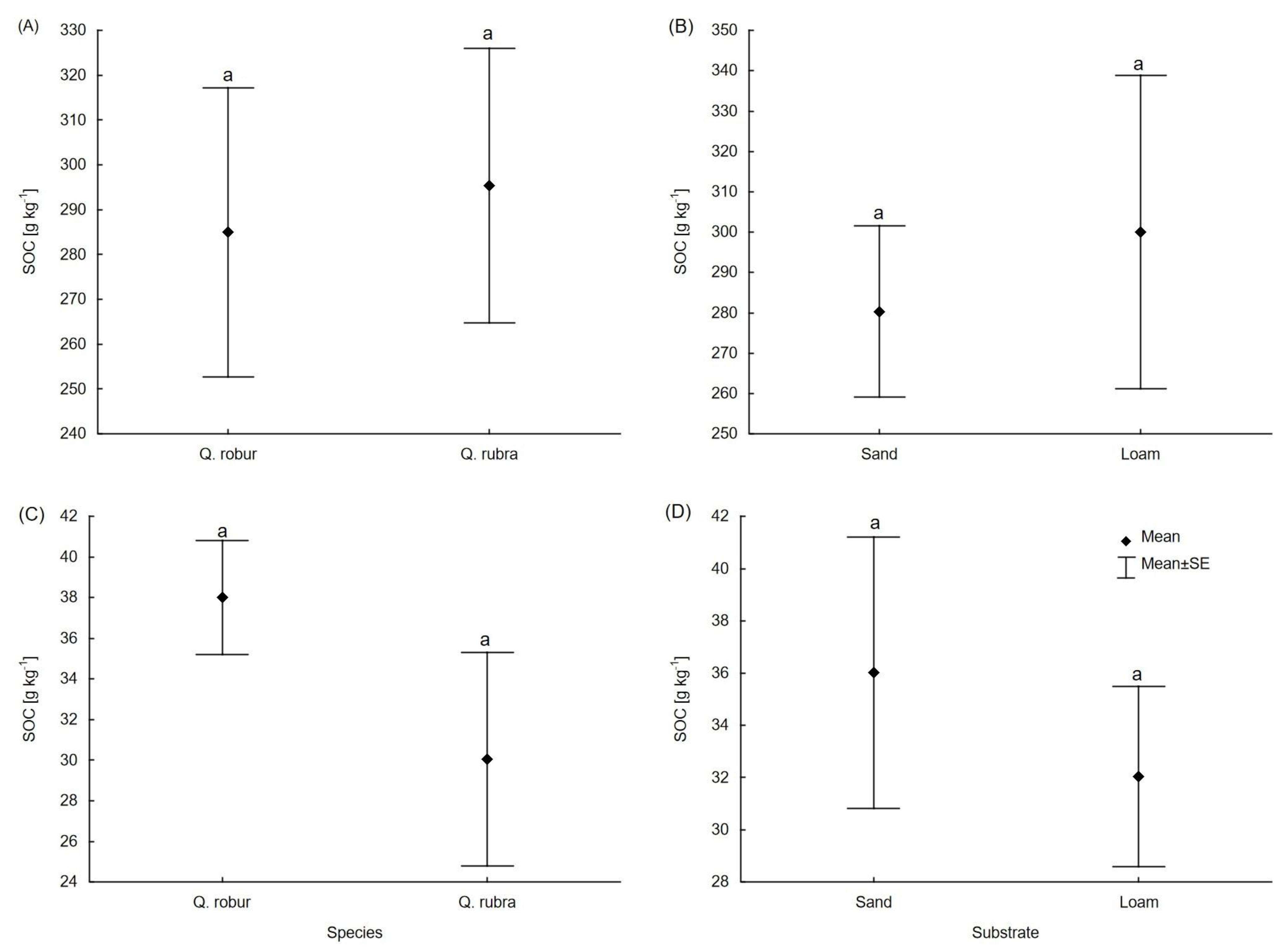
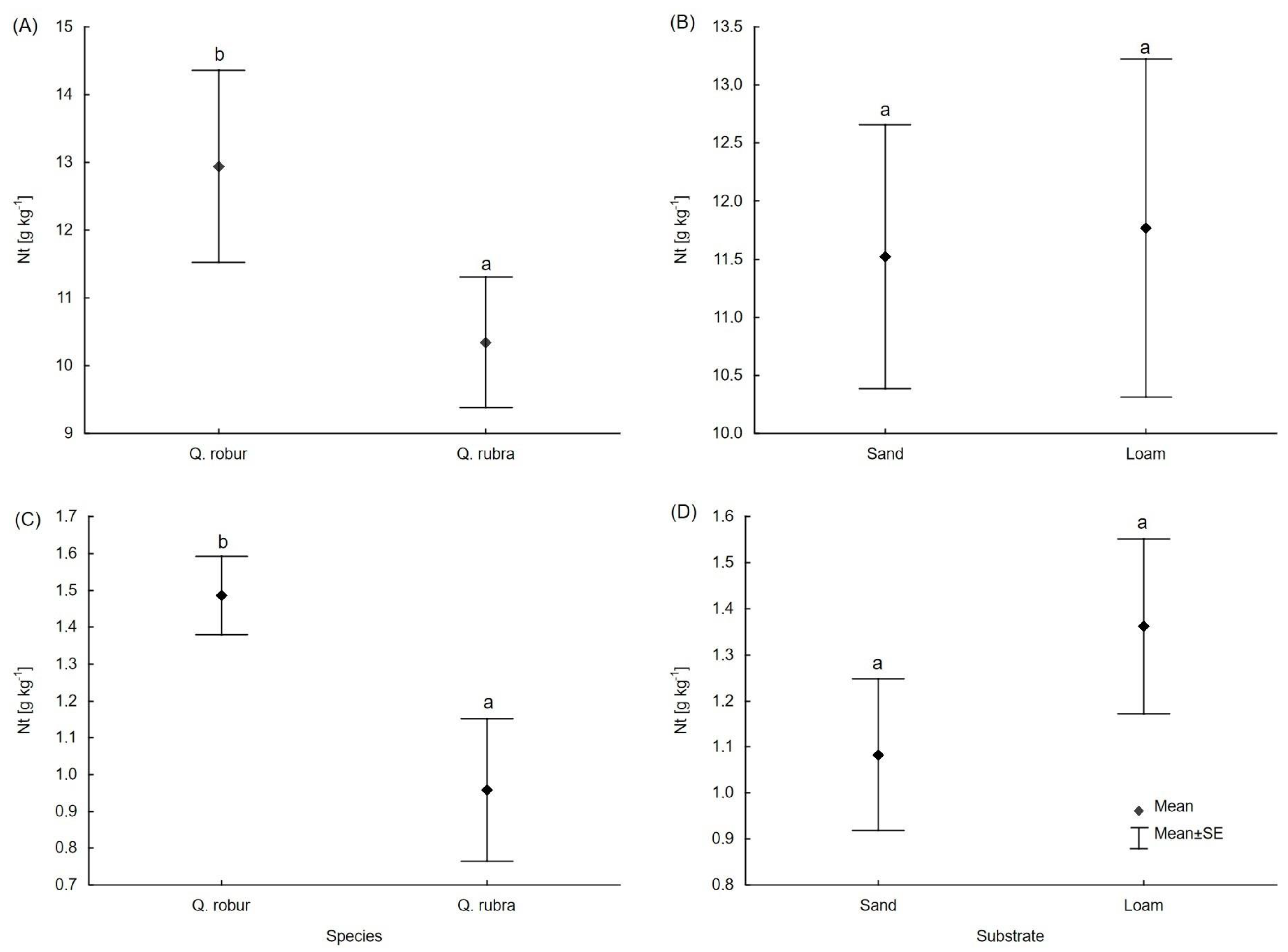
3.2. Carbon and Macronutrient Content
3.3. Microbial Biomass and Respiration
3.4. Relationship between Soil Parameters
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Flannigan, M.D.; Stocks, B.J.; Wotton, B.M. Climate change and forest fires. Sci. Total Environ. 2000, 262, 221–229. [Google Scholar] [CrossRef] [PubMed]
- Pant, S.; Cha, E.J. Potential changes in hurricane risk profile across the United States coastal regions under climate change scenarios. Struct. Saf. 2019, 80, 56–65. [Google Scholar] [CrossRef]
- Certini, G. Effects of fire on properties of forest soils: A review. Oecologia 2005, 143, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Agbeshie, A.A.; Abugre, S.; Atta-Darkwa, T.; Awuah, R. A review of the effects of forest fire on soil properties. J. For. Res. 2022, 33, 1419–1441. [Google Scholar] [CrossRef]
- Rossetti, I.; Cogoni, D.; Calderisi, G.; Fenu, G. Short-Term Effects and Vegetation Response after a Megafire in a Mediterranean Area. Land 2022, 11, 2328. [Google Scholar] [CrossRef]
- Francos, M.; Úbeda, X.; Pereira, P.; Alcañiz, M. Long-term impact of wild fire on soils exposed to different fire severities. A case study in Cadiretes Massif (NE Iberian Peninsula). Sci. Total Environ. 2018, 615, 664–671. [Google Scholar] [CrossRef]
- Kong, J.J.; Yang, J.; Bai, E. Long-term effects of wild fire on available soil nutrient composition and stoichiometry in a Chinese boreal forest. Sci. Total Environ. 2018, 642, 1353–1361. [Google Scholar] [CrossRef]
- Lavoie, M.; Mack, M.C. Spatial heterogeneity of understory vegetation and soil in an Alaskan upland boreal forest fire chronosequence. Biogeochemistry 2012, 107, 227–239. [Google Scholar] [CrossRef]
- De Long, J.R.; Dorrepaal, E.; Kardol, P.; Nilsson, M.-C.; Teuber, L.M.; Wardle, D.A. Understory plant functional groups and litter species identity are stronger drivers of litter decomposition than warming along a boreal forest post-fire successional gradient. Soil Biol. Biochem. 2016, 98, 159–170. [Google Scholar] [CrossRef]
- Baptiste, A.J.; Macario, P.A.; Islebe, G.A.; Vargas-Larreta, B.; Pool, L.; Valdez-Hernández, M.; López-Martínez, J.O. Secondary Succession under invasive species (Pteridium aquilinum) conditions in a seasonal dry tropical forest in southeastern Mexico. PeerJ 2019, 7, e6974. [Google Scholar] [CrossRef] [Green Version]
- Yezzi, A.L.; Nebbia, A.J.; Zalba, S.M. Interaction between fire and fragmentation in the successional stages of coastal dune grasslands of the southern Pampas, Argentina. Sci. Rep. 2019, 9, 15109. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stinca, A.; Ravo, M.; Marzaioli, R.; Marchese, G.; Cordella, A.; Rutigliano, F.A.; Esposito, A. Changes in Multi-Level Biodiversity and Soil Features in a Burned Beech Forest in the Southern Italian Coastal Mountain. Forests 2020, 11, 983. [Google Scholar] [CrossRef]
- Chu, T.; Guo, X.; Takeda, K. Remote sensing approach to detect post-fire vegetation regrowth in Siberian boreal larch forest. Ecol. Indic. 2016, 62, 32–46. [Google Scholar] [CrossRef]
- Coradini, K.; Krejčová, J.; Frouz, J. Potential of vegetation and woodland cover recovery during primary and secondary succession, a global quantitative review. Land Degrad. Dev. 2022, 33, 512–526. [Google Scholar] [CrossRef]
- Wang, X.; He, H.S.; Li, X.; Chang, Y.; Hu, Y.; Xu, C.; Bu, R.; Xie, F. Simulating the effects of reforestation on a large catastrophic fire burned landscape in Northeastern China. For. Ecol. Manag. 2006, 225, 82–93. [Google Scholar] [CrossRef]
- Groninger, J.; Skousen, J.; Angel, P.; Barton, C.; Burger, J.; Zipper, C. Chapter 8: Mine reclamation practices to enhance forest development through natural succession. In The Forestry Reclamation Approach: Guide to Successful Reforestation of Mined Lands; Adams, M.B., Ed.; Gen. Tech. Rep. NRS-169; U.S. Department of Agriculture, Forest Service, Northern Research Station: Newtown Square, PA, USA, 2017; pp. 8-1–8-7. [Google Scholar]
- Yazaki, T.; Hirano, T.; Sano, T. Biomass Accumulation and Net Primary Production during the Early Stage of Secondary Succession after a Severe Forest Disturbance in Northern Japan. Forests 2016, 7, 287. [Google Scholar] [CrossRef]
- Stevens-Rumann, C.S.; Morgan, P. Tree regeneration following wildfires in the western US: A review. Fire Ecol. 2019, 15, 15. [Google Scholar] [CrossRef] [Green Version]
- Ceitel, J.; Barzdajn, W.; Zientarski, J. The influence of the soil preparation after the forest fire on the survival and growth of the selected tree species. Sylwan 2003, 147, 3–13. [Google Scholar]
- Woś, B.; Józefowska, A.; Likus-Cieślik, J.; Chodak, M.; Pietrzykowski, M. Effect of tree species and soil texture on the carbon stock, macronutrient content, and physicochemical properties of regenerated postfire forest soils. Land Degrad. Dev. 2021, 32, 5227–5240. [Google Scholar] [CrossRef]
- Robakowski, P.; Balicki, P.; Łukaszewski, M.; Szczerba, M. Early growth of Scots pine, European larch and common oak at the young age on post-fire area located in the Sława Śląska Forest District (SW Poland). Ecol. Quest. 2014, 20, 53–61. [Google Scholar] [CrossRef]
- Nicolescu, V.-N.; Vor, T.; Mason, W.L.; Bastien, J.-C.; Brus, R.; Henin, J.-M.; Kupka, I.; Lavnyy, V.; La Porta, N.; Mohren, F.; et al. Ecology and management of northern red oak (Quercus rubra L. syn. Q. borealis F. Michx.) in Europe: A review. Forestry 2020, 93, 481–494. [Google Scholar]
- Gazda, A.; Szwagrzyk, J. Introduced species in Polish National Parks: Distribution, abundance and management approaches. In Introduced Tree Species in European Forests: Opportunities and Challenges; Krumm, F., Vítková, L., Eds.; European Forest Institute: Joensuu, Finland, 2016; pp. 168–175. [Google Scholar]
- Wohlgemuth, T.; Gossner, M.M.; Campagnaro, T.; Marchante, H.; van Loo, M.; Vacchiano, G.; Castro-Díez, P.; Dobrowolska, D.; Gazda, A.; Keren, S.; et al. Impact of non-native tree species in Europe on soil properties and biodiversity: A review. NeoBiota 2022, 78, 45–69. [Google Scholar] [CrossRef]
- Pötzelsberger, E.; Spiecker, H.; Neophytou, C.; Mohren, F.; Gazda, A.; Hasenauer, H. Growing Non-native Trees in European Forests Brings Benefits and Opportunities but Also Has Its Risks and Limits. Curr. For. Rep. 2020, 6, 339–353. [Google Scholar] [CrossRef]
- Stanek, M.; Piechnik, Ł.; Stefanowicz, A.M. Invasive red oak (Quercus rubra L.) modifies soil physicochemical properties and forest understory vegetation. For. Ecol. Manag. 2020, 472, 118253. [Google Scholar] [CrossRef]
- Riepšas, E.; Straigyte, L. Invasiveness and Ecological Effects of Red Oak (Quercus rubra L.) in Lithuanian Forests. Balt. For. 2008, 14, 122–130. [Google Scholar]
- Woziwoda, B.; Kopeć, D.; Witkowski, J. The negative impact of intentionally introduced Quercus rubra L. on a forest community. Acta Soc. Bot. Pol. 2014, 83, 39–49. [Google Scholar] [CrossRef]
- Gómez-González, S.; Torres-Díaz, C.; Valencia, G.; Torres-Morales, P.; Cavieres, L.A.; Pausas, J.G. Anthropogenic fires increase alien and native annual species in the Chilean coastal matorral. Divers. Distrib. 2011, 17, 58–67. [Google Scholar] [CrossRef]
- Bonanomi, G.; Incerti, G.; El-Gawad, A.M.A.; Sarker, T.C.; Stinca, A.; Motti, R.; Cesarano, G.; Teobaldelli, M.; Saulino, L.; Cona, F.; et al. Windstorm disturbance triggers multiple species invasion in an urban Mediterranean forest. iForest 2018, 11, 64–71. [Google Scholar] [CrossRef] [Green Version]
- Larson, D.L.; Anderson, P.J.; Newton, W. Alien plant invasion in mixed-grass prairie: Effects of vegetation type and anthropogenic disturbance. Ecol. Appl. 2001, 11, 128–141. [Google Scholar] [CrossRef]
- Kraus, D.; Silva, J.S.; Held, A.; Castellnou, M.; Krumm, F. Natural disturbances and invasive introduced plants—A specific view on fire. In Introduced Tree Species in European Forests: Opportunities and Challenges; Krumm, F., Vítková, L., Eds.; European Forest Institute: Joensuu, Finland, 2016; pp. 294–304. [Google Scholar]
- Pietrzykowski, M.; Woś, B. The Impact of Climate Change on Forest Tree Species Dieback and Changes in Their Distribution. In Climate Change and the Microbiome; Choudhary, D.K., Mishra, A., Varma, A., Eds.; Soil Biology 63; Springer: Cham, Switzerland, 2021; pp. 447–460. [Google Scholar]
- Suzuki, K.F.; Kobayashi, Y.; Seidl, R.; Senf, C.; Tatsumi, S.; Koide, D.; Azuma, W.A.; Higa, M.; Koyanagi, T.F.; Qian, S.; et al. The potential role of an alien tree species in supporting forest restoration: Lessons from Shiretoko National Park, Japan. For. Ecol. Manag. 2021, 493, 119253. [Google Scholar] [CrossRef]
- Woś, B.; Pająk, M.; Pietrzykowski, M. Soil Organic Carbon Pools and Associated Soil Chemical Properties under Two Pine Species (Pinus sylvestris L. and Pinus nigra Arn.) Introduced on Reclaimed Sandy Soils. Forests 2022, 13, 328. [Google Scholar] [CrossRef]
- Rai, P.K. Environmental Degradation by Invasive Alien Plants in the Anthropocene: Challenges and Prospects for Sustainable Restoration. Anthr. Sci. 2022, 1, 5–28. [Google Scholar] [CrossRef]
- Augusto, L.; Ranger, J.; Binkley, D.; Rothe, A. Impact of several common tree species of European temperate forests on soil fertility. Ann. For. Sci. 2002, 59, 233–253. [Google Scholar] [CrossRef] [Green Version]
- Woś, B.; Chodak, M.; Józefowska, A.; Pietrzykowski, M. Influence of tree species on carbon, nitrogen, and phosphorus stocks and stoichiometry under different soil regeneration scenarios on reclaimed and afforested mine and post-fire forest sites. Geoderma 2022, 415, 115782. [Google Scholar] [CrossRef]
- Hobbie, S.E. Effects of plant species on nutrient cycling. Trends Ecol. Evol. 1992, 7, 336–339. [Google Scholar] [CrossRef]
- Binkley, D.; Giardina, C. Why do tree species affect soils? The Warp and Woof of tree-soil interactions. Biogeochemistry 1998, 42, 89–106. [Google Scholar] [CrossRef]
- Pietrzykowski, M. Tree species selection and reaction to mine soil reconstructed at reforested post-mine sites: Central and eastern European experiences. Ecol. Eng. Suppl. 2019, 3, 100012. [Google Scholar] [CrossRef]
- Maiti, S.K.; Bandyopadhyay, S.; Mukhopadhyay, S. Chapter 14—Importance of selection of plant species for successful ecological restoration program in coal mine degraded land. In Phytorestoration of Abandoned Mining and Oil Drilling Sites; Bauddh, K., Korstad, J., Sharma, P., Eds.; Elsevier Inc.: Amsterdam, The Netherlands, 2021; pp. 325–357. [Google Scholar]
- Chodak, M.; Sroka, K.; Woś, B.; Pietrzykowski, M. Chemical and microbial properties of post-mining and post-fire soils afforested with different tree species. Appl. Soil Ecol. 2022, 171, 104321. [Google Scholar] [CrossRef]
- Szabla, K. Preconditions for outbreak and evolving of fires, some organizational activities, current silvicultural and protection problems on the burnt area in the Rudy Raciborskie Forest District. Sylwan 1994, 138, 75–83. [Google Scholar]
- Orczewska, A.; Prukop, M.; Strzelczyk, A. Recovery of the herbaceous layer in the young silver birch and black alder stands that developed spontaneously after a forest fire. Ecol. Res. 2016, 31, 125–133. [Google Scholar] [CrossRef] [Green Version]
- IUSS Working Group WRB. World reference base for soil resources 2014, update 2015. International soil classification system for naming soils and creating legends for soil maps. In World Soil Resources Reports; FAO: Rome, Italy, 2015; p. 105. [Google Scholar]
- Ferré, C.; Comolli, R. Effects of Quercus rubra L. on soil properties and humus forms in 50-year-old and 80-year-old forest stands of Lombardy plain. Ann. For. Sci. 2020, 77, 3. [Google Scholar] [CrossRef]
- Bonifacio, E.; Petrillo, M.; Petrella, F.; Tambone, F.; Celi, L. Alien red oak affects soil organic matter cycling and nutrient availability in low-fertility well-developed soils. Plant Soil 2015, 395, 215–229. [Google Scholar] [CrossRef]
- Jonczak, J.; Parzych, A.; Sobisz, Z. Decomposition of four tree species leaf litters in headwater riparian forest. Balt. For. 2015, 21, 133–143. [Google Scholar]
- Nicolini, F.; Topp, W. Soil properties in plantations of sessile oak (Quercus petraea) and red oak (Quercus rubra) in reclaimed lignite open-cast mines of the Rhineland. Geoderma 2005, 129, 65–72. [Google Scholar] [CrossRef]
- Hallet, R.A.; Hornbeck, J.W. Foliar and soil nutrient relationships in red oak and white pine forests. Can. J. For. Res. 1997, 27, 1233–1244. [Google Scholar] [CrossRef]
- Stanek, M.; Zubek, S.; Stefanowicz, A.M. Differences in phenolics produced by invasive Quercus rubra and native plant communities induced changes in soil microbial properties and enzymatic activity. For. Ecol. Manag. 2021, 482, 118901. [Google Scholar] [CrossRef]
- Demoling, F.; Figueroa, D.; Baath, E. Comparison of factors limiting bacterial growth in different soils. Soil Biol. Biochem. 2007, 39, 2485–2495. [Google Scholar] [CrossRef]
- Datta, A.; Shrestha, S.; Ferdous, Z.; Win, C.C. Strategies for enhancing phosphorus efficiency in crop production systems. In Nutrient Use Efficiency: From Basics to Advances; Rakshit, A., Singh, H.B., Sen, A., Eds.; Springer: New Delhi, India, 2015; pp. 59–71. [Google Scholar]
- Frouz, J.; Pižl, V.; Cienciala, E.; Kalčík, J. Carbon storage in post-mining forest soil, the role of tree biomass and soil bioturbation. Biogeochemistry 2009, 94, 111–121. [Google Scholar] [CrossRef]
- Pausas, J.G.; Bladé, C.; Valdecantos, A.; Seva, J.P.; Fuentes, D.; Alloza, J.A.; Vilagrosa, A.; Bautista, S.; Cortina, J.; Vallejo, R. Pines and oaks in the restoration of Mediterranean landscapes of Spain: New perspectives for an old practice—A review. Plant Ecol. 2004, 171, 209–220. [Google Scholar] [CrossRef]
- Enríquez-de-Salamanca, E. Dynamics of Mediterranean pine forests reforested after fires. J. For. Res. 2023, 34, 345–354. [Google Scholar] [CrossRef]
- Pietrzykowski, M.; Krzaklewski, W.; Likus, J.; Woś, B. Assessment of english oak (Quercus robur L.) growth in varied soil-substrate conditions of reclaimed Piaseczno sulfur mine dump. Folia For. Pol. 2015, 57, 28–32. [Google Scholar] [CrossRef] [Green Version]
- Lévy, G.; Becker, M.; Duhamel, D. A comparison of the ecology of pedunculate and sessile oaks: Radial growth in the centre and northwest of France. For. Ecol. Manag. 1992, 55, 51–63. [Google Scholar] [CrossRef]
- Ellenberg, H. Vegetation Ecology of Central Europe, 4th ed.; Cambridge University Press: Cambridge, UK, 2009. [Google Scholar]



{kind=link}
{kind=link}
| Effect | Soil Parameters/Horizons [cm] | |||||||
|---|---|---|---|---|---|---|---|---|
| Sand (2.00–0.05 mm) | Silt (0.05–0.002 mm) | Clay (<0.002 mm) | BD | pHH2O | Mass (d. w.) | |||
| 0–5 cm | Oi + Oe | 0–5 cm | Oi + Oe | |||||
| [%] | [g cm−3] | [kg m−2] | ||||||
| Species | Q. robur | 65 ± 9 a,1 | 32 ± 8 b | 3 ± 1 a | 1.15 ± 0.04 a | 5.0 ± 0.1 a | 4.1 ± 0.0 b | 4.3 ± 0.3 a |
| Q. rubra | 71 ± 8 a | 26 ± 7 a | 3 ± 1 a | 1.24 ± 0.09 a | 4.6 ± 0.2 a | 3.9 ± 0.1 a | 7.9 ± 1.3 b | |
| Parent material | Sand | 89 ± 1 b | 10 ± 1 a | 1 ± 0 a | 1.12 ± 0.09 a | 4.6 ± 0.1 a | 4.0 ± 0.1 a | 6.1 ± 1.2 a |
| Loam | 47 ± 3 a | 48 ± 3 b | 6 ± 0 b | 1.27 ± 0.03 a | 4.9 ± 0.2 a | 4.0 ± 0.1 a | 6.1 ± 1.1 a | |
| Effect | Nutrient/Horizons [cm] | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| S | Ca | K | Mg | Na | P | ||||||||
| Oi + Oe | 0–5 cm | Oi + Oe | 0–5 cm | Oi + Oe | 0–5 cm | Oi + Oe | 0–5 cm | Oi + Oe | 0–5 cm | Oi + Oe | 0–5 cm | ||
| [%] | [g kg−1] | ||||||||||||
| Species | Q. robur | 1.20 ± 0.12 a,1 | 0.24 ± 0.02 a | 4.62 ± 0.49 | 0.29 ± 0.05 b | 1.32 ± 0.17 a | 1.11 ± 0.26 b | 1.12 ± 0.06 b | 0.48 ± 0.12 b | 0.06 ± 0.00 a | 0.09 ± 0.01 a | 0.73 ± 0.05 b | 0.18 ± 0.02 a |
| Q. rubra | 1.10 ± 0.05 a | 0.17 ± 0.03 a | 3.50 ± 0.83 | 0.20 ± 0.04 a | 1.17 ± 0.14 a | 0.87 ± 0.23 a | 0.78 ± 0.13 a | 0.35 ± 0.10 a | 0.07 ± 0.01 a | 0.08 ± 0.01 a | 0.57 ± 0.05 a | 0.12 ± 0.02 a | |
| Parent material | Sand | 1.13 ± 0.07 a | 0.18 ± 0.03 a | 4.17 ± 0.72 a | 0.13 ± 0.03 a | 0.88 ± 0.07 a | 0.37 ± 0.03 a | 0.89 ± 0.14 a | 0.14 ± 0.02 a | 0.05 ± 0.00 a | 0.05 ± 0.00 a | 0.61 ± 0.06 a | 0.14 ± 0.03 a |
| Loam | 1.17 ± 0.12 a | 0.22 ± 0.02 a | 3.95 ± 0.70 a | 0.36 ± 0.03 b | 1.61 ± 0.09 b | 1.61 ± 0.11 b | 1.02 ± 0.09 a | 0.69 ± 0.06 b | 0.08 ± 0.01 b | 0.12 ± 0.00 b | 0.69 ± 0.05 a | 0.16 ± 0.02 a | |
| Effect | Characteristics/Horizons [cm] | ||||
|---|---|---|---|---|---|
| RESP | Cmic | ||||
| Oi + Oe | 0–5 cm | Oi + Oe | 0–5 cm | ||
| [μM CO2 g−1 24 h−1] | [μg g−1] | ||||
| Species | Q. robur | 40.96 ± 2.40 a, 1 | 1.00 ± 0.08 a | 2202.15 ± 180.01 a | 169.89 ± 14.97 a |
| Q. rubra | 39.11 ± 11.21 a | 0.94 ± 0.19 a | 2187.37 ± 460.45 a | 164.39 ± 29.77 a | |
| Parent material | Sand | 37.07 ± 6.07 a | 1.07 ± 0.19 a | 2143.74 ± 264.27 a | 187.47 ± 29.29 a |
| Loam | 43.01 ± 9.60 a | 0.87 ± 0.06 a | 2245.78 ± 416.96 a | 146.81 ± 11.67 a | |
| RESP | Cmic | SOC | |
|---|---|---|---|
| Sand | 0.09 | 0.19 | −0.01 |
| Silt | −0.05 | −0.15 | 0.03 |
| Clay | −0.33 | −0.36 | −0.29 |
| pH | 0.46 | 0.37 | 0.06 |
| SOC | 0.68 * | 0.47 | 1.00 |
| Nt | 0.49 | 0.23 | 0.84 * |
| S | 0.60 * | 0.38 | 0.78 * |
| Ca | 0.12 | 0.03 | 0.04 |
| K | −0.01 | −0.08 | 0.04 |
| Mg | 0.01 | −0.07 | 0.01 |
| Na | −0.15 | −0.25 | −0.09 |
| P | 0.62 * | 0.57 * | 0.53 * |
| BD | −0.68 * | −0.64 * | −0.48 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Woś, B.; Józefowska, A.; Wanic, T.; Pietrzykowski, M. Impact of Native Quercus robur and Non-Native Quercus rubra on Soil Properties during Post-Fire Ecosystem Regeneration. Diversity 2023, 15, 559. https://doi.org/10.3390/d15040559
Woś B, Józefowska A, Wanic T, Pietrzykowski M. Impact of Native Quercus robur and Non-Native Quercus rubra on Soil Properties during Post-Fire Ecosystem Regeneration. Diversity. 2023; 15(4):559. https://doi.org/10.3390/d15040559
Chicago/Turabian StyleWoś, Bartłomiej, Agnieszka Józefowska, Tomasz Wanic, and Marcin Pietrzykowski. 2023. "Impact of Native Quercus robur and Non-Native Quercus rubra on Soil Properties during Post-Fire Ecosystem Regeneration" Diversity 15, no. 4: 559. https://doi.org/10.3390/d15040559