
Variation in Plant Diversity along a Watershed in the Semi-Arid Lands of North Africa



Abstract
:1. Introduction
2. Materials and Methods
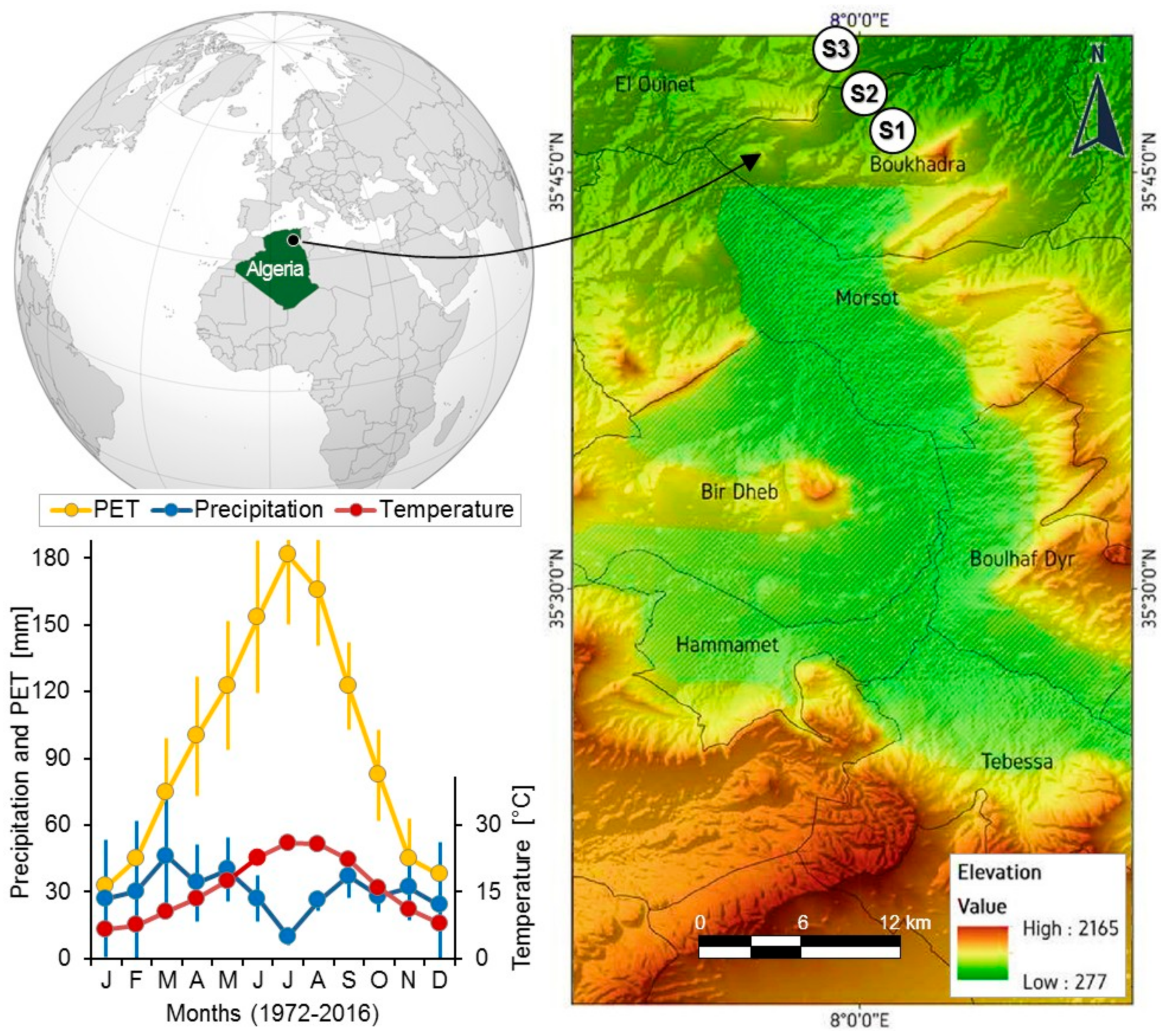
2.1. Study Area and Sampling Sites
2.2. Sampling Plants
2.3. Evaluation of Plant Cover, Occurrence, and Life forms
2.4. Alpha and Beta Diversity, Rarefaction, and Interpolation of Species Richness
2.5. Taxonomic Diversity
2.6. Data Management and Statistical Analysis
3. Results
3.1. Soil Characteristics of Vegetation Types
3.2. Floristic Composition
3.3. Biological Spectrum
3.4. Taxonomic Structures
3.5. Species Diversity/Alpha Diversity
3.6. Intra-Relationships between Species Richness Estimates
3.7. Similarity Analysis between Phytoecological Groups
3.8. Similarity Analysis between Sampling Sites at Different Watershed Scales
3.9. Spatial Relationships between Soil and Plant Functional Traits
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Oertli, B. The use of dragonflies in the assessment and monitoring of aquatic habitats. In Dragonflies and Damselflies: Model Organisms for Ecological and Evolutionary Research; Alex Córdoba-Aguilar, A., Ed.; Oxford University Press: Oxford, UK, 2008; pp. 79–95. [Google Scholar] [CrossRef]
- Simaika, J.P.; Samways, M.J. Using dragonflies to monitor and prioritize lotic systems: A South African perspective. Org. Divers. Evol. 2012, 12, 251–259. [Google Scholar] [CrossRef]
- El-Hawagry, M.S.; Gilbert, F. Catalogue of the Syrphidae of Egypt (Diptera). Zootaxa 2019, 4577, 201–248. [Google Scholar] [CrossRef] [PubMed]
- Wei, X.; Li, Q.; Zhang, M.; Giles-Hansen, K.; Liu, W.; Fan, H.; Wang, Y.; Zhou, G.; Piao, S.; Liu, S. Vegetation cover—Another dominant factor in determining global water resources in forested regions. Glob. Chang. Biol. 2018, 24, 786–795. [Google Scholar] [CrossRef]
- Zajac, E.; Zarzycki, J. The effect of thermal activity of colliery waste heap on vegetation development. Rocz. Ochr. Srodowiska 2013, 15, 1862–1880. [Google Scholar] [CrossRef]
- Belgacem, A.O.; Tarhouni, M.; Louhaichi, M. Effect of protection on plant community dynamics in the Mediterranean arid zone of southern Tunisia: A case study from Bou Hedma national park. Land Degrad. Dev. 2013, 24, 57–62. [Google Scholar] [CrossRef]
- Riis, T.; Sand-Jensen, K.; Vestergaard, O. Plant communities in lowland Danish streams: Species composition and environmental factors. Aquat. Bot. 2000, 66, 255–272. [Google Scholar] [CrossRef]
- Vannote, R.L.; Minshall, G.W.; Cummins, K.W.; Sedell, J.R.; Cushing, C.E. The river continuum concept. Can. J. Fish. Aquat. Sci. 1980, 37, 130–137. [Google Scholar] [CrossRef]
- Amirouche, R.; Misset, M.T. Flore spontanée d’Algérie: Différenciation écogéographique des espèces et polyploïdie. Cah. Agric. 2009, 18, 474–480. [Google Scholar] [CrossRef]
- Cayuela, L.; Rey Benayas, J.M.; Maestre, F.T.; Escudero, A. Early environments drive diversity and floristic composition in Mediterranean old fields: Insights from a long-term experiment. Acta Oecol. 2008, 34, 311–321. [Google Scholar] [CrossRef]
- Souahi, H.; Gharbi, A.; Gassarellil, Z. Growth and physiological responses of cereals species under lead stress. Int. J. Biosci. 2017, 11, 266–273. [Google Scholar] [CrossRef]
- Souahi, H.; Chebout, A.; Akrout, K.; Massaoud, N.; Gacem, R. Physiological responses to lead exposure in wheat, barley and oat. Environ. Chall. 2021, 4, 100079. [Google Scholar] [CrossRef]
- Souahi, H. Impact of lead on the amount of chlorophyll and carotenoids in the leaves of Triticum durum and T. aestivum, Hordeum vulgare and Avena sativa. Biosyst. Divers. 2021, 29, 207–210. [Google Scholar] [CrossRef]
- Ellis, J.E. Climate variability and complex ecosystem dynamics: Implications for pastoral development. In Living with Uncertainty: New Direction in Pastoral Development in Africa; Scoones, I., Ed.; Intermediate Technology Publications: London, UK, 1995; pp. 37–46. [Google Scholar]
- Fatmi, H.; Mâalem, S.; Harsa, B.; Dekak, A.; Chenchouni, H. Pollen morphological variability correlates with a large-scale gradient of aridity. Web Ecol. 2020, 20, 19–32. [Google Scholar] [CrossRef]
- Djebaili, S. Recherches Phytosociologiques et Phytoécologiques sur la Végétation des Hautes Plaines Steppiques et de l’Atlas Saharien. Ph.D. Thesis, University Montpellier, Montpellier, France, 1978. [Google Scholar]
- Quézel, P.; Santa, S. Nouvelle Fore de l’Algérie et des Régions Désertiques Méridionales; CNRS: Paris, France, 1963; Volume II. [Google Scholar]
- Ozenda, P. Flore du Sahara, 2nd ed.; CNRS: Paris, France, 1983. [Google Scholar]
- Bonneau, M.; Souchier, B. Pédologie 2. In Constituants et Propriétés du Sol; Masson: Paris, France, 1994. [Google Scholar]
- Mathieu, C.; Pieltain, F.; Jeanroy, E. Analyse Chimique de Sol: Méthodes Choisies; Tec & Doc: Paris, France, 2003. [Google Scholar]
- Bonham, C.D. Measurements for Terrestrial Vegetation; John Wiley & Sons: Hoboken, NJ, USA, 2013. [Google Scholar]
- Gillet, F. La phytosociologie synusiale intégrée. In Guide Méthodologique Document 1, 4th ed.; Documents de Laboratoire d’Ecologie Végétale; Revue et Corrigée; Université de Neuchâtel—Institut Botanique: Neuchâtel, Switzerland, 2000; 68p. [Google Scholar]
- Elzinga, C.L.; Salzer, D.W.; Willoughby, J.W.; Gibbs, J.P. Monitoring Plant and Animal Populations: A Handbook for Field Biologists; John Wiley & Sons: Hoboken, NJ, USA, 2009. [Google Scholar]
- Chao, A.; Gotelli, N.J.; Hsieh, T.C.; Sander, E.L.; Ma, K.H.; Colwell, R.K.; Ellison, A.M. Rarefaction and extrapolation with Hill numbers: A framework for sampling and estimation in species diversity studies. Ecol. Monogr. 2014, 84, 45–67. [Google Scholar] [CrossRef] [Green Version]
- Pielou, E.C. An introduction to mathematical ecology. Wiley Interscience. John Wiley & Sons, New York 1969. VIII + 286 S., 32 Abb., Preis 140 s. Biom. Z. 1971, 13, 219–220. [Google Scholar] [CrossRef]
- Colwell, R.K. EstimateS: Statistical Estimation of Species Richness and Shared Species from Samples. Version 9 and Earlier. User’s Guide and Application. 2013. Available online: http://purl.oclc.org/estimates (accessed on 22 December 2021).
- Bouallala, M.; Neffar, S.; Chenchouni, H. Vegetation traits are accurate indicators of how do plants beat the heat in drylands: Diversity and functional traits of vegetation associated with water towers in the Sahara Desert. Ecol. Indic. 2020, 114, 106364. [Google Scholar] [CrossRef]
- Passy, S.I.; Legendre, P. Power law relationships among hierarchical taxonomic categories in algae reveal a new paradox of the plankton. Glob. Ecol. Biogeogr. 2006, 15, 528–535. [Google Scholar] [CrossRef]
- Enquist, B.J.; Haskell, J.P.; Tiffney, B.H. General patterns of taxonomic and biomass partitioning in extant and fossil plant communities. Nature 2002, 419, 610–613. [Google Scholar] [CrossRef]
- Wang, Z.Q. Geostatistics and Its Application in Ecology; Science Press: Beijing, China, 2001; pp. 162–192. [Google Scholar]
- Zhang, J.; Song, C.; Wenyan, Y. Tillage effects on soil carbon fractions in the Sanjiang Plain, Northeast China. Soil Tillage Res. 2007, 93, 102–108. [Google Scholar] [CrossRef]
- Burghardt, W. Soils in urban and industrial environments. Z. Pflanz. Bodenkd. 1994, 157, 205–214. [Google Scholar] [CrossRef]
- Ray, J.G.; George, J. Phytosociology of roadside communities to identify ecological potentials of tolerant species. J. Ecol. Nat. Environ. 2009, 1, 184–190. [Google Scholar] [CrossRef]
- Pulchérie, M.N.; Ndemba Etim, S.I.N.G.; Djumyom Wafo, G.V.; Djocgoue, P.F.; Kengne Noumsi, I.M.; Ngnien, A.W. Floristic surveys of hydrocarbon-polluted sites in some Cameroonian cities (Central Africa). Int. J. Phytoremediation 2018, 20, 191–204. [Google Scholar] [CrossRef]
- Al Gifri, A.N.; Hussein, M.A. Plant communities along the road from Aden to Sheikh Salem, Abyen, Yemen. Feddes Rep. 1993, 104, 267–270. [Google Scholar] [CrossRef]
- Abdel Khalik, K.; Al-Gohary, I.; Al-Sodany, Y. Floristic composition and vegetation: Environmental relationships of Wadi Fatimah, Mecca, Saudi Arabia. Arid. Land Res. Manag. 2017, 31, 316–334. [Google Scholar] [CrossRef]
- Zine, H.; Hakkou, R.; Elgadi, S.; Diarra, A.; El Adnani, M.; Ouhammou, A. Floristic and ecological monitoring on a store-and-release cover in arid and semi-arid environment of Kettara mine, Morocco. Acta Ecol. Sin. 2021, 41, 432–441. [Google Scholar] [CrossRef]
- Durand, J.H. Irrigable Soils; Cultural and Technical Cooperation Agency: Paris, France, 1983. (In French) [Google Scholar]
- Koull, N.; Chehma, A. Soil characteristics and plant distribution in saline wetlands of Oued Righ, northeastern Algeria. J. Arid. Land 2016, 8, 948–959. [Google Scholar] [CrossRef]
- Barrett, G. Vegetation communities on the shores of a salt lake in semi-arid Western Australia. J. Arid. Environ. 2006, 67, 77–89. [Google Scholar] [CrossRef]
- Boudjabi, S.; Chenchouni, H. Soil fertility indicators and soil stoichiometry in semi-arid steppe rangelands. Catena 2022, 210, 105910. [Google Scholar] [CrossRef]
- Bradai, L.; Bouallala, M.H.; Bouziane, N.F.; Zaoui, S.; Neffar, S.; Chenchouni, H. An appraisal of eremophyte diversity and plant traits in a rocky desert of the Sahara. Folia Geobot. 2015, 50, 239–252. [Google Scholar] [CrossRef]
- Azizi, M.; Chenchouni, H.; Belarouci, M.E.H.; Bradai, L.; Bouallala, M. Diversity of psammophyte communities on sand dunes of the Sahara Desert. J. King Saud Univ. Sci. 2021, 33, 101656. [Google Scholar] [CrossRef]
- Gutterman, Y. Strategies of seed dispersal and germination in plants inhabiting deserts. Bot. Rev. 1994, 60, 373–425. [Google Scholar] [CrossRef]
- Sádlo, J.; Chytrý, M.; Pergl, J.; Pyšek, P. Plant dispersal strategies: A new classification based on the multiple dispersal modes of individual species. Preslia 2018, 90, 1–22. [Google Scholar] [CrossRef] [Green Version]
- Macheroum, A.; Kadik, L.; Neffar, S.; Chenchouni, H. Environmental drivers of taxonomic and phylogenetic diversity patterns of plant communities in semi-arid steppe rangelands of North Africa. Ecol. Indic. 2021, 132, 108279. [Google Scholar] [CrossRef]
- Raunkiaer, C. The Life Forms of Plants and Statistical Plant Geography; Clarendon Press: Oxford, UK, 1934. [Google Scholar]
- Gamoun, M.; Belgacem, A.O.; Hanchi, B.; Neffati, M.; Gillet, F. Impact of grazing on the floristic diversity of arid rangelands in South Tunisia. Rev. Ecol. Terre Vie 2012, 67, 271–282. [Google Scholar]
- Hakkou, R.; Benzaazoua, M.; Bussiere, B. Laboratory evaluation of the use of alkaline phosphate wastes for the control of acidic mine drainage. Mine Water Environ. 2009, 28, 206–218. [Google Scholar] [CrossRef]
- Ramade, F. Eléments d’Ecologie: Ecologie Fondamentale, 4th ed.; Dunod: Paris, France, 2009. [Google Scholar]
- García-Fayos, P.; Bochet, E.; Cerdà, A. Seed removal susceptibility through soil erosion shapes vegetation composition. Plant Soil 2010, 334, 289–297. [Google Scholar] [CrossRef] [Green Version]
- Souahi, H.; Meksem Amara, L.; Meksem, N.; Maalem, S.; Djebar, M.R. Sulfonylureas effect on soil chemical properties and yield crop in Semi-arid region of Algeria. J. Biodivers. Environ. Sci. 2015, 7, 157–167. [Google Scholar]
- Pennings, S.C.; Callaway, R.M. Salt marsh plant zonation: The relative importance of competition and physical factors. Ecology 1992, 73, 681–690. [Google Scholar] [CrossRef]
- Chenchouni, H. Edaphic factors controlling the distribution of inland halophytes in an ephemeral salt lake “Sabkha ecosystem” at North African semi-arid lands. Sci. Total Environ. 2017, 575, 660–671. [Google Scholar] [CrossRef]
- Bezzalla, A.; Houhamdi, M.; Chenchouni, H. Vegetation Analysis of Chott Tinsilt and Sebkhet Ezzemoul (Two Ramsar Sites in Algeria) in Relation to Soil Proprieties. In Exploring the Nexus of Geoecology, Geography, Geoarcheology and Geotourism; Springer: Cham, Switzerland, 2019; pp. 39–42. [Google Scholar] [CrossRef]
- Bossé, B.; Bussiere, B.; Hakkou, R.; Maqsoud, A.; Benzaazoua, M. Assessment of phosphate limestone wastes as a component of a store-and-release cover in a semiarid climate. Mine Water Environ. 2013, 32, 152–167. [Google Scholar] [CrossRef]
- Grigore, M.N. From Molecules to Ecosystems towards Biosaline Agriculture. In Handbook of Halophytes; Springer: Cham, Switzerland, 2020. [Google Scholar] [CrossRef]
- Souahi, H.; Meksem Amara, L.; Nedjoud, G.; Djebar, M.R. Physiology and biochemistry effects of herbicides sekator and zoom on two varieties of wheat (Waha and HD) in semi-arid region. Annu. Res. Rev. Biol. 2015, 449–459. [Google Scholar] [CrossRef]
- Souahi, H.; Meksem Amara, L.; Djebar, M.R. Effects of sulfonylurea herbicides on protein content and antioxidants activity in wheat in semi-arid region. Int. J. Adv. Eng. Manag. Sci. 2016, 2, 239625. [Google Scholar]
- Bunn, S.E.; Arthington, A.H. Basic principles and ecological consequences of altered flow regimes for aquatic biodiversity. Environ. Manag. 2002, 30, 492–507. [Google Scholar] [CrossRef] [Green Version]
- Fausch, K.D.; Torgersen, C.E.; Baxter, C.V.; Li, H.W. Landscapes to riverscapes: Bridging the gap between research and conservation of stream fishes: A continuous view of the river is needed to understand how processes interacting among scales set the context for stream fishes and their habitat. BioScience 2002, 52, 483–498. [Google Scholar] [CrossRef] [Green Version]
- Lytle, D.A.; Poff, N.L. Adaptation to natural flow regimes. Trends Ecol. Evol. 2004, 19, 94–100. [Google Scholar] [CrossRef] [PubMed]
- McMullen, L.E.; Lytle, D.A. Quantifying invertebrate resistance to floods: A global-scale meta-analysis. Ecol. Appl. 2012, 22, 2164–2175. [Google Scholar] [CrossRef]
- Beismann, H.; Wilhelmi, H.; Baillères, H.; Spatz, H.C.; Bogenrieder, A.; Speck, T. Brittleness of twig bases in the genus Salix: Fracture mechanics and ecological relevance. J. Exp. Bot. 2000, 51, 617–633. [Google Scholar] [CrossRef]
- Bhandari, J.; Zhang, Y. Effect of altitude and soil properties on biomass and plant richness in the grasslands of Tibet, China, and Manang District, Nepal. Ecosphere 2019, 10, e02915. [Google Scholar] [CrossRef]
- Giweta, M. Role of litter production and its decomposition, and factors affecting the processes in a tropical forest ecosystem: A review. J. Ecol. Environ. 2020, 44, 11. [Google Scholar] [CrossRef]
- Salahab, Y.M.S.; Scholes, M.C. Effect of temperature and litter quality on decomposition rate of Pinus patula needle litter. Procedia Environ. Sci. 2011, 6, 180–193. [Google Scholar] [CrossRef] [Green Version]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Characteristics | Statistics | Downstream | Midstream | Upstream | Overall |
|---|---|---|---|---|---|
| Soil Properties | |||||
| Clay (%) | Mean ± SD Min–Max CV; Med | 1.3 ± 0.75 0.26–2.6 57.65; 1.22 | 1.07 ± 1.26 0.08–3.78 118.55; 0.36 | 3.85 ± 4.16 0.38–13.64 108.01; 2.61 | 2.07 ± 2.77 0.08–13.64 133.32; 1.22 |
| Silt (%) | Mean ± SD Min–Max CV; Med | 1.8 ± 1.34 0.18–3.69 74.1; 1.76 | 1.99 ± 2.85 0.08–9.12 143.37; 1.48 | 5.64 ± 9.79 0.14–31.24 173.6; 2.4 | 3.14 ± 5.98 0.08–31.24 190.26; 1.76 |
| Sand (%) | Mean ± SD Min–Max CV; Med | 29.08 ± 8.32 13.7–41.3 28.61; 29.35 | 24.35 ± 19.04 11.15–73.65 78.19; 17.44 | 22.85 ± 7.33 12.51–32.72 32.07; 23.42 | 25.42 ± 12.52 11.15–73.65 49.23; 23.42 |
| Gravel (%) | Mean ± SD Min–Max CV; Med | 67.81 ± 10.11 52.41–85.86 14.91; 67.2 | 72.6 ± 20.89 20.08–85.6 28.78; 79.7 | 67.66 ± 14.76 35.04–84.86 21.81; 72 | 69.36 ± 15.43 20.08–85.86 22.25; 72.84 |
| pH | Mean ± SD Min–Max CV; Med | 7.51 ± 0.28 7.23–7.96 3.75; 7.43 | 7.24 ± 0.14 7.09–7.54 1.99; 7.21 | 7.21 ± 0.27 6.89–7.75 3.75; 7.16 | 7.32 ± 0.27 6.89–7.96 3.68; 7.24 |
| Electrical conductivity (µS/cm) | Mean ± SD Min–Max CV; Med | 718.2 ± 163.9 449–890 22.82; 798 | 2245.1 ± 423.9 1535–2850 18.88; 2150 | 1211 ± 244.3 953–1787 20.17; 1150 | 1391.4 ± 708.8 449–2850 50.94; 1150 |
| Organic matter (%) | Mean ± SD Min–Max CV; Med | 1.45 ± 0.24 1.14–1.79 16.53; 1.42 | 1.12 ± 0.19 0.76–1.32 16.99; 1.17 | 0.28 ± 0.06 0.2–0.37 20.03; 0.28 | 0.95 ± 0.53 0.2–1.79 55.62; 1.14 |
| Organic carbon (%) | Mean ± SD Min–Max CV; Med | 0.84 ± 0.14 0.66–1.04 16.53; 0.83 | 0.65 ± 0.11 0.44–0.77 16.99; 0.68 | 0.16 ± 0.03 0.12–0.22 20.03; 0.16 | 0.55 ± 0.31 0.12–1.04 55.62; 0.66 |
| Soil-surface cover (%) | |||||
| Total vegetation cover | Mean ± SD Min–Max CV; Med | 53.78 ± 8.04 43–67 14.96; 52 | 25.44 ± 4.98 19–34 19.56; 25 | 67.56 ± 13.19 45–82 19.53; 71 | 48.93 ± 20.01 19–82 40.89; 51 |
| Plant litter | Mean ± SD Min–Max CV; Med | 14.78 ± 6.1 2–24 41.27; 15 | 11.22 ± 6.4 3–24 57.02; 10 | 17.56 ± 3.32 12–24 18.92; 18 | 14.52 ± 5.87 2–24 40.41; 15 |
| Coarse materials | Mean ± SD Min–Max CV; Med | 21.33 ± 6.3 11–28 29.55; 23 | 27.22 ± 14 9–49 51.42; 26 | 16.89 ± 4.08 9–21 24.13; 19 | 21.81 ± 9.81 9–49 44.97; 20 |
| Bare ground | Mean ± SD Min–Max CV; Med | 25.56 ± 5.36 20–36 20.99; 24 | 42.44 ± 9.57 25–57 22.54; 43 | 18.22 ± 9.12 5–34 50.05; 17 | 28.74 ± 13.02 5–57 45.29; 27 |
| Family | Species | RLF | Upstream | Midstream | Downstream | Total | ||||
|---|---|---|---|---|---|---|---|---|---|---|
| N | Occ | N | Occ | N | Occ | N | Occ | |||
| Amaranthaceae | Beta vulgaris Thell. | Ther | 129 | 100 | 157 | 88.9 | - | - | 286 | 63 |
| [4.76%] | Salsola vermiculata L. | Cham | 23 | 66.7 | 26 | 55.6 | 36 | 88.9 | 85 | 70.4 |
| Apiaceae * | Scandix pecten-veneris L. | Ther | 31 | 66.7 | 9 | 55.6 | 20 | 44.4 | 60 | 55.6 |
| Asteraceae | Anacyclus radiatus Lois. | Ther | 10 | 33.3 | - | - | - | - | 10 | 11.1 |
| [38.10%] | Atractylis delicatula L. | Hemi | 23 | 88.9 | 31 | 66.7 | 24 | 88.9 | 78 | 81.5 |
| Atractylis humilis L. | Hemi | 121 | 66.7 | - | - | - | - | 121 | 22.2 | |
| Bellis sylvestris Cirillo | Hemi | 20 | 77.8 | - | - | 18 | 66.7 | 38 | 48.1 | |
| Calendula arvensis L. | Ther | 55 | 100 | 74 | 77.8 | 3 | 22.2 | 132 | 66.7 | |
| Carduncellus pinnatus Desf. | Hemi | 11 | 44.4 | 70 | 66.7 | 234 | 100 | 315 | 70.4 | |
| Carduus pycnocephalus L. | Ther | 2 | 11.1 | - | - | - | - | 2 | 3.7 | |
| Carthamus lanatus L. | Ther | 3 | 11.1 | 1 | 11.1 | 31 | 66.7 | 35 | 29.6 | |
| Echinops spinosus L. | Cham | 1 | 11.1 | - | - | - | - | 1 | 3.7 | |
| Hedypnois cretica L. | Ther | 8 | 33.3 | 47 | 33.3 | 8 | 55.6 | 63 | 40.7 | |
| Hertia cheirifolia L. | Hemi | 1 | 11.1 | 1 | 11.1 | - | - | 2 | 7.41 | |
| Matthiola lunata DC. | Ther | 1 | 11.1 | - | - | - | - | 1 | 3.7 | |
| Onopordum acanthium L. | Hemi | 23 | 33.3 | - | - | - | - | 23 | 11.1 | |
| Reichardia picroides L. | Ther | 25 | 55.6 | 18 | 44.4 | 9 | 44.4 | 52 | 48.1 | |
| Scolymus hispanicus L. | Hemi | - | - | - | - | 41 | 100 | 41 | 33.3 | |
| Xanthium spinosum L. | Ther | 23 | 44.4 | 76 | 100 | - | - | 99 | 48.1 | |
| Boraginaceae * | Echium italicum L. | Ther | 3 | 22.2 | - | - | - | - | 3 | 7.41 |
| Brassicaceae | Eruca vesicaria L. Car. | Ther | 44 | 88.9 | 9 | 22.2 | - | - | 53 | 37 |
| [7.14%] | Moricandia arvensis DC | Hemi | 62 | 88.9 | 11 | 33.3 | - | - | 73 | 40.7 |
| Sisymurum irio L. | Ther | 23 | 66.7 | 13 | 33.3 | - | - | 36 | 33.3 | |
| Caryophyllaceae * | Paronychia argentea Lam. | Hemi | 3 | 22.2 | - | - | - | - | 3 | 7.41 |
| Chenopodiaceae | Arthrocnemum indicum Willd. | Hemi | 6 | 11.1 | - | - | - | - | 6 | 3.7 |
| [4.76%] | Atriplex halimus L. | Cham | 97 | 100 | - | - | 280 | 100 | 377 | 66.7 |
| Cupressaceae * | Juniperus oxycedrus L. | Phan | 0 | -- | 1 | 11.1 | - | - | 1 | 3.7 |
| Euphorbiaceae * | Euphorbia helioscapia L. | Ther | 4 | 11.1 | - | - | - | - | 4 | 3.7 |
| Fabaceae * | Retama raetam L. | Phan | 65 | 44.4 | - | - | 104 | 66.7 | 169 | 37 |
| Frankeniaceae * | Frankenia Thymifolia Desf. | Cham | 7 | 44.4 | - | - | - | - | 7 | 14.8 |
| Geraniaceae * | Erodium cicutarium L. | Ther | 33 | 33.3 | - | - | - | - | 33 | 11.1 |
| Lamiaceae * | Marrubium vulgare L. | Hemi | 12 | 66.7 | - | - | - | - | 12 | 22.2 |
| Malvaceae * | Malva sylvestris L. | Hemi | 41 | 100 | 3 | 11.1 | - | - | 44 | 37 |
| Plantaginaceae * | Plantago lenceolata L. | Hemi | 3 | 22.2 | 4 | 11.1 | - | - | 7 | 11.1 |
| Poaceae | Ampelodesmos mauritanicus Poir. | Hemi | 40 | 66.7 | - | - | - | - | 40 | 22.2 |
| [14.29%] | Arundo donax L. | Geo | - | - | 74 | 11.1 | - | - | 74 | 3.7 |
| Bromus rubens L. | Ther | 39 | 22.2 | - | - | - | - | 39 | 7.41 | |
| Hordeum maritimim Huds | Ther | 75 | 55.6 | 26 | 33.3 | 62 | 66.7 | 163 | 51.9 | |
| Lolium perenne L. | Hemi | 200 | 100 | 11 | 44.4 | 45 | 77.8 | 256 | 74.1 | |
| Stipa tenacissima L. | Hemi | 29 | 66.7 | 23 | 77.8 | - | - | 52 | 48.1 | |
| Rhamnaceae * | Ziziphus lotus L. | Cham | 3 | 22.2 | - | - | - | - | 3 | 7.41 |
| Tamaricaceae * | Tamarix balansea J.Gay | Phan | 8 | 22.2 | - | - | 35 | 33.3 | 43 | 18.5 |
| Families = 18 | Genera = 41, Species = 42 | N = | 1307 | 685 | 950 | 2942 | ||||
| Variables | Genus/Species (G/S) Ratio | Family/Species (F/S) Ratio | |||||||
|---|---|---|---|---|---|---|---|---|---|
| Upstream | Midstream | Downstream | Total | Upstream | Midstream | Downstream | Total | ||
| Descriptive statistics | |||||||||
| Minimum | 1 | 1 | 1 | 1 | 0.41 | 0.40 | 0.43 | 0.4 | |
| Maximum | 1.06 | 1.14 | 1 | 1.14 | 0.70 | 0.75 | 0.67 | 0.75 | |
| Median | 1 | 1 | 1 | 1 | 0.50 | 0.50 | 0.50 | 0.5 | |
| Mean | 1.02 | 1.02 | 1 | 1.01 | 0.52 | 0.51 | 0.52 | 0.51 | |
| Standard deviation | 0.03 | 0.05 | 0 | 0.03 | 0.08 | 0.11 | 0.08 | 0.09 | |
| Coefficient of variation (CV) | 0.02 | 0.04 | 0 | 0.03 | 0.15 | 0.20 | 0.14 | 0.17 | |
| Pearson correlation tests | |||||||||
| Plant litter | r | 0.082 | −0.479 | −0.067 | −0.231 | 0.090 | −0.232 | −0.621 | −0.245 |
| P | 0.834 | 0.192 | 0.864 | 0.245 | 0.818 | 0.548 | 0.075 | 0.217 | |
| Coarse materials | r | 0.063 | 0.396 | −0.015 | 0.271 | −0.083 | 0.409 | 0.295 | 0.251 |
| P | 0.873 | 0.292 | 0.969 | 0.172 | 0.831 | 0.275 | 0.441 | 0.206 | |
| Bare soil | r | −0.275 | 0.264 | 0.059 | 0.068 | −0.225 | −0.336 | 0.399 | −0.123 |
| P | 0.473 | 0.493 | 0.881 | 0.735 | 0.561 | 0.377 | 0.288 | 0.540 | |
| Total vegetation cover | r | 0.632 | 0.639 | −0.403 | 0.157 | 0.204 | −0.278 | −0.215 | 0.020 |
| P | 0.068 | 0.064 | 0.282 | 0.433 | 0.598 | 0.470 | 0.579 | 0.920 | |
| pH | r | 0.549 | −0.133 | 0.522 | −0.021 | −0.191 | −0.072 | −0.573 | −0.234 |
| P | 0.126 | 0.733 | 0.149 | 0.916 | 0.622 | 0.854 | 0.107 | 0.240 | |
| Electrical conductivity | r | 0.275 | 0.540 | 0.169 | 0.337 | −0.523 | −0.390 | −0.878 | −0.224 |
| P | 0.474 | 0.134 | 0.665 | 0.086 | 0.149 | 0.299 | 0.002 | 0.262 | |
| Soil organic carbon | r | −0.276 | 0.240 | 0.642 | −0.153 | −0.087 | −0.557 | 0.032 | −0.095 |
| P | 0.473 | 0.533 | 0.062 | 0.446 | 0.824 | 0.119 | 0.935 | 0.636 | |
| Gravel | r | 0.518 | 0.067 | −0.019 | 0.186 | 0.145 | 0.374 | 0.320 | 0.288 |
| P | 0.153 | 0.864 | 0.961 | 0.353 | 0.709 | 0.321 | 0.401 | 0.145 | |
| Sand | r | −0.235 | −0.026 | −0.062 | −0.109 | 0.141 | −0.295 | −0.234 | −0.192 |
| P | 0.543 | 0.948 | 0.873 | 0.589 | 0.717 | 0.441 | 0.545 | 0.336 | |
| Silt | r | −0.386 | −0.225 | 0.333 | −0.166 | −0.195 | −0.575 | −0.652 | −0.230 |
| P | 0.305 | 0.560 | 0.382 | 0.407 | 0.615 | 0.105 | 0.057 | 0.249 | |
| Clay | r | −0.515 | −0.211 | 0.354 | −0.184 | −0.306 | −0.449 | −0.558 | −0.240 |
| P | 0.156 | 0.586 | 0.350 | 0.357 | 0.423 | 0.225 | 0.118 | 0.229 | |
| Biodiversity Information | Upstream | Midstream | Downstream | Overall |
|---|---|---|---|---|
| Samples | 9 | 9 | 9 | 27 |
| Individuals (computed) | 1307 | 685 | 919 | 2942 |
| S(est) ± SD | 39 ± 1.82 | 21 ± 3.01 | 15 ± 0 | 42 ± 2.06 |
| S(est) 95% CI lower bound | 35.43 | 15.09 | 15 | 37.96 |
| S(est) 95% CI upper bound | 42.57 | 26.91 | 15 | 46.04 |
| Singletons | 3 | 3 | 0 | 3 |
| Doubletons | 1 | 0 | 0 | 2 |
| Uniques | 7 | 6 | 0 | 7 |
| Duplicates | 6 | 1 | 1 | 5 |
| ACE | 40.64 | 24.12 | 15 | 43.93 |
| ICE | 43.17 | 25.32 | 15 | 47.48 |
| S(Chao 1) | 40.5 | 24 | 15 | 43 |
| Chao 1 95% CI lower bound | 39.15 | 21.35 | 15 | 42.09 |
| Chao 1 95% CI upper bound | 54.07 | 46.66 | 15.48 | 52.68 |
| Chao 1 SD (analytical) | 2.6 | 4.57 | 0.22 | 1.82 |
| S(Chao 2) | 41.7 | 27.7 | 15 | 45.4 |
| Chao 2 95% CI lower bound | 39.48 | 22.17 | 15 | 42.62 |
| Chao 2 95% CI upper bound | 53.95 | 59.12 | 16.06 | 60.3 |
| Chao 2 SD (analytical) | 2.88 | 7.32 | 0.41 | 3.54 |
| S(Jack 1) ± SD | 45.22 ± 3.2 | 26.33 ± 1.89 | 15 ± 0 | 48.74 ± 2.97 |
| S(Jack 2) | 46.6 | 30.3 | 15 | 50.8 |
| Bootstrap mean | 42.18 | 23.3 | 15.14 | 45.34 |
| MMRuns mean | 44.35 | 25.22 | 16.3 | 44.54 |
| MMMeans (1 run) | 43.99 | 24.6 | 16.29 | 44.51 |
| Cole rarefaction | 39 | 21 | 15 | 42 |
| Alpha mean | 7.56 | 4.1 | 2.55 | 6.94 |
| Alpha SD (analytical) | 0.55 | 0.41 | 0.27 | 0.43 |
| Shannon mean | 3.07 | 2.48 | 2.14 | 3.09 |
| Shannon exponential mean | 21.44 | 11.96 | 8.53 | 22.03 |
| Simpson inv mean | 15.34 | 8.98 | 6.03 | 16.02 |
| Watershed Sampling Site | S1 (S = 39) | S1 (S = 39) | S2 (S = 21) |
|---|---|---|---|
| S2 (S = 21) | S3 (S = 15) | S3 (S = 15) | |
| Shared species observed | 19 | 14 | 10 |
| ACE first sample | 40.64 | 40.64 | 24.13 |
| ACE second sample | 24.13 | 15 | 15 |
| Chao shared estimated | 21.43 | 14 | 0 |
| Classic Jaccard index (%) | 46.3 | 35.0 | 38.5 |
| Classic Sørensen index (%) | 63.3 | 51.9 | 55.6 |
| Raw Chao–Jaccard index (%) | 57.5 | 48.2 | 31.9 |
| Estimated Chao–Jaccard index (%) | 57.8 | 48.2 | 32.6 |
| Raw Chao–Sørensen index (%) | 73.0 | 65.0 | 48.4 |
| Estimated Chao–Sørensen index (%) | 73.3 | 65.0 | 49.1 |
| Morisita–Horn index (%) | 45.0 | 35.5 | 22.7 |
| Bra–Curtis index (%) | 40.2 | 35.1 | 23.3 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Souahi, H.; Gacem, R.; Chenchouni, H. Variation in Plant Diversity along a Watershed in the Semi-Arid Lands of North Africa. Diversity 2022, 14, 450. https://doi.org/10.3390/d14060450
Souahi H, Gacem R, Chenchouni H. Variation in Plant Diversity along a Watershed in the Semi-Arid Lands of North Africa. Diversity. 2022; 14(6):450. https://doi.org/10.3390/d14060450
Chicago/Turabian StyleSouahi, Hana, Rania Gacem, and Haroun Chenchouni. 2022. "Variation in Plant Diversity along a Watershed in the Semi-Arid Lands of North Africa" Diversity 14, no. 6: 450. https://doi.org/10.3390/d14060450