
Formica picea and F. candida (Hymenoptera: Formicidae): Synonyms or Two Species?

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
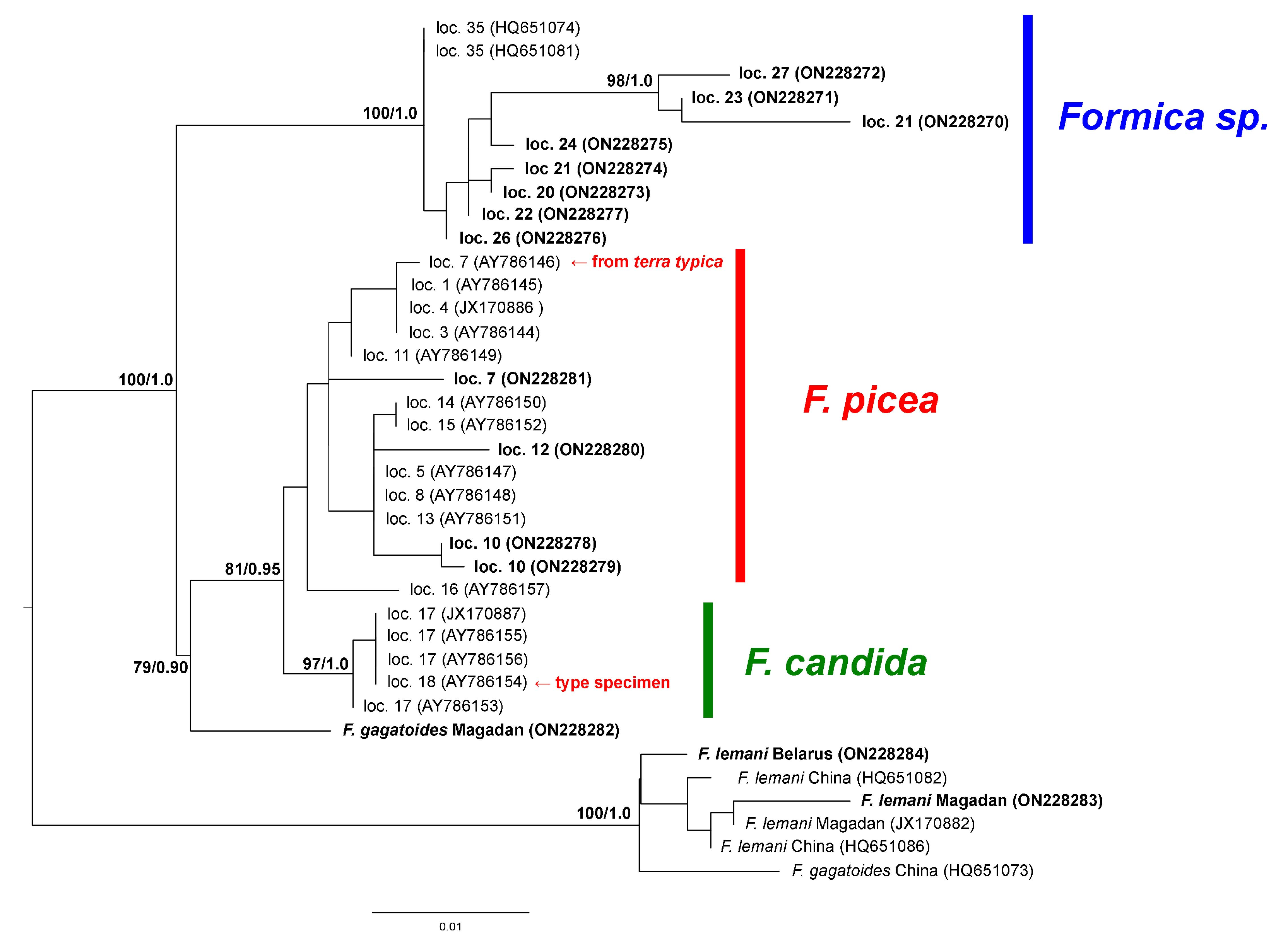
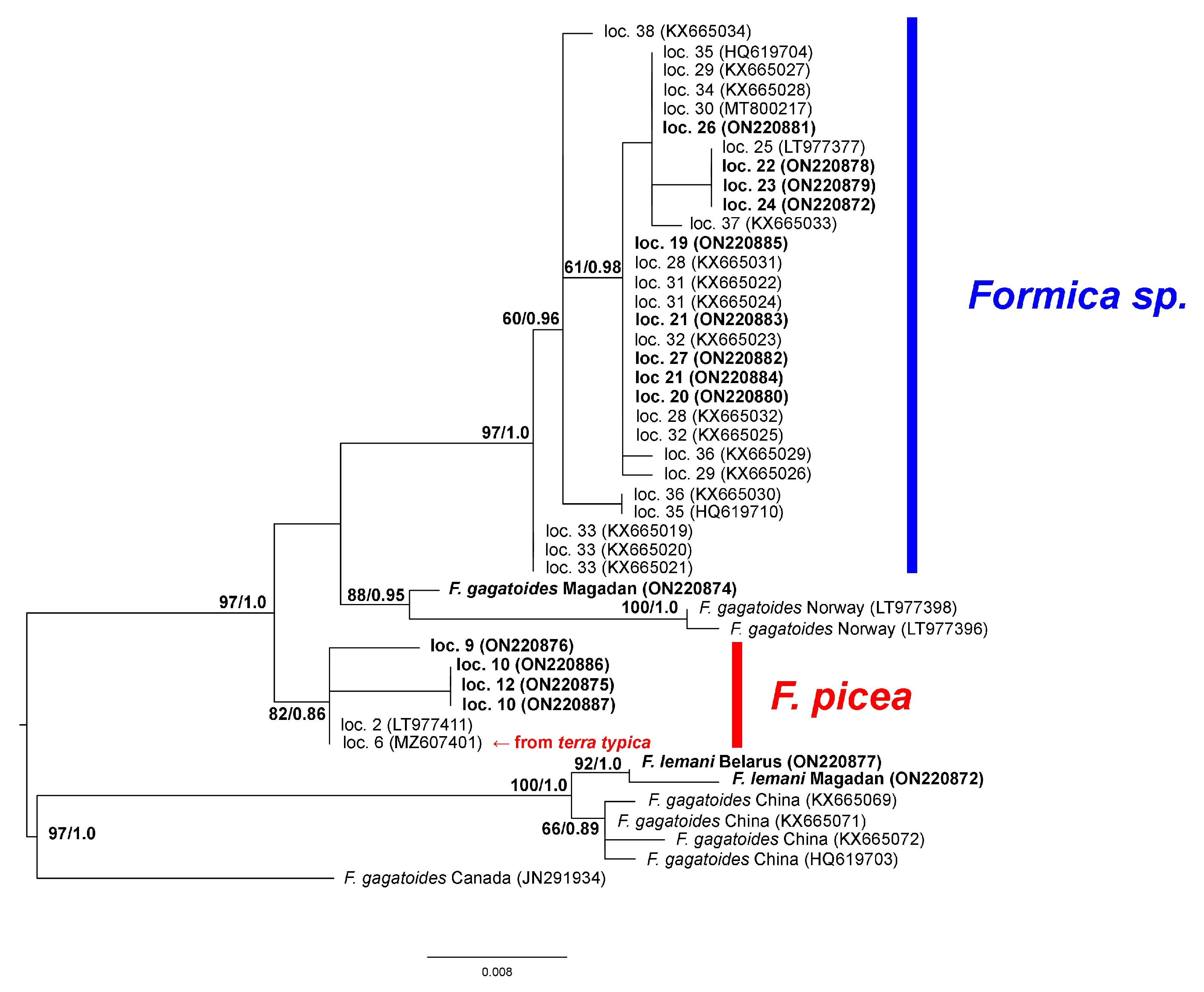
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Borowiec, M.L.; Moreau, C.S.; Rabeling, C. Ants: Phylogeny and Classification. In Encyclopedia of Social Insects; Springer International Publishing: Cham, Switzerland, 2020; pp. 1–18. [Google Scholar]
- Valenzuela-González, J.E.; Pérez-Toledo, G.R.; García-Martínez, M.A. Adelomyrmex dorae sp. nov. Garcia-Martinez (Hymenoptera: Formicidae): A New Species Supported by Parsimony Analysis of Morphological Characters. Trans. Am. Entomol. Soc. 2017, 143, 713–727. [Google Scholar] [CrossRef]
- Goropashnaya, A. Phylogeographic Structure and Genetic Variation in Formica Ants. Ph.D. Thesis, Uppsala University, Uppsala, Sweden, 20 December 2003. [Google Scholar]
- Schär, S.; Talavera, G.; Espadaler, X.; Rana, J.D.; Andersen, A.; Cover, S.P.; Vila, R. Do Holarctic ant species exist? Trans-Beringian dispersal and homoplasy in the Formicidae. J. Biogeogr. 2018, 45, 1917–1928. [Google Scholar] [CrossRef]
- Goropashnaya, A.V.; Fedorov, V.B.; Seifert, B.; Pamilo, P. Phylogenetic relationships of Palaearctic Formica species (Hymenoptera, Formicidae) based on mitochondrial cytochrome b sequences. PLoS ONE 2012, 7, e41697. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roslin, T.; Somervuo, P.; Pentinsaari, M.; Hebert, P.D.N.; Agda, J.; Ahlroth, P.; Anttonen, P.; Aspi, J.; Blagoev, G.; Blanco, S.; et al. A molecular-based identification resource for the arthropods of Finland. Mol. Ecol. Resour. 2022, 22, 803–822. [Google Scholar] [CrossRef]
- Chen, Y.; Zhou, S. Phylogenetic relationships based on DNA barcoding among 16 species of the ant genus Formica (Hymenoptera: Formicidae) from China. J. Insect Sci. 2017, 17, 117. [Google Scholar] [CrossRef] [Green Version]
- Park, S.; Noh, P.; Kang, J.-Y. Endosymbionts and phage WO infections in Korean ant species (Hymenoptera: Formicidae). Proc. Natl. Inst. Ecol. Repub. Korea 2020, 1, 52–57. [Google Scholar] [CrossRef]
- Chen, Z.; Zhou, S.Y.; Ye, D.; Chen, Y.; Lu, C. Molecular phylogeny of the ant subfamily Formicinae (Hymenoptera, Formicidae) from China Based on mitochondrial genes. Sociobiology 2013, 60, 135–144. [Google Scholar] [CrossRef] [Green Version]
- Shekhovtsov, S.V.; Berman, D.I.; Bulakhova, N.A.; Makarova, O.L.; Peltek, S.E. Phylogeography of earthworms from high latitudes of Eurasia. Acta Zool. Acad. Sci. Hung. 2018, 64, 369–382. [Google Scholar] [CrossRef]
- Dlussky, G.M. Ants of the Genus Formica (Hymenoptera: Formicidae g. Formica); Nauka: Moscow, Russia, 1967; p. 236. [Google Scholar]
- Bolton, B. A New General Catalogue of the Ants of the World; Harvard University Press: Cambridge, UK, 1995; p. 502. [Google Scholar]
- Seifert, B. The “Black Bog Ant” Formica picea Nylander, 1846—A species different from Formica candida Smith, 1878 (Hymenoptera: Formicidae). Myrmecolog. Nachr. 2004, 6, 29–38. [Google Scholar]
- Zakharov, A.A.; Dlussky, G.M.; Goryunov, D.N.; Gilyov, A.V.; Zryanin, V.A.; Fedoseeva, E.B.; Gorokhovskaya, E.A.; Radchenko, A.G. Monitoring of the Formica Ants; KMK Scientific Press: Moscow, Russia, 2013; p. 99. [Google Scholar]
- Ward, P.S.; Brady, S.G.; Fisher, B.L.; Schultz, T.R. The evolution of myrmicine ants: Phylogeny and biogeography of a hyperdiverse ant clade (Hymenoptera: Formicidae). Syst. Entomol. 2015, 40, 61–81. [Google Scholar] [CrossRef] [Green Version]
- Antonov, I.A.; Bukin, Y.S. Molecular phylogenetic analysis of the ant genus Formica L. (Hymenoptera: Formicidae) from Palearctic region. Russ. J. Genet. 2016, 52, 810–820. [Google Scholar] [CrossRef]
- Berman, D.I.; Alfimov, A.V.; Zhigulskaya, Z.A.; Leirikh, A.N. Overwintering and Cold-Hardiness of Ants in the Northeast of Asia; Pensoft Publishers: Sofia-Moscow, Russia, 2010; p. 294. [Google Scholar]
- Radchenko, A.G. Ants (Hymenoptera, Formicidae) of Ukraine; Institute of Zoology of Ukraine NAS: Kyiv, Ukraine, 2016; p. 495. [Google Scholar]
- Folmer, O.; Hoeh, W.R.; Black, M.B.; Vrijenhoek, R.C. Conserved primers for PCR amplification of mitochondrial DNA from different invertebrate phyla. Mol. Mar. Biol. Biotechnol. 1994, 3, 294–299. [Google Scholar]
- Shekhovtsov, S.V.; Shipova, A.A.; Poluboyarova, T.V.; Vasiliev, G.V.; Golovanova, E.V.; Geraskina, A.P.; Bulakhova, N.A.; Szederjesi, T.; Peltek, S.E. Species delimitation of the Eisenia nordenskioldi complex (Oligochaeta, Lumbricidae) using transcriptomic data. Front. Genet. 2020, 11, 1508. [Google Scholar] [CrossRef]
- Stamatakis, A. RAxML version 8: A tool for phylogenetic analysis and post-analysis of large phylogenies. Bioinformatics 2014, 30, 1312–1313. [Google Scholar] [CrossRef]
- Nylander, J.A.A. MrModeltest Ver. 2; Evolutionary Biology Centre, Uppsala University: Uppsala, Sweden, 2004. [Google Scholar]
- Ronquist, F.; Teslenko, M.; Van Der Mark, P.; Ayres, D.L.; Darling, A.; Höhna, S.; Larget, B.; Liu, L.; Suchard, M.A.; Huelsenbeck, J.P. MrBayes 3.2: Efficient Bayesian phylogenetic inference and model choice across a large model space. Syst. Biol. 2012, 61, 539–542. [Google Scholar] [CrossRef] [Green Version]
- Seifert, B. Cryptic species in ants (Hymenoptera: Formicidae) revisited: We need a change in the alpha-taxonomic approach. Myrmecol. News 2009, 12, 149–166. [Google Scholar]
- Bernasconi, C.; Cherix, D.; Seifert, B.; Pamilo, P. Molecular taxonomy of the Formica rufa group (red wood ants) (Hymenoptera: Formicidae): A new cryptic species in the Swiss Alps. Myrmecol. News 2011, 14, 37–47. [Google Scholar]
- Seifert, B. Inconvenient hyperdiversity—The traditional concept of “Pheidole pallidula” includes four cryptic species (Hymenoptera: Formicidae). SOIL Org. 2016, 88, 1–17. [Google Scholar]
- Fournier, D.; Tindo, M.; Kenne, M.; Mbenoun Masse, P.S.; Van Bossche, V.; De Coninck, E.; Aron, S. Genetic structure, nestmate recognition and behaviour of two cryptic species of the invasive big-headed ant Pheidole megacephala. PLoS ONE 2012, 7, e31480. [Google Scholar] [CrossRef] [Green Version]
- Seifert, B.; Yazdi, A.B.; Schultz, R. Myrmica martini sp. n.—A cryptic species of the Myrmica scabrinodis species complex (Hymenoptera: Formicidae) revealed by geometric morphometrics and nest-centroid clustering. Myrmecol. News 2014, 19, 171–183. [Google Scholar]
- Ebsen, J.R.; Boomsma, J.J.; Nash, D.R. Phylogeography and cryptic speciation in the Myrmica scabrinodis Nylander, 1846 species complex (Hymenoptera: Formicidae), and their conservation implications. Insect Conserv. Divers. 2019, 12, 467–480. [Google Scholar] [CrossRef] [Green Version]
- Seifert, B. A taxonomic revision of the Palaearctic members of the Formica rufa group (Hymenoptera: Formicidae)–The famous mound-building red wood ants. Myrmecol. News 2021, 31, 133–179. [Google Scholar] [CrossRef]
- Pamilo, P.; Kulmuni, J. Genetic identification of Formica rufa group species and their putative hybrids in northern Europe. Myrmecol. News 2022, 32. [Google Scholar] [CrossRef]
- Sanllorente, O.; Lorite, P.; Ruano, F.; Palomeque, T.; Tinaut, A. Phylogenetic relationships between the slave-making ants Rossomyrmex and their Proformica hosts in relation to other genera of the ant tribe Formicini (Hymenoptera: Formicidae). J. Zool. Syst. Evol. Res. 2018, 56, 48–60. [Google Scholar] [CrossRef]
- Smith, F. Scientific Results of the Second Yarkand Mission; Based upon the Collections and Notes of the Late Ferdinand Stoliczka, Ph.D.; Superintendent of Government Printing (Government of India): Calcutta, India, 1878; p. 22. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
| Location | Species | cox1 | cytb |
|---|---|---|---|
| Formica picea | |||
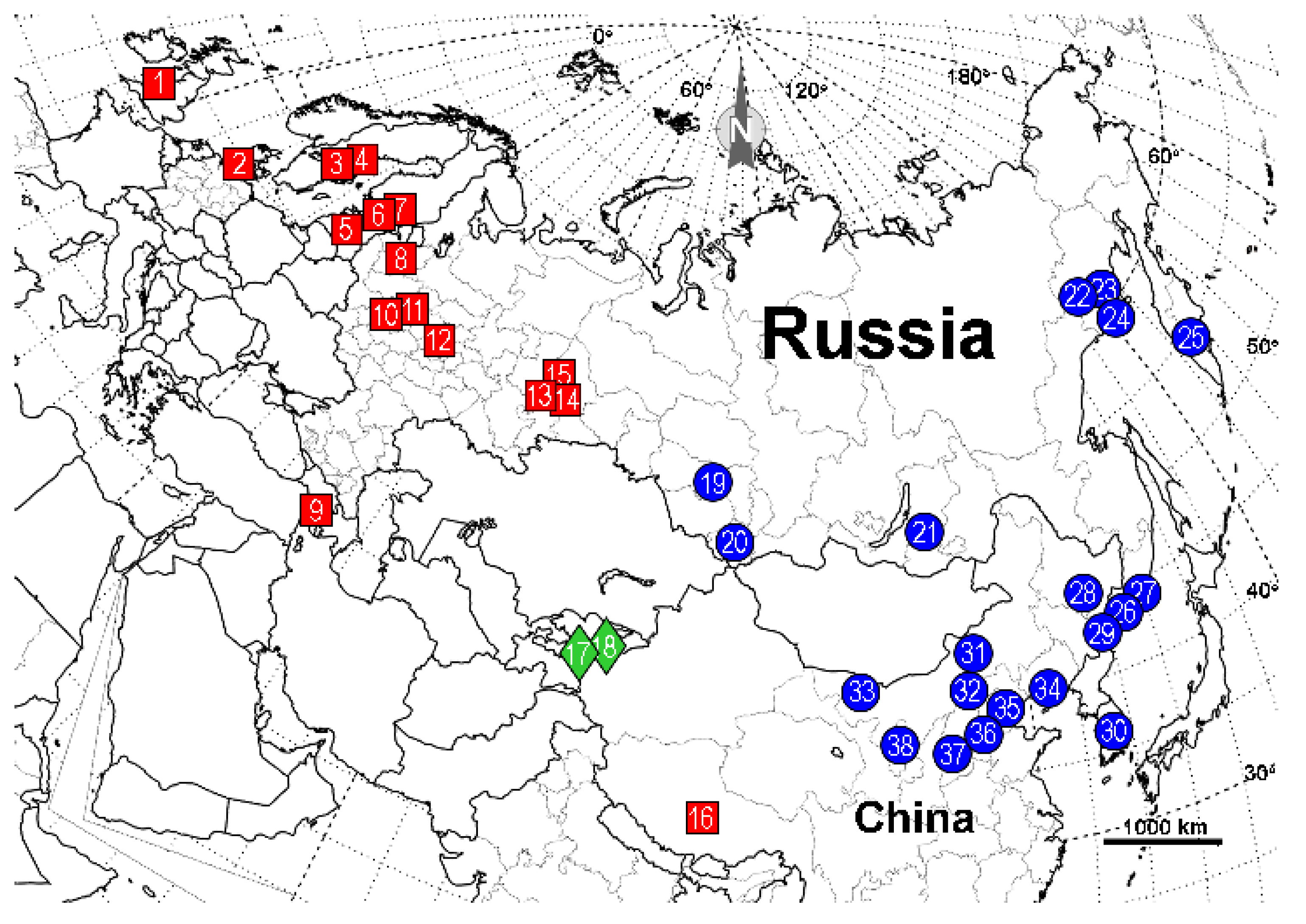
| 1 | United Kingdom: Cors Goch National Nature Reserve, S. Wales | - | AY786145 (as F. candida) [3] |
| 2 | Denmark: N55.83 E12.56 | LT977411 [4] | - |
| 3 | Sweden: Trajmossen, Brattforsheden | - | AY786144 (as F. candida) [3] |
| 4 | Sweden: Varmland | - | JX170886 [5] |
| 5 | Estonia: Torma | - | AY786147 (as F. candida) [3] |
| 6 | Finland: Nylandia, Raasepori, Harpar Stortraesket | MZ607401 * [6] | - |
| 7 | Finland: Helsinki, Vihti | - | AY786146 * (as F. candida) [3] |
| 8 | Russia: Novgorod oblast | - | AY786148 (as F. candida) [3] |
| 9 | Armenia: Aragaz mtn, N40.43203 E44.23642 | ON220876 | ON228281 |
| 10 | Russia: Moscow oblast, Odintsovo district, raised bog, moss tussock, N55.733 E36.850 | ON220886-87 | ON228278-79 |
| 11 | Russia: Moscow oblast, collected by G. Dlussky in 2000 | - | AY786149 (as F. candida) [3] |
| 12 | Russia: Nizhniy, Novgorod oblast, Kerzhenskiy Natural Reserve, Vishenskoye swamp, N56.467 E44.500 | ON220875 | ON228280 |
| 13 | Russia: Sverdlovsk oblast, Revda | - | AY786151 (as F. candida) [3] |
| 14 | Russia: Sverdlovsk oblast, Yekaterinburg | - | AY786150 (as F. candida) [3] |
| 15 | Russia: Perm oblast, 30 km SSW of Kachkanar | - | AY786152 (as F. candida) [3] |
| 16 | China: Quinghai, Tibet | - | AY786157 (as F. candida) [3] |
| Formica candida | |||
| 17 | Kyrgyzstan: Alai Valley | - | AY786154 **, AY786153 [3], JX170887 [5] |
| 18 | Kyrgyzstan: Tian-Shan | - | AY786155-AY786156 [3] |
| Formica sp. | |||
| 19 | Russia: Novosibirsk oblast, Iskitim district, Listvyanskiy town, N54.450 E83.483 | ON220885 | - |
| 20 | Russia: Altai Republic, Ongudai town, N50.733 E86.133 | ON220880 | ON228273 |
| 21 | Russia: Buryatia Republic, Kizhinga town, N51.84639 E109.91278 | ON220883-84 | ON228270, ON228274 |
| 22 | Russia: Magadan oblast, 15 km from Yagodnoye town, bank of Sokhatina river, N62.517 E149.617 | ON220878 | ON228277 |
| 23 | Russia: Magadan oblast, 16 km from Yagodnoye town, bank of Detrin river, N62.750 E150.467 | ON220879 | ON228271 |
| 24 | Russia: Magadan oblast, Arman town, high floodplain, N59.667 E150.117 | ON220872 | ON228275 |
| 25 | Russia, Kamchatka oblast, N53.19 E158.48 | LT977377 (as F. candida) [4] | - |
| 26 | Russia: Primorye krai, Lazo town, Lazovka river, N43.38056 E133.89778 | ON220881 | ON228276 |
| 27 | Russia: Primorye krai, Sinii Utes town, N43.06222 E131.35472 | ON220882 | ON228272 |
| 28 | China: Harbin, HelongJiang | KX665031-32 (as F. candida) [7] | - |
| 29 | China: Yanbian, Jilin | KX665026-27 (as F. candida) [7] | - |
| 30 | South Korea | MT800217 (as F. candida) [8] | - |
| 31 | Xilin Gol League, Inner Mongolia | KX665022, KX665024 (as F. candida) [7] | - |
| 32 | Ulanqab, Inner Mongolia | KX665023, KX665025 (as F. candida) [7] | - |
| 33 | Helan Mountains, Inner Mongolia | KX665019-21 (as F. candida) [7] | - |
| 34 | Qinglong, Hebei | KX665028 (as F. candida) [7] | - |
| 35 | China: Hebei | HQ619704, HQ619710 (as F. candida) [9] | HQ651081, HQ651074 (as F. candida) [9] |
| 36 | Xiaowutai Mountain, Hebei | KX665029-30 (as F. candida) [7] | - |
| 37 | Zhong-Tiao Mountains, Shanxi | KX665033 (as F. candida) [7] | - |
| 38 | Liupan Mountains, Ningxia | KX665034 (as F. candida) [7] | |
| Outgroups | |||
| F. lemani Belarus: Vitebsk oblast, Braslav district, bank of Uklya lake, N55.60001 E27.333918 | ON220877 | ON228284 | |
| F. lemani Magadan, Arman town high river terrace, N59.867 E150.567 | ON220872 | ON228283 | |
| F. lemani China | - | HQ651082, HQ651086 [9] | |
| F. lemani Magadan | - | JX170882 [10] | |
| F. gagatoides China: Ningxia | - | HQ651073 [9] | |
| F. gagatoides China | KX665069, KX665071, KX665072 [7], HQ619703 | - | |
| F. gagatoides Canada | JN291934 | - | |
| F. gagatoides Norway | LT977396, LT977398 [4] | - | |
| F. gagatoides Russia: Magadan oblast, Arman town, N59.667 E150.117 | ON220874 | ON228282 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhigulskaya, Z.A.; Shekhovtsov, S.V.; Poluboyarova, T.V.; Berman, D.I. Formica picea and F. candida (Hymenoptera: Formicidae): Synonyms or Two Species? Diversity 2022, 14, 613. https://doi.org/10.3390/d14080613
Zhigulskaya ZA, Shekhovtsov SV, Poluboyarova TV, Berman DI. Formica picea and F. candida (Hymenoptera: Formicidae): Synonyms or Two Species? Diversity. 2022; 14(8):613. https://doi.org/10.3390/d14080613
Chicago/Turabian StyleZhigulskaya, Zoya A., Sergei V. Shekhovtsov, Tatiana V. Poluboyarova, and Daniil I. Berman. 2022. "Formica picea and F. candida (Hymenoptera: Formicidae): Synonyms or Two Species?" Diversity 14, no. 8: 613. https://doi.org/10.3390/d14080613