
Diversity and Big Tree Patterns in the Brazilian Amazon

 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Area
2.2. Data Analysis
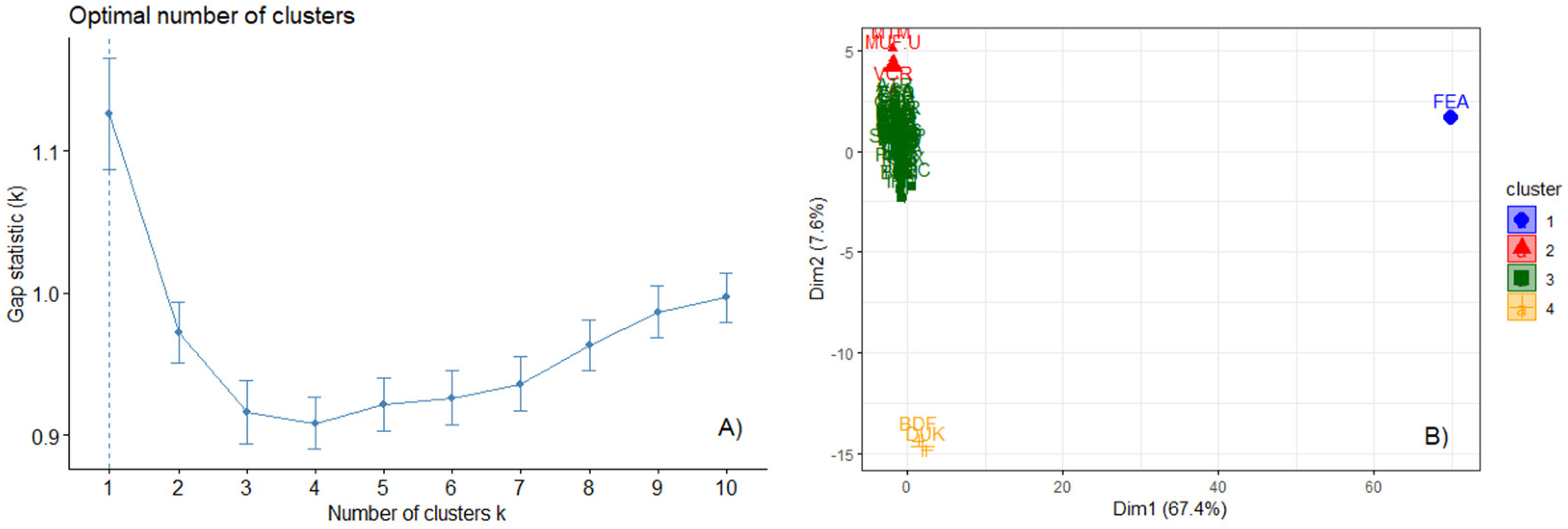
3. Results
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Enquist, B.J.; Abraham, A.J.; Harfoot, M.B.J.; Malhi, Y.; Doughty, C.E. The Megabiota Are Disproportionately Important for Biosphere Functioning. Nat. Commun. 2020, 11, 699. [Google Scholar] [CrossRef] [PubMed]
- Schweiger, A.H.; Svenning, J.-C. Analogous Losses of Large Animals and Trees, Socio-Ecological Consequences, and an Integrative Framework for Rewilding-Based Megabiota Restoration. People Nat. 2020, 2, 29–41. [Google Scholar] [CrossRef]
- Harris, D.J.; Ndolo Ebika, S.T.; Sanz, C.M.; Madingou, M.P.N.; Morgan, D.B. Large Trees in Tropical Rain Forests Require Big Plots. Plants People Planet 2021, 3, 282–294. [Google Scholar] [CrossRef]
- Ali, A.; Wang, L.-Q. Big-Sized Trees and Forest Functioning: Current Knowledge and Future Perspectives. Ecol. Indic. 2021, 127, 107760. [Google Scholar] [CrossRef]
- Poorter, L.; Craven, D.; Jakovac, C.; van der Sande, M.; Amissah, L.; Bongers, F.; Chazdon, R.; Farrior, C.; Kambach, S.; Meave, J.; et al. Multidimensional Tropical Forest Recovery. Science 2021, 374, 1370–1376. [Google Scholar] [CrossRef]
- Réjou-Méchain, M.; Fayolle, A.; Nasi, R.; Gourlet-Fleury, S.; Doucet, J.-L.; Gallye, M.; Hubertb, D.; Pasquier, A.; Billandb, A. Detecting Large-Scale Diversity Patterns in Tropical Trees Can We Trust Commercial Forest Inventories? For. Ecol. Manag. 2011, 261, 9. [Google Scholar] [CrossRef]
- Tuomisto, H.; Van doninck, J.; Ruokolainen, K.; Moulatlet, G.M.; Figueiredo, F.O.G.; Sirén, A.; Cárdenas, G.; Lehtonen, S.; Zuquim, G. Discovering Floristic and Geoecological Gradients across Amazonia. J. Biogeogr. 2019, 46, 1734–1748. [Google Scholar] [CrossRef]
- Baker, T.R.; Phillips, O.L.; Malhi, Y.; Almeida, S.; Arroyo, L.; Fiore, A.D.; Erwin, T.; Higuchi, N.; Killeen, T.J.; Laurance, S.G.; et al. Increasing Biomass in Amazonian Forest Plots. Philos. Trans. R. Soc. Lond. Ser. B Biol. Sci. 2004, 359, 353–365. [Google Scholar] [CrossRef]
- ter Steege, H.; Pitman, N.; Phillips, O.; Chave, J.; Sabatier, D.; Duque, A.; Molino, J.-F.; Prévost, M.-F.; Spichiger, R.; Castellanos, H.; et al. Continental-Scale Patterns of Canopy Tree Composition and Function Across Amazonia. Nature 2006, 443, 444–447. [Google Scholar] [CrossRef]
- Malhi, Y.; Aragão, L.E.O.C.; Galbraith, D.; Huntingford, C.; Fisher, R.; Zelazowski, P.; Sitch, S.; McSweeney, C.; Meir, P. Exploring the Likelihood and Mechanism of a Climate-Change-Induced Dieback of the Amazon Rainforest. Proc. Natl. Acad. Sci. USA 2009, 106, 20610–20615. [Google Scholar] [CrossRef] [Green Version]
- Lewis, S.L.; Sonké, B.; Sunderland, T.; Begne, S.K.; Lopez-Gonzalez, G.; van der Heijden, G.M.F.; Phillips, O.L.; Affum-Baffoe, K.; Baker, T.R.; Banin, L.; et al. Above-Ground Biomass and Structure of 260 African Tropical Forests. Philos. Trans. R. Soc. B 2013, 368, 20120295. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bastin, J.; Rutishauser, E.; Kellner, J.R.; Saatchi, S.; Pélissier, R.; Hérault, B.; Slik, F.; Bogaert, J.; De Cannière, C.; Marshall, A.R.; et al. Pan-tropical Prediction of Forest Structure from the Largest Trees. Glob. Ecol. Biogeogr. 2018, 27, 1366–1383. [Google Scholar] [CrossRef]
- Ter Steege, H.; Pitman, N.; Sabatier, D.; Baraloto, C.; Salomão, R.; Guevara Andino, J.; Phillips, O.; Castilho, C.; Magnusson, W.; Molino, J.-F.; et al. Hyperdominance in the Amazonian Tree Flora. Science 2013, 342, 1243092. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morrone, J. Biogeographical Regionalisation of the Neotropical Region. Zootaxa 2014, 3782, 1–110. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Souza, L.A.S.; Eisenlohr, P.V. Drivers of Floristic Variation in Biogeographic Transitions: Insights from the Ecotone between the Largest Biogeographic Domains of South America. Acta Bot. Bras. 2020, 34, 155–166. [Google Scholar] [CrossRef]
- Pérez-Luque, A.J.; Benito, B.M.; Bonet-García, F.J.; Zamora, R. Ecological Diversity within Rear-Edge: A Case Study from Mediterranean Quercus Pyrenaica Willd. Forests 2020, 12, 10. [Google Scholar] [CrossRef]
- Draper, F.C.; Costa, F.R.C.; Arellano, G.; Phillips, O.L.; Duque, A.; Macía, M.J.; ter Steege, H.; Asner, G.P.; Berenguer, E.; Schietti, J.; et al. Amazon Tree Dominance across Forest Strata. Nat. Ecol. Evol. 2021, 5, 757–767. [Google Scholar] [CrossRef]
- Kearsley, E.; Hufkens, K.; Verbeeck, H.; Bauters, M.; Beeckman, H.; Boeckx, P.; Huygens, D. Large-sized Rare Tree Species Contribute Disproportionately to Functional Diversity in Resource Acquisition in African Tropical Forest. Ecol. Evol. 2019, 9, 4349–4361. [Google Scholar] [CrossRef]
- Lutz, J.A.; Furniss, T.J.; Johnson, D.J.; Davies, S.J.; Allen, D.; Alonso, A.; Anderson-Teixeira, K.J.; Andrade, A.; Baltzer, J.; Becker, K.M.L.; et al. Global Importance of Large-diameter Trees. Glob. Ecol. Biogeogr. 2018, 27, 849–864. [Google Scholar] [CrossRef] [Green Version]
- ForestPlots.net; Blundo, C.; Carilla, J.; Grau, R.; Malizia, A.; Malizia, L.; Osinaga-Acosta, O.; Bird, M.; Bradford, M.; Catchpole, D.; et al. Taking the Pulse of Earth’s Tropical Forests Using Networks of Highly Distributed Plots. Biol. Conserv. 2021, 260, 108849. [Google Scholar] [CrossRef]
- Chave, J.; Coomes, D.; Jansen, S.; Lewis, S.L.; Swenson, N.G.; Zanne, A.E. Towards a Worldwide Wood Economics Spectrum. Ecol. Lett. 2009, 12, 351–366. [Google Scholar] [CrossRef]
- Zanne, A.E.; Lopez-Gonzalez, G.; Coomes, D.A.; Ilic, J.; Jansen, S.; Lewis, S.L.; Miller, R.B.; Swenson, N.G.; Wiemann, M.C.; Chave, J. Towards a Worldwide Wood Economics Spectrum 2009; 2047488 bytes; Dryad: Lewis County, WA, USA, 2009. [Google Scholar]
- Chave, J.; Réjou-Méchain, M.; Búrquez, A.; Chidumayo, E.; Colgan, M.S.; Delitti, W.B.C.; Duque, A.; Eid, T.; Fearnside, P.M.; Goodman, R.C.; et al. Improved Allometric Models to Estimate the Aboveground Biomass of Tropical Trees. Glob. Chang. Biol. 2014, 20, 3177–3190. [Google Scholar] [CrossRef] [PubMed]
- McElhinny, C.; Gibbons, P.; Brack, C.; Bauhus, J. Forest and Woodland Stand Structural Complexity: Its Definition and Measurement. For. Ecol. Manag. 2005, 218, 1–24. [Google Scholar] [CrossRef]
- Chao, A.; Gotelli, N.J.; Hsieh, T.C.; Sander, E.L.; Ma, K.H.; Colwell, R.K.; Ellison, A.M. Rarefaction and Extrapolation with Hill Numbers: A Framework for Sampling and Estimation in Species Diversity Studies. Ecol. Monogr. 2014, 84, 45–67. [Google Scholar] [CrossRef] [Green Version]
- Pinho, B.X.; Peres, C.A.; Leal, I.R.; Tabarelli, M. Critical role and collapse of tropical mega-trees: A key global resource. In Advances in Ecological Research; Elsevier: Amsterdam, The Netherlands, 2020; Volume 62, pp. 253–294. ISBN 978-0-12-821134-2. [Google Scholar]
- Li, S.; Su, J.; Lang, X.; Liu, W.; Ou, G. Positive Relationship between Species Richness and Aboveground Biomass across Forest Strata in a Primary Pinus Kesiya Forest. Sci. Rep. 2018, 8, 2227. [Google Scholar] [CrossRef]
- Mustin, K.; Carvalho, W.D.; Hilário, R.R.; Costa-Neto, S.V.; Silva, C.; Vasconcelos, I.M.; Castro, I.J.; Eilers, V.; Kauano, É.E.; Mendes-Junior, R.N.G.; et al. Biodiversity, Threats and Conservation Challenges in the Cerrado of Amapá, an Amazonian Savanna. Nat. Conserv. 2017, 22, 107–127. [Google Scholar] [CrossRef]
- De Lima, R.B.; Rutishauser, E.; da Silva, J.A.A.; Guedes, M.C.; Herault, B.; de Oliveira, C.P.; da Silva Aparício, P.; Sotta, E.D.; da Silva, D.A.S.; Ferreira, R.L.C. Accurate Estimation of Commercial Volume in Tropical Forests. For. Sci. 2021, 67, 14–21. [Google Scholar] [CrossRef]
- Pitman, N.C.A.; Terborgh, J.W.; Silman, M.R.; Núñez, P.; Neill, D.A.; Cerón, C.E.; Palacios, W.A.; Aulestia, M. Dominance and Distribution of Tree Species in Upper Amazonian Terra Firme Forests. Ecology 2001, 82, 2101–2117. [Google Scholar] [CrossRef]
- Pitman, N.C.A.; Silman, M.R.; Terborgh, J.W. Oligarchies in Amazonian Tree Communities: A Ten-Year Review. Ecography 2013, 36, 114–123. [Google Scholar] [CrossRef]
- Arellano, G.; Jørgensen, P.M.; Fuentes, A.F.; Loza, M.I.; Torrez, V.; Macía, M.J. Oligarchic Patterns in Tropical Forests: Role of the Spatial Extent, Environmental Heterogeneity and Diversity. J. Biogeogr. 2016, 43, 616–626. [Google Scholar] [CrossRef]
- Tuomisto, H.; Ruokolainen, K.; Yli-Halla, M. Dispersal, Environment, and Floristic Variation of Western Amazonian Forests. Science 2003, 299, 241–244. [Google Scholar] [CrossRef] [PubMed]
- Gorgens, E.B.; Motta, A.Z.; Assis, M.; Nunes, M.H.; Jackson, T.; Coomes, D.; Rosette, J.; Aragão, L.E.O.C.; Ometto, J.P. The Giant Trees of the Amazon Basin. Front. Ecol. Environ. 2019, 17, 373–374. [Google Scholar] [CrossRef] [Green Version]
- Gorgens, E.B.; Nunes, M.H.; Jackson, T.; Coomes, D.; Keller, M.; Reis, C.R.; Valbuena, R.; Rosette, J.; Almeida, D.R.A.; Gimenez, B.; et al. Resource Availability and Disturbance Shape Maximum Tree Height across the Amazon. Glob. Chang. Biol. 2021, 27, 177–189. [Google Scholar] [CrossRef] [PubMed]
- Aragão, L.E.O.C.; Poulter, B.; Barlow, J.B.; Anderson, L.O.; Malhi, Y.; Saatchi, S.; Phillips, O.L.; Gloor, E. Environmental Change and the Carbon Balance of Amazonian Forests: Environmental Change in Amazonia. Biol. Rev. 2014, 89, 913–931. [Google Scholar] [CrossRef] [PubMed]
- Marvin, D.C.; Asner, G.P.; Knapp, D.E.; Anderson, C.B.; Martin, R.E.; Sinca, F.; Tupayachi, R. Amazonian Landscapes and the Bias in Field Studies of Forest Structure and Biomass. Proc. Natl. Acad. Sci. USA 2014, 111, E5224–E5232. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fauset, S.; Johnson, M.O.; Gloor, M.; Baker, T.R.; Monteagudo, M.A.; Brienen, R.J.W.; Feldpausch, T.R.; Lopez-Gonzalez, G.; Malhi, Y.; ter Steege, H.; et al. Hyperdominance in Amazonian Forest Carbon Cycling. Nat. Commun. 2015, 6, 6857. [Google Scholar] [CrossRef] [PubMed]
- Duque, A.; Muller-Landau, H.; Valencia, R.; Cardenas, D.; Davies, S.; Oliveira, A.; Pérez, Á.; Romero-Saltos, H.; Vicentini, A. Insights into Regional Patterns of Amazonian Forest Structure, Diversity, and Dominance from Three Large Terra-Firme Forest Dynamics Plots. Biodivers. Conserv. 2017, 26, 669–686. [Google Scholar] [CrossRef]
- Draper, F.C.; Asner, G.P.; Honorio Coronado, E.N.; Baker, T.R.; García-Villacorta, R.; Pitman, N.C.A.; Fine, P.V.A.; Phillips, O.L.; Zárate Gómez, R.; Amasifuén Guerra, C.A.; et al. Dominant Tree Species Drive Beta Diversity Patterns in Western Amazonia. Ecology 2019, 100, e02636. [Google Scholar] [CrossRef] [Green Version]
- Crowther, T.W.; Glick, H.B.; Covey, K.R.; Bettigole, C.; Maynard, D.S.; Thomas, S.M.; Smith, J.R.; Hintler, G.; Duguid, M.C.; Amatulli, G.; et al. Mapping Tree Density at a Global Scale. Nature 2015, 525, 201–205. [Google Scholar] [CrossRef]
- Lines, E.R.; Zavala, M.A.; Purves, D.W.; Coomes, D.A. Predictable Changes in Aboveground Allometry of Trees along Gradients of Temperature, Aridity and Competition: Predictable Variation in Tree Aboveground Allometry. Glob. Ecol. Biogeogr. 2012, 21, 1017–1028. [Google Scholar] [CrossRef]
- Iida, Y.; Kohyama, T.S.; Kubo, T.; Kassim, A.R.; Poorter, L.; Sterck, F.; Potts, M.D. Tree Architecture and Life-History Strategies across 200 Co-Occurring Tropical Tree Species: Tree Architecture and Life-History Strategies across 200 Species. Funct. Ecol. 2011, 25, 1260–1268. [Google Scholar] [CrossRef]
- Rutishauser, E.; Hérault, B.; Petronelli, P.; Sist, P. Tree Height Reduction After Selective Logging in a Tropical Forest. Biotropica 2016, 48, 285–289. [Google Scholar] [CrossRef]
- Giardina, F.; Konings, A.G.; Kennedy, D.; Alemohammad, S.H.; Oliveira, R.S.; Uriarte, M.; Gentine, P. Tall Amazonian Forests Are Less Sensitive to Precipitation Variability. Nat. Geosci. 2018, 11, 405–409. [Google Scholar] [CrossRef]
- Jucker, T.; Hardwick, S.R.; Both, S.; Elias, D.M.O.; Ewers, R.M.; Milodowski, D.T.; Swinfield, T.; Coomes, D.A. Canopy Structure and Topography Jointly Constrain the Microclimate of Human-modified Tropical Landscapes. Glob. Chang. Biol. 2018, 24, 5243–5258. [Google Scholar] [CrossRef] [PubMed] [Green Version]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
de Lima, R.B.; Görgens, E.B.; Batista, A.P.B.; da Silva, D.A.S.; de Oliveira, C.P.; de Sousa, C.S.C. Diversity and Big Tree Patterns in the Brazilian Amazon. Diversity 2022, 14, 503. https://doi.org/10.3390/d14070503
de Lima RB, Görgens EB, Batista APB, da Silva DAS, de Oliveira CP, de Sousa CSC. Diversity and Big Tree Patterns in the Brazilian Amazon. Diversity. 2022; 14(7):503. https://doi.org/10.3390/d14070503
Chicago/Turabian Stylede Lima, Robson Borges, Eric Bastos Görgens, Anderson Pedro Bernardina Batista, Diego Armando Silva da Silva, Cinthia Pereira de Oliveira, and Carla Samara Campelo de Sousa. 2022. "Diversity and Big Tree Patterns in the Brazilian Amazon" Diversity 14, no. 7: 503. https://doi.org/10.3390/d14070503