
DNA Barcoding and Distribution of Gastropods and Malacostracans in the Lower Danube Region

 , , , , and
, , , , and
Abstract
:
1. Introduction
2. Materials and Methods
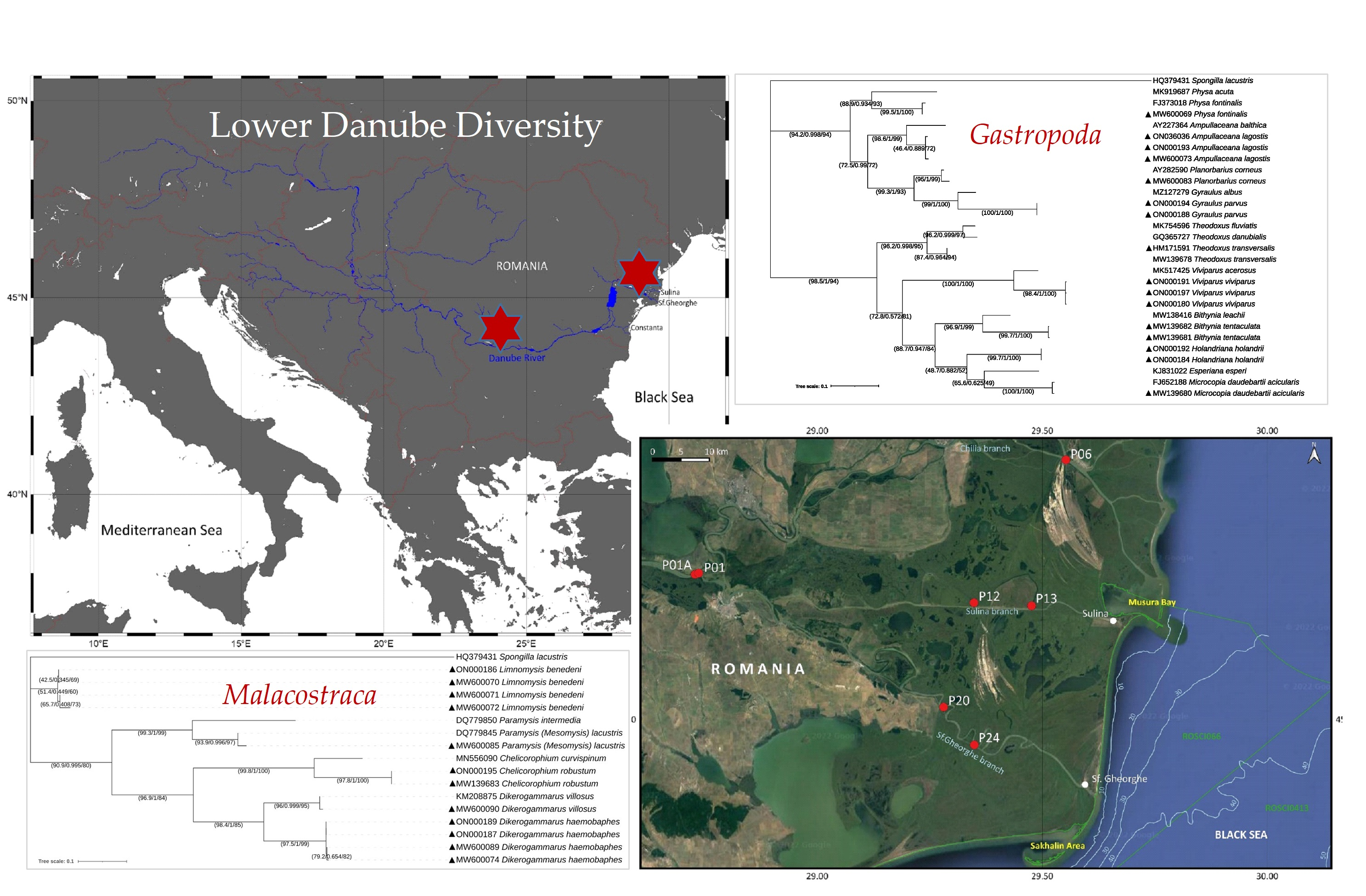
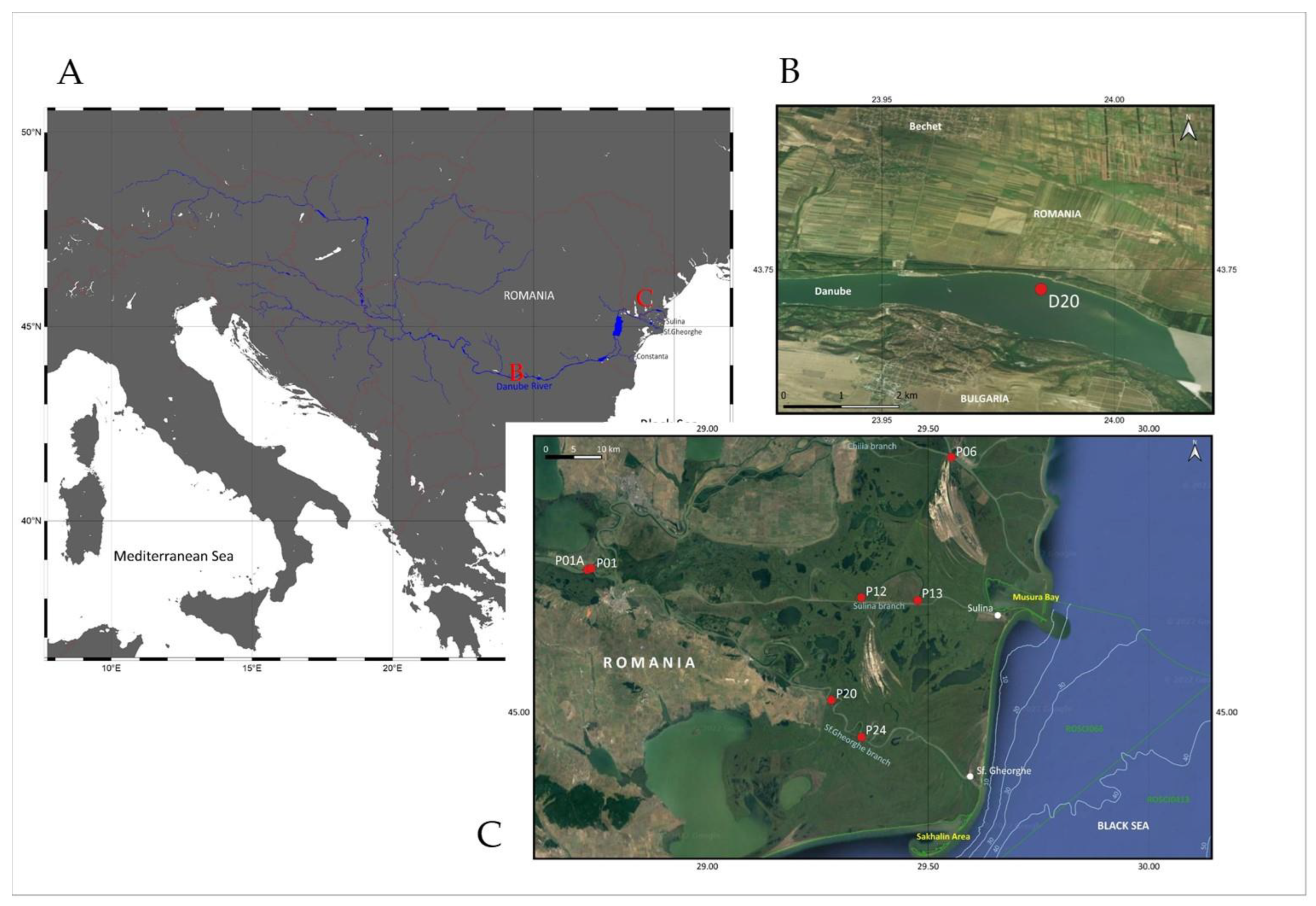
2.1. Study Area, Sampling and Sample Preparation
2.2. Morphological Identification
2.3. DNA Extraction, PCR Amplification and COI Gene Sequencing
2.4. Phylogenetic Analysis and Calculation of Intra- and Interspecific Genetic Distance
3. Results
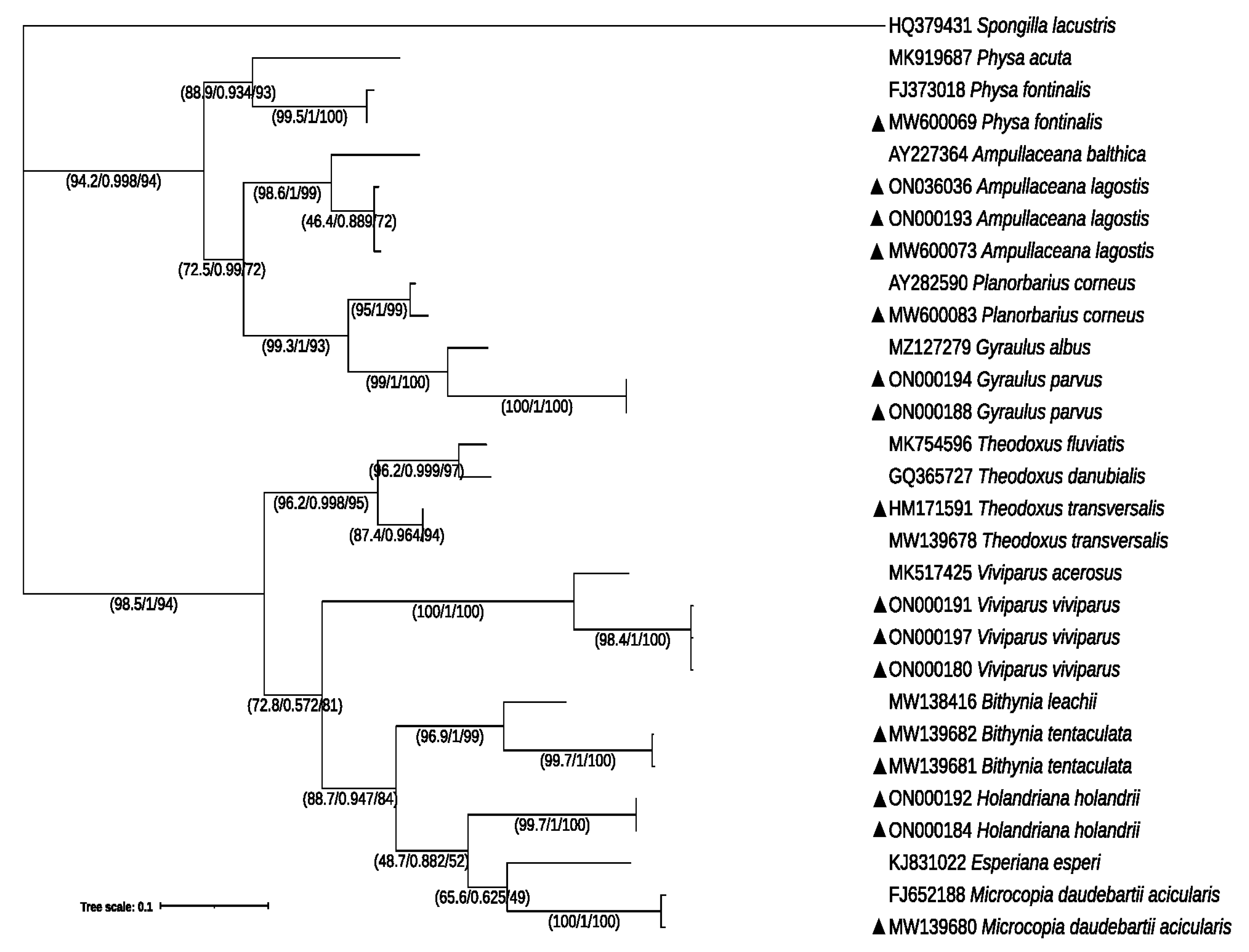
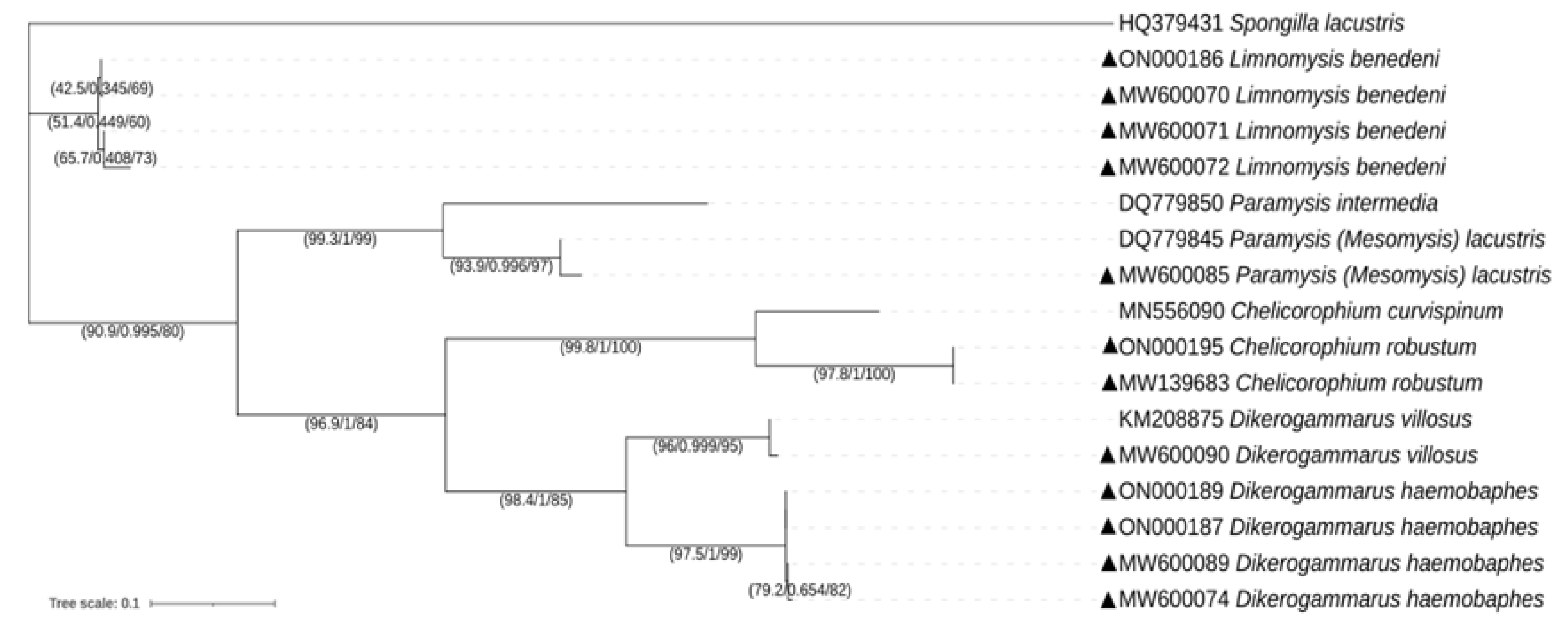
3.1. Molecular Identification and Phylogeny of Gastropods and Crustaceans
3.2. Distribution and Ecology of Gastropod and Crustacean Species along the Lower Danube Region
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Helmer, R.; Hespanhol, I. Water Pollution Control—A Guide to the Use of Water Quality Management Principles; United Nations Environment Programme, the Water Supply & Sanitation Collaborative Council and the World Health Organization WHO/UNEP: New York, NY, USA, 1997; Available online: https://www.who.int/water_sanitation_health/resourcesquality/wpcbegin.pdf (accessed on 25 April 2022).
- Gâştescu, P.; Ţuchiu, E. The Danube River in the Pontic Sector—Hydrologycal Regime. Water Resources and Wetlands. In Proceedings of the 1st Water and Wetlands Resources Conference, Tulcea, Romania, 14–16 September 2012; Gâştescu, P., Lewis, W., Jr., Breţcan, P., Eds.; Riscuri si Catastrofe: Cluj-Napoca, Romania, 2012; Volume 12, pp. 13–26, ISBN 978-606-605-038-8. [Google Scholar]
- ICPDR. Danube River Basin District Management Plan; ICPDR: Vienna, Austria, 2009. [Google Scholar]
- Oaie, G.; Secrieru, D.; Bondar, C.; Szobotka, Ş.; Duţu, L.; Manta, T. Lower Danube River: Characterization of sediments and pollutants. GeoEcoMarina 2015, 21, 19–34. [Google Scholar]
- Hanganu, J.; Grigoras, I.; Stefan, N.; Sarbu, I.; Zhmud, E.; Dubyna, D.; Menke, U.; Drost, H. Transboundary Vegetation Map of the Biosphere Reserve Danube Delta; Danube Delta Institute DDI, Institute for Inland Water Management and Waste Water Treatment RIZA, Danube Biosphere Reserve of the National Academy of Sciences of Ukraine DPA, I&KIB: Tulcea, Romania, 2002. [Google Scholar]
- Mee, L.D. The Black Sea in crisis: A need for concerted international action. Ambio 1992, 21, 278–286. [Google Scholar]
- Hutchinson, G.E. A Treatise on Limnology. In The Zoobenthos; John Wiley & Sons: New York, NY, USA, 1993; Volume 4, p. 944. [Google Scholar]
- Wallace, J.B.; Webster, J.R. The role of macroinvertebrates in stream ecosystem function. Annu. Rev. Entomol. 1996, 41, 115–139. [Google Scholar] [CrossRef]
- Clarke, K.D.; Knoechel, R.; Ryan, P.M. Influence of trophic role and life-cycle duration on timing and magnitude of benthic macroinvertebrate response to whole-lake enrichment. Can. J. Fish. Aquat. Sci. 1997, 54, 89–95. [Google Scholar] [CrossRef]
- Gomes, M.A.; Novelli, R.; Zalmon, I.R.; Souza, C.M. Malacological assemblages in sediments of eastern Brazilian continental shelf, coordinates 108 and 208 S, between Bahia and Espírito Santo State. Bios 2004, 12, 11–24. [Google Scholar]
- Tubić, B.; Simić, V.; Zorić, K.; Gačić, Z.; Atanacković, Z.; Csányi, B.; Paunović, M. Stream section types of the Danube River in Serbia according to the distribution of macroinvertebrates. Biologia 2013, 68, 294–302. [Google Scholar] [CrossRef]
- Rosemond, A.D.; Mullholland, P.J.; Ellwood, J.W. Top-down and bottom-up control of stream periphyton: Effects of nutrients and herbivores. Ecology 1993, 74, 1264–1280. [Google Scholar] [CrossRef]
- Brown, K.M.; Alexander, J.E.; Thorp, J.H. Differences in the ecology and distribution of lotic pulmonate and prosobranch gastropods. Am. Malacol. Bull. 1998, 14, 91–101. [Google Scholar]
- Frank, C.; Jungbluth, J.; Richnovszky, A. Die Mollusken der Donau vom Schwarzwald bis zum Schwarzen Meer; Akaprint: Budapest, Hungary, 1990; pp. 1–142. [Google Scholar]
- Moog, O.; Humpesch, U.H.; Konar, M. The distribution of benthic invertebrates along the Austrian stretch of the River Danube and its relevance as an indicator of zoogeographical and water quality patterns—Part 1. Arch. Hydroboil. Robiol. 1995, 9 Suppl. 101, 121–213. [Google Scholar] [CrossRef]
- Wesselingh, F.P.; Neubauer, T.A.; Anistratenko, V.V.; Vinarski, M.V.; Yanina, T.; ter Poorten, J.J.; Kijashko, P.; Albrecht, C.; Anistratenko, O.Y.; D’Hont, A.; et al. Mollusc species from the Pontocaspian region—An expert opinion list. ZooKeys 2019, 827, 31–124. [Google Scholar] [CrossRef] [Green Version]
- Glazier, D.S. Amphipoda. Reference Module in Earth Systems and Environmental Sciences; Elsevier: Amsterdam, The Netherlands, 2014; p. 49. [Google Scholar] [CrossRef]
- Roast, S.D.; Thompson, R.S.; Widdows, J.; Jones, M.B. Mysids and environmental monitoring: A case for their use in estuaries. Mar. Freshw. Res. 1998, 49, 827–832. [Google Scholar] [CrossRef]
- Alonso, Á.; De Lange, H.J.; Peeters, E.T.H.M. Development of a feeding behavioural bioassay using the freshwater amphipod Gammarus pulex and the Multispecies Freshwater Biomonitor. Chemosphere 2009, 75, 341–346. [Google Scholar] [CrossRef] [PubMed]
- Grabowski, M.; Bacela-Spychalska, K.; Pešić, V. Reproductive traits and conservation needs of the endemic gammarid Laurogammarus scutarensis (Schäferna, 1922) from the Skadar Lake system, Balkan Peninsula. Limnologica 2014, 47, 44–51. [Google Scholar] [CrossRef]
- Borza, P.; Huber, T.; Leitner, P.; Remund, N.; Graf, W. Success factors and future prospects of Ponto-Caspian peracarid (Crustacea: Malacostraca) invasions: Is “the worst over”? Biol. Invasions 2017, 19, 1517–1532. [Google Scholar] [CrossRef] [Green Version]
- Heino, J.; Louhi, P.; Muotka, T. Identifying the scales of variability in stream macroinvertebrate abundance, functional composition and assemblage structure. Freshw. Biol. 2004, 49, 1230–1239. [Google Scholar] [CrossRef]
- Seymour, M.; Deiner, K.; Altermatt, F. Scale and scope matter when explaining varying patterns of community diversity in riverine metacommunities. Basic Appl. Ecol. 2016, 17, 134–144. [Google Scholar] [CrossRef] [Green Version]
- Frézal, L.; Leblois, R. Four years of DNA barcoding: Current advances and prospects. Infect. Genet. Evol. 2008, 8, 727–736. [Google Scholar] [CrossRef] [Green Version]
- Macher, J.N.; Salis, R.K.; Blakemore, K.S.; Tollrian, R.; Matthaei, C.D.; Leese, F. Multiple-stressor effects on stream invertebrates: DNA barcoding reveals contrasting responses of cryptic mayfly species. Ecol. Indic. 2016, 61, 159–169. [Google Scholar] [CrossRef]
- Hebert, P.D.N.; Cywinska, A.; Ball, S.L. Biological identifications through DNA barcodes. Proc. R. Soc. Lond. B 2003, 270, 313–321. [Google Scholar] [CrossRef] [Green Version]
- Puillandre, N.; Cruaud, C.; Kantor, Y.I. Cryptic species in Gemmuloborsonia (Gastropoda: Conoidea). J. Mollus. Stud. 2009, 76, 11–23. [Google Scholar] [CrossRef] [Green Version]
- Sands, A.F.; Glöer, P.; Gürlek, M.E.; Albrecht, C.; Neubauer, T.A. A revision of the extant species of Theodoxus (Gastropoda, Neritidae) in Asia, with the description of three new species. Zoosyst. Evol. 2020, 96, 25–66. [Google Scholar] [CrossRef] [Green Version]
- Lipinskaya, T.; Radulovici, A.E. DNA barcoding of alien Ponto-Caspian amphipods from the Belarusian part of the Central European invasion corridor. Genome 2017, 60, 963–964. [Google Scholar] [CrossRef] [Green Version]
- Audzijonyte, A.; Daneliya, M.; Väinölä, R. Comparative phylogeography of Ponto-Caspian mysid crustaceans: Isolation and exchange among dynamic inland sea basins. Mol. Ecol. 2006, 15, 2969–2984. [Google Scholar] [CrossRef] [PubMed]
- Barco, A.; Claremont, M.; Reid, D.G.; Houart, R.; Bouchet, P.; Williams, S.T.; Cruaud, C.; Couloux, A.; Oliverio, M. A molecular phylogenetic framework for the Muricidae, a diverse family of carnivorous gastropods. Mol. Phylogenet. Evol. 2010, 56, 1025–1039. [Google Scholar] [CrossRef] [PubMed]
- Layton, K.K.; Martel, A.L.; Hebert, P.D.N. Patterns of DNA Barcode Variation in Canadian Marine Molluscs. PLoS ONE 2014, 9, e95003. [Google Scholar] [CrossRef] [Green Version]
- Graf, W.; Csányi, B.; Leitner, P.; Paunović, M.; Huber, T.; Szekeres, J.; Nagy, C.; Borza, P. Joint Danube Survey 3, Full Report on Macroinvertebrates; ICPDR, International Commission for the Protection of the Danube River: Sofia, Bulgaria, 2014; p. 87. Available online: www.icpdr.org (accessed on 20 May 2022).
- Glöer, P.; Sȋrbu, I. New freshwater molluscs species found in the Romanian fauna. Heldia 2005, 6, 229–238. [Google Scholar]
- Gomoiu, M.T.; Begun, T.; Opreanu, P.; Teaca, A. Present state of benthic ecosystem in Razelm-Sinoie Lagoon Complex (RSLC). In Proceedings of the 37th IAD Conference, The Danube River Basin in a Changing World, Chişinău, Moldova, 29 October–1 November 2008; pp. 108–112. [Google Scholar]
- Paraschiv, G.M.; Begun, T.; Teaca, A.; Bucur, M.; Tofan, L. New data about benthal populations of the Golovita and Zmeica lakes. J. Environ. Prot. Ecol. 2010, 11, 253–260. [Google Scholar]
- Begun, T.; Teacă, A.; Mureşan, M.; Pavel, A.B. Current state of the mollusc populations in the Razim-Sinoe Lagoon System. Anim. Sci. 2020, 63, 553–561. [Google Scholar]
- Begun, T. Complex Ecological Studies of Crustaceans Populations (Cumaceans and Mysidaceans) of the North Western Black Sea Coast. Ph.D. Thesis, University “Ovidius” Constanta, Constanța, Romania, 2006; p. 276. [Google Scholar]
- Paraschiv, G.M.; Schroder, V.; Samargiu, M.D.; Sava, D. Ecological study of zoobenthos communities from the Matita and Merhei lakes (Danube Delta). Res. J. Agric. Sci. 2007, 39, 489–498. [Google Scholar]
- Graf, W.; Csányi, B.; Leitner, P.; Paunović, M.; Janeček, B.; Šporka, F.; Chiriac, G.; Stubauer, I.; Ofenböck, T. Joint Danube Survey 2, Report on Macroinvertebrates; ICPDR International Commission for the Protection of the Danube River: Vienna, Austria, 2008; p. 87. [Google Scholar]
- Borza, P.; Csányi, B.; Paunović, M. Corophiids (Amphipoda, Corophioidea) of the river Danube—The results of a longitudinal urvey. Crustaceana 2010, 83, 839–849. [Google Scholar]
- Stoica, C.; Gheorghe, S.; Petre, J.; Lucaciu, I.; Nita-Lazar, M. Tools for assessing Danube Delta systems with macro invertebrates. Environ. Eng. Manag. J. 2014, 13, 2243–2252. [Google Scholar] [CrossRef]
- Cristescu, M.E.; Hebert, P.D.; Onciu, T.M. Phylogeography of Ponto-Caspian crustaceans: A benthic–planktonic comparison. Mol. Ecol. 2003, 12, 985–996. [Google Scholar] [CrossRef] [PubMed]
- Rewicz, T.; Wattier, R.; Grabowski, M.; Rigaud, T.; Bącela-Spychalska, K. Out of the Black Sea: Phylogeography of the invasive killer shrimp Dikerogammarus villosus across Europe. PLoS ONE 2015, 10, e0118121. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jażdżewska, A.M.; Rewicz, T.; Mamos, T.; Wattier, R.; Bącela-Spychalska, K.; Grabowski, M. Cryptic diversity and mtDNA phylogeography of the invasive demon shrimp, Dikerogammarus haemobaphes (Eichwald, 1841), in Europe. NeoBiota 2020, 57, 53–86. [Google Scholar] [CrossRef]
- Cristescu, M.E.A.; Hebert, P.D.N. The ‘Crustacean Seas’—An evolutionary perspective on the Ponto-Caspian peracarids. Can. J. Fish. Aquat. Sci. 2005, 62, 505–551. [Google Scholar] [CrossRef]
- Ross, K.S.; Haites, N.E.; Kelly, K.F. Repeated freezing and thawing of peripheral blood and DNA in suspension: Effects on DNA yield and integrity. J. Med. Genet. 1990, 27, 569–570. [Google Scholar] [CrossRef] [Green Version]
- SR EN ISO 10870; Water Quality—Guidelines for the Selection of Sampling Methods and Devices for Benthic Macroinvertebrates in Fresh Waters. ISO: Geneva, Switzerland, 2012.
- Grossu, A.V. Mollusca—Gastropoda Prosobranchia si Opistobranchia; Fauna Republicii Populare Române: Bucharest, Romana, 1956; Volume 3. [Google Scholar]
- Cărăuşu, S.; Dobreanu, E.; Manolache, C. Fauna Republicii Populare Romȋne. Crustacea. Volumul IV. Fascicula 4. Amphipoda. Forme Salmastre şi de apă Dulce; Editura Acedemiei Republicii Populare Romȋne: Bucharest, Romana, 1955; pp. 1–401. [Google Scholar]
- Băcescu, M. Crustacea: Mysidacea; Editura Academiei Republicii Populare Romîne: Bucureşti, Romania, 1954; Volume 126, p. 52. [Google Scholar]
- Iancu, L.; Carter, D.O.; Junkins, E.N.; Purcarea, C. Using bacterial and necrophagous insect dynamics for postmortem interval estimation during cold season: Novel case study in Romania. Forensic Sci. Int. 2015, 254, 106–117. [Google Scholar] [CrossRef]
- Folmer, O.; Black, M.; Hoeh, W.; Lutz, R.; Vrijenhoek, R. DNA primers for amplification of mitochondrial cytochrome c oxidase subunit I from diverse metazoan invertebrates. Mol. Mar. Biol. Biotechnol. 1994, 3, 294–299. [Google Scholar]
- Available online: https://blast.ncbi.nlm.nih.gov (accessed on 28 April 2022).
- Leray, M.; Yang, J.Y.; Meyer, C.P.; Mills, S.C.; Agudelo, N.; Ranwez, V.; Machida, R.J. A new versatile primer set targeting a short fragment of the mitochondrial COI region for metabarcoding metazoan diversity: Application for characterizing coral reef fish gut contents. Front. Zool. 2013, 10, 34. [Google Scholar] [CrossRef] [Green Version]
- Wilkinson, S.P.; Davy, S.K.; Bunce, M.; Stat, M. Taxonomic identification of environmental DNA with informatic sequence classification trees. PeerJ Prepr. 2018, 6, e26812v1. [Google Scholar] [CrossRef]
- Edgar, R.C. MUSCLE: Multiple sequence alignment with high accuracy and high throughput. Nucleic Acids Res. 2004, 32, 1792–1797. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Trifinopoulos, J.; Nguyen, L.T.; von Haeseler, A.; Minh, B.Q. W-IQ-TREE: A fast online phylogenetic tool for maximum likelihood analysis. Nucleic Acids Res. 2016, 44, W232–W235. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Letunic, I.; Bork, P. Interactive Tree of Life (iTOL) v5: An online tool for phylogenetic tree display and annotation. Nucleic Acids Res. 2021, 49, W293–W296. [Google Scholar] [CrossRef] [PubMed]
- Kimura, M. A simple method for estimating evolutionary rates of base substitutions through comparative studies of nucleotide sequences. J. Mol. Evol. 1980, 16, 111–120. [Google Scholar] [CrossRef]
- Tamura, K.; Stecher, G.; Kumar, S. MEGA 11: Molecular Evolutionary Genetics Analysis Version 11. Mol. Biol. Evol. 2021, 38, 3022–3027. [Google Scholar] [CrossRef]
- Radulovici, A.E.; Archambault, P.; Dufresne, F. DNA Barcodes for Marine Biodiversity: Moving Fast Forward? Diversity 2010, 2, 450–472. [Google Scholar] [CrossRef] [Green Version]
- Hebert, P.D.N.; Penton, E.H.; Burns, J.M.; Janzen, D.H.; Hallwachs, W. Ten Species in One: DNA Barcoding Reveals Cryptic Species in the Neotropical Skipper Butterfly Astraptes fulgerator. Proc. Natl. Acad. Sci. USA 2004, 101, 14812–14817. [Google Scholar] [CrossRef] [Green Version]
- Sweeney, B.W.; Battle, J.M.; Jackson, J.K.; Dapkey, T. Can DNA Barcodes of Stream Macroinvertebrates Improve Descriptions of Community Structure and Water Quality? J. N. Am. Benthol. Soc. 2011, 30, 195–216. [Google Scholar] [CrossRef]
- Hebert, P.D.N.; Stoeckle, M.Y.; Zemlak, T.S.; Francis, C.M. Identification of Birds through DNA Barcodes. PLoS Biol. 2004, 2, e312. [Google Scholar] [CrossRef] [Green Version]
- Borges, L.; Hollatz, C.; Lobo, J.; Cunha, A.M.; Vilela, A.P.; Calado, G.; Coelho, R.; Costa, A.C.; Ferreira, M.S.G.; Costa, M.H.; et al. With a little help from DNA barcoding: Investigating the diversity of Gastropoda from the Portuguese coast. Sci. Rep. 2016, 6, 20226. [Google Scholar] [CrossRef] [Green Version]
- Lawton, S.P.; Allan, F.; Hayes, P.M.; Smit, N.J. DNA barcoding of the medically important freshwater snail Physa acuta reveals multiple invasion events into Africa. Acta Trop. 2018, 188, 86–92. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Buršić, M.; Iveša, L.; Jaklin, A.; Arko Pijevac, M.; Kučinić, M.; Štifanić, M.; Neal, L.; Bruvo Mađarić, B. DNA Barcoding of Marine Mollusks Associated with Corallina officinalis Turfs in Southern Istria (Adriatic Sea). Diversity 2021, 13, 196. [Google Scholar] [CrossRef]
- Costa, F.; deWaard, J.R.; Boutillier, J.; Ratnasingham, S.; Dooh, R.T.; Hajibabaei, M.; Hebert, P.D.N. Biological identifications through DNA barcodes: The case of the Crustacea. Can. J. Fish. Aquat. Sci. 2007, 64, 272–295. [Google Scholar] [CrossRef]
- Costa, F.; Henzler, C.; Lunt, D.; Whiteley, N.M.; Rock, J. Probing marine Gammarus (Amphipoda) taxonomy with DNA barcodes. Syst. Biodivers. 2009, 7, 365–379. [Google Scholar] [CrossRef]
- Audzijonyte, A.; Wittmann, K.J.; Ovcarenko, I.; Väinölä, R. Invasion phylogeography of the Ponto-Caspian crustacean Limnomysis benedeni dispersing across Europe. Divers. Distrib. 2009, 15, 346–355. [Google Scholar] [CrossRef]
- Zhang, D.-X.; Hewitt, G.M. Assessment of the universality and utility of a set of conserved mitochondrial primers in insects. Insect Mol. Biol. 1997, 6, 143–150. [Google Scholar] [CrossRef]
- Cox, A.J.; Hebert, P.D.N. Colonization, extinction and phylogeographic patterning in a freshwater crustacean. Mol. Ecol. 2001, 10, 371–386. [Google Scholar] [CrossRef]
- Wares, J.P.; Cunningham, C.W. Phylogeography and historical ecology of the North Atlantic intertidal. Evolution 2001, 12, 2455–2469. [Google Scholar] [CrossRef]
- Rysiewska, A.; Hofman, S.; Osikowski, A.; Beran, L.; Pešić, V.; Falniowski, A. Viviparus mamillatus (Küster, 1852), and partial congruence between the morphology-, allozyme- and DNA-based phylogeny in European Viviparidae (Caenogastropoda: Architaenioglossa). Folia Malacol. 2019, 27, 43–51. [Google Scholar] [CrossRef] [Green Version]
- Beran, L.; Horsák, M.; Hofman, S. First records of Viviparus acerosus (Bourguignat, 1862) (Gastropoda: Viviparidae) from the Czech Republic outside its native range. Folia Malacol. 2019, 27, 223–229. [Google Scholar] [CrossRef] [Green Version]
- Lorencová, E.; Beran, L.; Nováková, M.; Horsáková, V.; Rowson, B.; Jaroslav, Č.; Hlaváč, J.Č.; Nekola, J.C.; Horsák, M. Invasion at the population level: A story of the freshwater snails Gyraulus parvus and G. laevis. Hydrobiol. 2021, 848, 4661–4671. [Google Scholar] [CrossRef]
- Pjatakova, G.M.; Tarasov, A.G. Caspian Sea amphipods: Biodiversity, systematic position and ecological peculiarities of some species. Int. J. Salt Lake Res. 1996, 5, 63–79. [Google Scholar] [CrossRef]
- Müller, J.C.; Schramm, S.; Seitz, A. Genetic and morphological differentiation of Dikerogammarus invaders and their invasion history in Central Europe. Freshw. Biol. 2002, 47, 2039–2048. [Google Scholar] [CrossRef]
- Mamos, T.; Grabowski, M.; Rewicz, T.; Bojko, J.; Strapagiel, D.; Burzyński, A. Mitochondrial Genomes, Phylogenetic Associations, and SNP Recovery for the Key Invasive Ponto-Caspian Amphipods in Europe. Int. J. Mol. Sci. 2021, 22, 10300. [Google Scholar] [CrossRef] [PubMed]
- Marescaux, J.; Latli, A.; Lorquet, J.; Virgo, J.; Van Doninck, K.; Beisel, J.N. Benthic macro-invertebrate fauna associated with Dreissena mussels in the Meuse River: From incapacitating relationships to facilitation. Aquat. Ecol. 2016, 50, 15–28. [Google Scholar] [CrossRef]
- ICPDR. Joint Danube Survey 2. Final Scientific Report; ICPDR: Vienna, Austria, 2008. [Google Scholar]
- Martinovic-Vitanovic, V.; Raković, M.; Popović, N.; Kalafatic, V. Qualitative study of Mollusca communities in the Serbian Danube stretch (river km 1260–863.4). Biologia 2013, 68, 112–130. [Google Scholar] [CrossRef] [Green Version]
- ICPDR. Joint Danube Survey 1. Ecological Status Characterization. Macrozoobenthos; ICPDR: Vienna, Austria, 2002. [Google Scholar]
- Surugiu, V.; Mustaţă, G.; Hârţăscu, M. Contributions to the qualitative and quantitative study of the macrozoobenthos from the Danube—Black Sea Canal. Stud. Şi Cercet. Biol. Univ. Din Bacău 2004, 9, 75–80. [Google Scholar]
- Georgiev, D.; Hubenov, Z. Freshwater snails (Mollusca: Gastropoda) of Bulgaria: An updated annotated checklist. Folia Malacol. 2013, 21, 237–263. [Google Scholar] [CrossRef] [Green Version]
- Sȋrbu, I.; Sȋrbu, M.; Benedek, A. The freshwater molluskfauna from Banat (Romania). Trav. du Mus. Natl. d’Hist. Nat. Grigore Antipa 2010, 53, 21–43. [Google Scholar] [CrossRef]
- Solymos, P.; Feher, Z. Theodoxus transversalis (C. Pfeiffer1828). In IUCN 2011. IUCN Red List of Threatened Species. Version 2011. 2011. Available online: www.iucnredlist.org (accessed on 18 May 2022).
- Novakovic, B.; Markovic, V.; Tomović, J. Distribution of the snail Amphimelania holandrii Pfeiffer, 1828 (Melanopsidae; Gastropoda) in Serbia in the 2009–2012 period. Water Res. Manag. J. Belgrade 2013, 3, 21–26. [Google Scholar]
- ICPDR. Joint Danube Survey 3. Full Report on Macroinvertebrates; ICPDR: Vienna, Austria, 2014. [Google Scholar]
- Muskó, I.B. The life history of Dikerogammarus haemobaphes (Eichw.) (Crustacea: Amphipoda) living on macrophytes in Lake Balaton (Hungary). Arch. Für Hydrobiol. 1993, 127, 227–238. [Google Scholar] [CrossRef]
- Boets, P.; Lock, K.; Messiaen, M.; Goethals, P.L.M. Combining data-driven methods and lab studies to analyse the ecology of Dikerogammarus villosus. Ecol. Inform. 2010, 5, 133–139. [Google Scholar] [CrossRef]
- Clinton, K.E.; Mathers, K.L.; Constable, D.; Gerrard, C.; Wood, P.J. Substrate preferences of coexisting invasive amphipods, Dikerogammarus villosus and Dikerogammarus haemobap-hes, under field and laboratory conditions. Biol. Invasions 2018, 20, 2187–2196. [Google Scholar] [CrossRef] [Green Version]
- Lipták, B.; Šporka, F.; Necpálová, K.; Stloukal, E. First record of Ponto-Caspian amphipod Corophium curvispinum in Slovakside of the Danube River. Folia Faun. Slovaca 2012, 17, 183–186. [Google Scholar]
- Lipták, B. Non-indigenous invasivefreshwater crustaceans (Crustacea: Malacostraca) in Slovakia. Water Res. Manag. 2013, 3, 21–31. [Google Scholar]
- Lipinskaya, T.; Makaranka, A.; Razlutskij, V.; Semenchenko, V. First records of the alien amphipod Dikerogammarus haemobaphes (Eichwald, 1841) in the Neman River basin (Belarus). BioInvasions Rec. 2021, 10, 319–325. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Station | Year of Sampling | Coordinates | Depth (m) | Substrate Type | |
|---|---|---|---|---|---|
| Lat. (α) | Long. (λ) | ||||
| P01 | 2019, 2020, 2021 | 45°13′36.23′′ | 28°43′57.49′′ | 24.0 | MS |
| P01A | 2019, 2020, 2021 | 45°13′36.23′′ | 28°43′57.49′′ | 24.0 | MS |
| P06 | 2019, 2020, 2021 | 45°24′19.16′′ | 29°33′12.71′′ | 6.8 | SM |
| P12 | 2019, 2020, 2021 | 45°10′53.65′′ | 29.20′47.84′′ | 4.7 | MSM |
| P13 | 2019, 2020, 2021 | 45°10′35.34′′ | 29°28′28.66′′ | 5.1 | MSM |
| P20 | 2019, 2020, 2021 | 45°01′10.2′′ | 29°16′36.64′′ | 13.8 | S |
| P24 | 2019, 2020, 2021 | 44°57′33.80′′ | 29°20′48.80′′ | 19.5 | M |
| D21 | 2019, 2021 | 43°44′13.0′′ | 23°59′4.3′′ | 3.7 | AV |
| Species/Orders | Sampling Sites | Depth (m) | Type of Substrate | |||||||
|---|---|---|---|---|---|---|---|---|---|---|
| P01 | P01A | P06 | P12 | P13 | P20 | P24 | D20 | |||
| Gastropoda | ||||||||||
| Caenogastropoda | ||||||||||
| Holandriana holandrii | 2021 | 3.7 | AV | |||||||
| Microcolpia daudebartii acicularis | 2019 | 2019 | 4.7 | MS | ||||||
| 2021 | 2021 | 24 | MSM | |||||||
| Architaenioglossa | ||||||||||
| Viviparus viviparus | 2020 | 2020 | 4.7 | MS | ||||||
| 2021 | 2021 | 24 | MSM | |||||||
| Littorinimorpha | ||||||||||
| Bithynia tentaculata | 2020 | 2020 | 5.1 | MS | ||||||
| 2021 | 2021 | 24 | MSM | |||||||
| Cycloneritida | ||||||||||
| Theodoxus transversalis | 2019 | 3.7 | AV | |||||||
| 2021 | ||||||||||
| Hygrophila | ||||||||||
| Ampullaceana lagotis | 2020 | 6.8 | SM | |||||||
| 2021 | ||||||||||
| Physa fontinalis | 2020 | 5.1 | MSM | |||||||
| 2021 | ||||||||||
| Planorbarius corneus | 2020 | 4.7 | MSM | |||||||
| 2021 | ||||||||||
| Gyraulus parvus | 2021 | 3.7 | AV | |||||||
| Malacostraca | ||||||||||
| Amphipoda | ||||||||||
| Chelicorophium robustum | 2019 | 2019 | 2019 | 2019 | 4.7 | MS | ||||
| 2021 | 2021 | 2021 | 2021 | 6.8 | MSM | |||||
| 24 | ||||||||||
| Dikerogammarus haemobaphes | 2019 | 2019 | 2019 | 2019 | 2019 | 4.7 | MS | |||
| 2020 | 2021 | 2020 | 2020 | 2020 | 5.1 | MSM | ||||
| 2021 | 2021 | 2021 | 2021 | 6.8 | ||||||
| 24 | ||||||||||
| Dikerogammarus villosus | 2020 | 2021 | 2020 | 2020 | 2020 | 2020 | 4.7 | MS | ||
| 2021 | 2021 | 2021 | 2021 | 2021 | 5.1 | MSM | ||||
| 13.8 | ||||||||||
| 19.5 | ||||||||||
| 24 | ||||||||||
| Mysida | ||||||||||
| Limnomysis benedeni | 2020 | 5.1 | MSM | |||||||
| 2021 | ||||||||||
| Paramysis (Mesomysis) lacustris | 2020 | 2020 | 4.7 | MSM | ||||||
| 2021 | 2021 | 5.1 | ||||||||
| 1. | 2. | 3. | 4. | 5. | 6. | 7. | 8. | 9. | 10. | 11. | 12. | 13. | 14. | 15. | 16. | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1. ON000184 H. holandrii | - | |||||||||||||||
| 2. ON000192 H. holandrii | 0.000 | - | ||||||||||||||
| 3.MW139680 M. daudebartii acicularis | 0.205 | 0.207 | - | |||||||||||||
| 4. ON000180 V. viviparus | 0.281 | 0.282 | 0.241 | - | ||||||||||||
| 5. ON000191 V. viviparus | 0.275 | 0.276 | 0.241 | 0.003 | - | |||||||||||
| 6. ON000197 V. viviparus | 0.277 | 0.278 | 0.240 | 0.003 | 0.003 | - | ||||||||||
| 7. MW139681 B. tentaculata | 0.235 | 0.236 | 0.240 | 0.274 | 0.268 | 0.271 | - | |||||||||
| 8. MW139682 B. tentaculata | 0.242 | 0.241 | 0.243 | 0.278 | 0.268 | 0.278 | 0.003 | - | ||||||||
| 9. MW139678 T. transversalis | 0.240 | 0.238 | 0.211 | 0.263 | 0.259 | 0.261 | 0.224 | 0.225 | - | |||||||
| 10. ON000193 A. lagotis | 0.327 | 0.317 | 0.327 | 0.347 | 0.337 | 0.345 | 0.319 | 0.321 | 0.277 | - | ||||||
| 11. ON036036 A. lagotis | 0.299 | 0.294 | 0.307 | 0.332 | 0.329 | 0.329 | 0.307 | 0.307 | 0.257 | 0.004 | - | |||||
| 12. MW600073 A. lagotis | 0.315 | 0.305 | 0.306 | 0.333 | 0.330 | 0.332 | 0.308 | 0.306 | 0.258 | 0.005 | 0.007 | - | ||||
| 13. MW600069 P. fontinalis | 0.365 | 0.352 | 0.355 | 0.315 | 0.312 | 0.312 | 0.298 | 0.298 | 0.272 | 0.180 | 0.180 | 0.185 | - | |||
| 14. MW600083 P. corneus | 0.345 | 0.329 | 0.302 | 0.305 | 0.302 | 0.304 | 0.321 | 0.318 | 0.273 | 0.173 | 0.172 | 0.175 | 0.202 | - | ||
| 15. ON000188 G. parvus | 0.362 | 0.345 | 0.333 | 0.356 | 0.347 | 0.353 | 0.299 | 0.307 | 0.295 | 0.215 | 0.207 | 0.208 | 0.218 | 0.164 | - | |
| 16. ON000194 G. parvus | 0.362 | 0.362 | 0.350 | 0.378 | 0.372 | 0.378 | 0.316 | 0.320 | 0.316 | 0.232 | 0.219 | 0.226 | 0.230 | 0.175 | 0.000 | - |
| 1. | 2. | 3. | 4. | 5. | 6 | 7. | 8. | 9. | 10. | 11. | 12. | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1. MW139683 C.robustum | - | |||||||||||
| 2. ON000195 C. robustum | 0.000 | - | ||||||||||
| 3. MW600074 D. haemobaphes | 0.332 | 0.342 | - | |||||||||
| 4. MW600089 D. haemobaphes | 0.331 | 0.336 | 0.003 | - | ||||||||
| 5. ON000187 D. haemobaphes | 0.337 | 0.332 | 0.004 | 0.002 | - | |||||||
| 6. ON000189 D. haemobaphes | 0.338 | 0.331 | 0.002 | 0.002 | 0.000 | - | ||||||
| 7. MW600090 D. villosus | 0.322 | 0.332 | 0.204 | 0.199 | 0.205 | 0.203 | - | |||||
| 8. MW600070 L. benedeni | 0.350 | 0.373 | 0.350 | 0.347 | 0.358 | 0.349 | 0.326 | - | ||||
| 9. MW600071 L. benedeni | 0.348 | 0.370 | 0.353 | 0.350 | 0.361 | 0.353 | 0.333 | 0.008 | - | |||
| 10. MW600072 L. benedeni | 0.374 | 0.400 | 0.380 | 0.383 | 0.389 | 0.381 | 0.355 | 0.025 | 0.020 | - | ||
| 11. ON000186 L. benedeni | 0.350 | 0.370 | 0.348 | 0.344 | 0.355 | 0.346 | 0.333 | 0.002 | 0.006 | 0.027 | - | |
| 12. MW600085 P. lacustris | 0.366 | 0.389 | 0.336 | 0.333 | 0.347 | 0.348 | 0.346 | 0.310 | 0.306 | 0.338 | 0.305 | - |
| Species | Density (Individuals/m2) | |||||||
|---|---|---|---|---|---|---|---|---|
| P01 * | P01A * | P06 * | P12 * | P13 * | P20 * | P24 * | D20 ** | |
| Gastropoda | ||||||||
| M. daudebartii acicularis | 20 | 20 | ||||||
| V. viviparus | 20 | 20 | ||||||
| B. tentaculata | 10 | 10 | ||||||
| A. lagotis | 10 | |||||||
| P. fontinalis | 10 | |||||||
| P. corneus | 30 | |||||||
| H. holandrii | 2 | |||||||
| T. transversalis | 3 | |||||||
| G. parvus | 2 | |||||||
| Malacostraca | ||||||||
| C. robustum | 10 | 30 | 20 | 420 | ||||
| D. haemobaphes | 10 | 130 | 210 | 20 | 300 | 30 | ||
| D. villosus | 40 | 110 | 10 | 30 | 10 | 20 | ||
| L. benedeni | 10 | |||||||
| P. lacustris | 10 | 30 | ||||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Menabit, S.; Begun, T.; Teacă, A.; Mureşan, M.; Lavin, P.; Purcarea, C. DNA Barcoding and Distribution of Gastropods and Malacostracans in the Lower Danube Region. Diversity 2022, 14, 533. https://doi.org/10.3390/d14070533
Menabit S, Begun T, Teacă A, Mureşan M, Lavin P, Purcarea C. DNA Barcoding and Distribution of Gastropods and Malacostracans in the Lower Danube Region. Diversity. 2022; 14(7):533. https://doi.org/10.3390/d14070533
Chicago/Turabian StyleMenabit, Selma, Tatiana Begun, Adrian Teacă, Mihaela Mureşan, Paris Lavin, and Cristina Purcarea. 2022. "DNA Barcoding and Distribution of Gastropods and Malacostracans in the Lower Danube Region" Diversity 14, no. 7: 533. https://doi.org/10.3390/d14070533