
Morphological and Molecular Phylogenetic Analysis of a Lemanea Specimen (Batrachospermales, Rhodophyta) from China


Abstract
:1. Introduction
2. Materials and Methods
3. Results
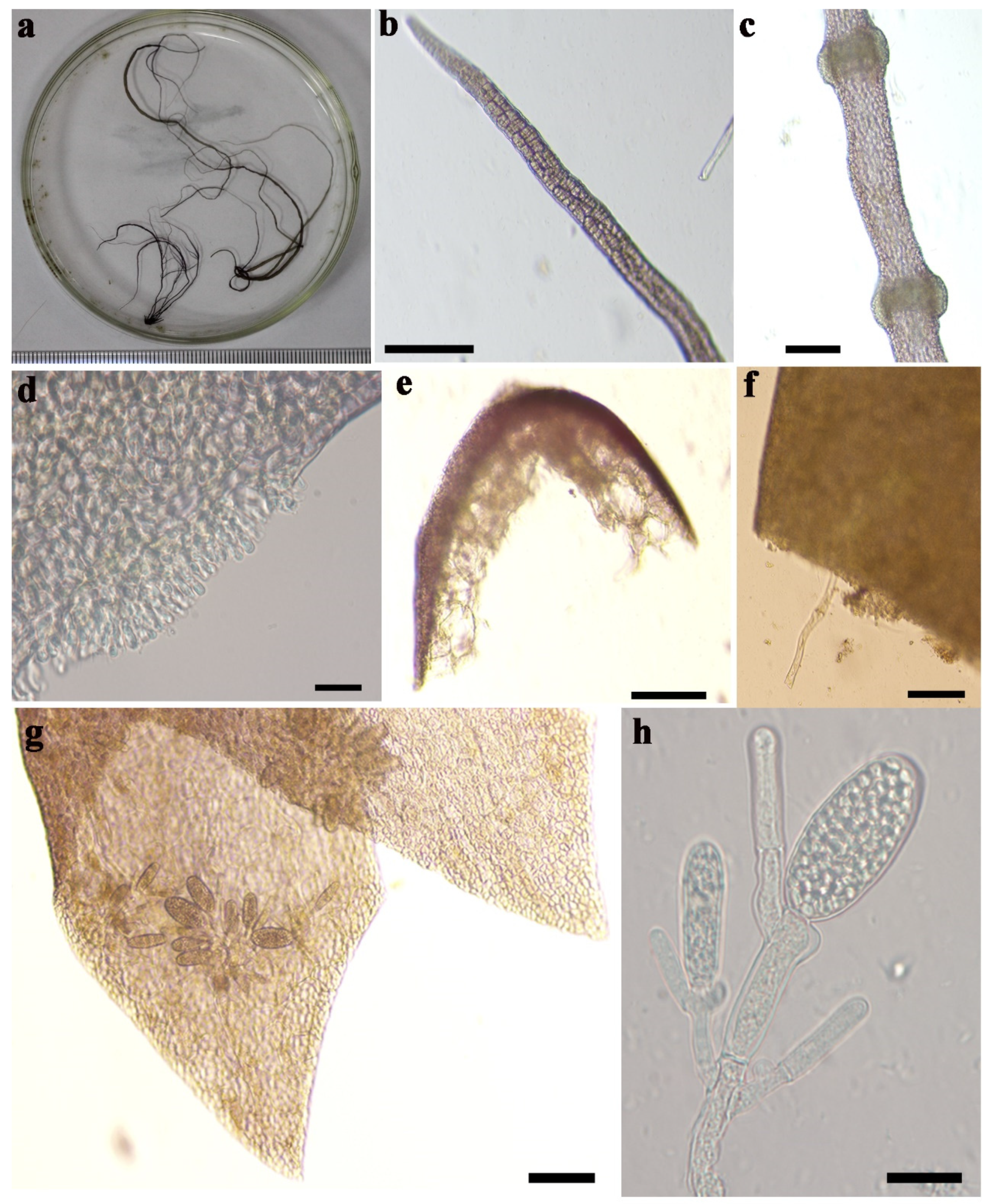
3.1. Morphological Characterization
3.2. Molecular Analysis
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Wehr, J.D.; Sheath, R.G.; Kociolek, J.P. Freshwater Algae of North America: Ecology and Classification; Elsevier: San Diego, CA, USA, 2015; ISBN 978-0-12-385876-4. [Google Scholar]
- Entwisle, T.J.; Vis, M.L.; Chiasson, W.B.; Necchi, O., Jr.; Sherwood, A.R. Systematics of the Batrachospermales (Rhodophyta)—A synthesis. J. Phycol. 2009, 45, 704–715. [Google Scholar] [CrossRef] [PubMed]
- Guiry, M.D.; Guiry, G.M. AlgaeBase. World-Wide Electronic Publication, National University of Ireland, Galway. Available online: http://www.algaebase.org/ (accessed on 18 March 2022).
- Vis, M.L.; Necchi, O., Jr. Subphylum Eurhodophytina, Class Florideophyceae, Subclass Nemaliophycidae, Order Batrachospermales. In Freshwater Red Alga: Phylogeny, Taxonomy and Biogeography; Vis, M.L., Necchi, O., Jr., Eds.; Springer Nature: Cham, Switzerland, 2021; pp. 129–332. ISBN 978-3-030-83969-7. [Google Scholar]
- Vis, M.L.; Sheath, R.G. Systematics of the freshwater red algal family Lemaneaceae in North America. Phycologia 1992, 31, 164–179. [Google Scholar] [CrossRef]
- Singh, M.R.; Gupta, A. Morphological characteristics of rare and endangered species of Lemanea (Rhodophyta) from the rivers of Manipur, north-east India. Botanique 2010, 14, 47–54. [Google Scholar]
- Bhosale, R.; Rout, J.; Chaugule, B. The ethnobotanical study of an edible fresh water red algae, Lemanea fluviatilis (L.) C. Ag. from Manipur, India. Ethnobot. Res. Appl. 2012, 10, 69–76. Available online: www.ethnobotanyjournal.org/vol10/i1547-3465-10-069 (accessed on 1 May 2022). [CrossRef] [Green Version]
- Singh, M.R.; Gupta, A. Biomass and productivity study of red algae from rivers of Manipur, North East India. In Ecoplanning, Biodiversity and Climate Change; Gupta, A., Ed.; Pointer Publishers: Jaipur, India, 2014; pp. 118–134. ISBN 978-81-7132-763-8. [Google Scholar]
- Snežana, B.; Simić, S. Morphological and ecological characteristics of rare and endangered species Lemanea fluviatilis (L.) C. Ag. (Lemaneaceae, Rhodophyta) on new localities in Serbia. Kragujev. J. Sci. 2007, 29, 97–106. [Google Scholar]
- Saber, A.A.; Cantonati, M.; Vis, M.L.; Anesi, A.; Guella, G. Multifaceted characterization of a Lemanea fluviatilis population (Batrachospermales, Rhodophyta) from a glacial stream in the south–eastern Alps. J. Czech Phycol. Soc. 2016, 16, 234–243. [Google Scholar] [CrossRef] [Green Version]
- Manolaki, P.; Guo, K.; Vieira, C.; Papastergiadou, E.; Riis, T. Hydromorphology as a controlling factor of macrophytes assemblage structure and functional traits in the semi-arid European Mediterranean streams. Sci. Total Environ. 2020, 703, 134658. [Google Scholar] [CrossRef]
- Ganesan, E.K.; West, J.A.; Zuccarello, G.C.; de Goer, S.L.; Rout, J. Lemanea manipurensis sp. nov. (Batrachospermales), a freshwater red algal species from North-East India. Algae 2015, 30, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Han, J.F.; Nan, F.R.; Feng, J.; Lv, J.P.; Liu, Q.; Kociolek, J.P.; Xie, S.L. Sheathia jinchengensis (Batrachospermales, Rhodophyta), a new freshwater red algal species described from North China. Phytotaxa 2018, 367, 63–70. [Google Scholar] [CrossRef]
- Necchi, O.; West, J.A.; Ganesan, E.K.; Yasmin, F.; Rai, S.K.; Rossignolo, N.L. Diversity of the genus Sheathia (Batrachospermales, Rhodophyta) in northeast India and east Nepal. Algae 2019, 34, 277–288. [Google Scholar] [CrossRef] [Green Version]
- Vis, M.L.; Saunders, G.W.; Sheath, R.G.; Dunse, K.; Entwisle, T.J. Phylogeny of the Batrachospermales (Rhodophyta) inferred from rbcL and 18S ribosomal DNA gene sequences. J. Phycol. 1998, 34, 341–350. [Google Scholar] [CrossRef]
- Li, Q.; Ji, L.; Xie, S.L. Phylogenetic analysis of Batrachospermales (Florideophyceae, Rhodophyta) based on chloroplast rbcL sequences. Acta Hydrob.Sinica 2010, 34, 20–28. [Google Scholar] [CrossRef]
- Thirb, H.H.; Benson-Evans, K. Cytological studies on Lemanea fluviatilis in the River Usk. Brit. Phycol. Bull. 1982, 17, 401–409. [Google Scholar] [CrossRef] [Green Version]
- Pertti, E.; Janina, K. Distribution and ecology of freshwater red algae (Rhodophyta) in some central Finnish rivers. Nord. J. Bot. 1996, 16, 107–117. [Google Scholar] [CrossRef]
- Eloranta, P.; Kwadrans, J. Freshwater red algae (Rhodophyta). Identification guide to European taxa, particularly to those in Finland. Norrlinia 2007, 15, 1–103. [Google Scholar]
- Kučera, P.; Grulich, V.; Fránková, M.; Bureš, P. Distribution of freshwater red algal family Lemaneaceae (Rhodophyta) in the Czech Republic: An update. J. Czech Phycol. Soc. 2008, 8, 125–128. [Google Scholar] [CrossRef] [Green Version]
- Temniskova, D.; Stoyneva, P.M.; Kirjakov, K.I. Red List of the Bulgarian algae. I. Macroalgae. Phytologia Balc. 2008, 14, 193–206. [Google Scholar]
- Simić, S.; Djordjević, N. Lemanea fucina Bory, 1808 (Lemaneaceae, Rhodophyta), a rare species with a variable morphology: First record in the Republic of Montenegro. Arch. Biol. Sci. 2011, 63, 511–515. [Google Scholar] [CrossRef]
- Ceschin, S.; Bisceglie, S.; Ricci, S. Contribution to the knowledge of red algae (Rhodophyta) of some rivers in central Italy. Cryptogam. Algol. 2012, 33, 61–67. [Google Scholar] [CrossRef]
- Khan, M. On edible Lemanea Bory de St Vincent—A freshwater red alga from India. Hydrobio. 1973, 43, 171–175. [Google Scholar] [CrossRef]
- Lipkovsky, E.; Barinova, S.; Nevo, E. Note: The first record of Lemanea fluviatilis L. Agardh (Rhodophyta, Nemaliales, Lemaneaceae), a fresh-water species in the upper Jordan River, Israel. Isr. J. Plant Sci. 2008, 56, 121–122. [Google Scholar] [CrossRef]
- Jao, C.C. Studies on the freshwater algae of China VIII. A preliminary account of the Chinese freshwater Rhodophyceae. Sinensia 1941, 12, 245–289. [Google Scholar]
- Xie, S.L.; Shi, Z.X.; Wang, R.N. Taxonomy of the Lemaneaceae (Batrachospermales, Rhodophyta) in China. J. Integr. Plant Biol. 2004, 46, 883–888. [Google Scholar]
- Wang, M.S. The community structure of the attached algae in Little Three Gorges of Jialing River during lower water periods and their uses in water quality evaluation. J. Southwest China Norm. Univ. Nat. Sci. 1994, 19, 305–310. [Google Scholar]
- Fang, L.Y. Research on Traditional Use of Aquatic Plants of Xishuangbanna Dai People; Xishuangbanna Tropical Botanical Garden, Chinese Academy of Sciences: Xishuangbanna, China, 2006. [Google Scholar]
- Li, X.Y.; Peng, M.C.; Dong, S.K.; Liu, S.L.; Li, J.P.; Yang, Z.F. Ecological risk assessment of hydropower dam construction on aquatic species in middle reaches of Lancang River, Southwest China based on Eshippo model. Chin. J. Appl. Ecol. 2013, 24, 517–526. [Google Scholar]
- Saunders, G.W. Gel purification of red algal genomic DNA: An inexpensive and rapid method for the isolation of polymerase chain reaction-friendly DNA. J. Phycol. 1993, 29, 251–254. [Google Scholar] [CrossRef]
- Vis, M.L.; Sheath, R.G. Biogeography of Batrachospermum gelatinosum (Batrachospermales, Rhodophyta) in North America based on molecular and morphological data. J. Phycol. 1997, 33, 520–526. [Google Scholar] [CrossRef]
- Freshwater, D.W.; Rueness, J. Phylogenetic relationships of some European Gelidium (Gelidiales, Rhodophyta) species based on rbcL nucleotide sequence analysis. Phycologia 1994, 33, 187–194. [Google Scholar] [CrossRef]
- Saunders, G.W.; Kraft, G.T. Small-subunit rRNA gene sequences from representatives of selected families of the Gigartinales and Rhodymeniales (Rhodophyta). 1. Evidence for the Plocamiales ord. nov. Can. J. Bot. 1994, 72, 1250–1263. [Google Scholar] [CrossRef]
- Thompson, J.D.; Gibson, T.J.; Plewniak, F.; Jeanmougin, F.; Higgins, D.G. The Clustal_X windows interface: Flexible strategies for multiple sequence alignment aided by quality analysis tools. Nucleic Acids Res. 1997, 25, 4876–4882. [Google Scholar] [CrossRef] [Green Version]
- Tamura, K.; Peterson, D.; Peterson, N.; Stecher, G.; Nei, M.; Kumar, S. MEGA5: Molecular evolutionary genetics analysis using maximum likelihood, evolutionary distance, and maximum parsimony methods. Mol. Biol. Evol. 2011, 28, 2731–2739. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Posada, D.; Buckley, T.R. Model selection and model averaging in phylogenetics: Advantages of Akaike information criterion and Bayesian approaches over likelihood ratio tests. Syst. Biol. 2004, 53, 793–808. [Google Scholar] [CrossRef] [PubMed]
- Felsenstein, J. Evolutionary trees from DNA sequences: A maximum likelihood approach. J. Mol. Evol. 1981, 17, 368–376. [Google Scholar] [CrossRef] [PubMed]
- Guindon, S.; Gascuel, O. A simple, fast, and accurate algorithm to estimate large phylogenies by maximum likelihood. Syst. Biol. 2003, 52, 696–704. [Google Scholar] [CrossRef] [Green Version]
- Ronquist, F.; Huelsenbeck, J.P. MrBayes 3: Bayesian phylogenetic inference under mixed models. Bioinformatics 2003, 19, 1572–1574. [Google Scholar] [CrossRef] [Green Version]
- Vis, M.L.; Harper, J.T.; Saunders, G.W. Large subunit rDNA and rbcL gene sequence data place Petrohua bernabei gen. et sp. nov. in the Batrachospermales (Rhodophyta), but do not provide further resolution among taxa in this order. Phycol. Res. 2007, 55, 103–112. [Google Scholar] [CrossRef]
- Chou, J.; Wang, W. Batrachospermum arcuatum Kylin (Batrachospermales, Rhodophyta), a freshwater red alga newly recorded in Taiwan. Taiwania Taipei 2006, 51, 58–63. [Google Scholar] [CrossRef]
- Chiasson, W.B.; Salomaki, E.D.; Vis, M.L. New collections of freshwater red algae (Batrachospermales, Rhodophyta) from historically important areas in France. Cryptogam. Algol. 2014, 35, 303–316. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| GLYZC | L. manipurensis | L. sinica | L. crassa | L. ramosa | L. simplex | |
|---|---|---|---|---|---|---|
| Thallus height/cm | 10–13 | 10 | 9–16 | 9–13 | 1–3 | 2 |
| Branch | No branch at the base, scarcely branched at the lower part | Sparse, irregular branching in the lowermost region of the erect shoots | Opposite, alternate or forked branches | No branch | Densely branched, especially in the upper part | No branch |
| Stalk | No obvious stalk | No obvious stalk | Slender stalk | Slender stalk | No obvious stalk | No obvious stalk, |
| Node | Obviously expanded | Expanded | Obviously expanded | Obviously expanded | Obviously expanded or not | irregularly corrugated constriction |
| Carposporophyte | Ellipsoidal, single, 41–52 µm in length, 12–25 µm in diameter | Ellipsoidal, single, 80–120 µm in length, 30–50 µm in diameter | Columnar obovate, single, 75–95 µm in length, 30–50 µm in diameter | Columnar obovate, oval or dumbbell shape; single; 90–120 µm in length; 50–80 µm in diameter | Oval, single, 80–100 µm in length, 25–60 µm in diameter | Columnar obovate, single, 110–140 µm in length, 50–70 µm in diameter |
| Spermatangium | Continuous wide rings on the nodes | Isolated irregular patches, sometimes completely covering the axis | Continuous wide rings on the nodes on the upper part; incomplete rings on the lower part | In isolated patches, usually 5–6 obvious protrusions | In isolated patches, usually 6–8, or incomplete rings | In isolated patches, irregular, usually 2–4 formed rings |
| Reference | This study | [12] | [27] | [27] | [27] | [27] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nan, F.; Zhao, Y.; Feng, J.; Lv, J.; Liu, Q.; Liu, X.; Xie, S. Morphological and Molecular Phylogenetic Analysis of a Lemanea Specimen (Batrachospermales, Rhodophyta) from China. Diversity 2022, 14, 479. https://doi.org/10.3390/d14060479
Nan F, Zhao Y, Feng J, Lv J, Liu Q, Liu X, Xie S. Morphological and Molecular Phylogenetic Analysis of a Lemanea Specimen (Batrachospermales, Rhodophyta) from China. Diversity. 2022; 14(6):479. https://doi.org/10.3390/d14060479
Chicago/Turabian StyleNan, Fangru, Yiding Zhao, Jia Feng, Junping Lv, Qi Liu, Xudong Liu, and Shulian Xie. 2022. "Morphological and Molecular Phylogenetic Analysis of a Lemanea Specimen (Batrachospermales, Rhodophyta) from China" Diversity 14, no. 6: 479. https://doi.org/10.3390/d14060479