
The Importance of Forest Elephants for Vegetation Structure Modification and Its Influence on the Bird Community of a Mid-Elevation Forest on Mount Cameroon, West-Central Africa

 , and
, and
Abstract
:1. Introduction
2. Material and Methods
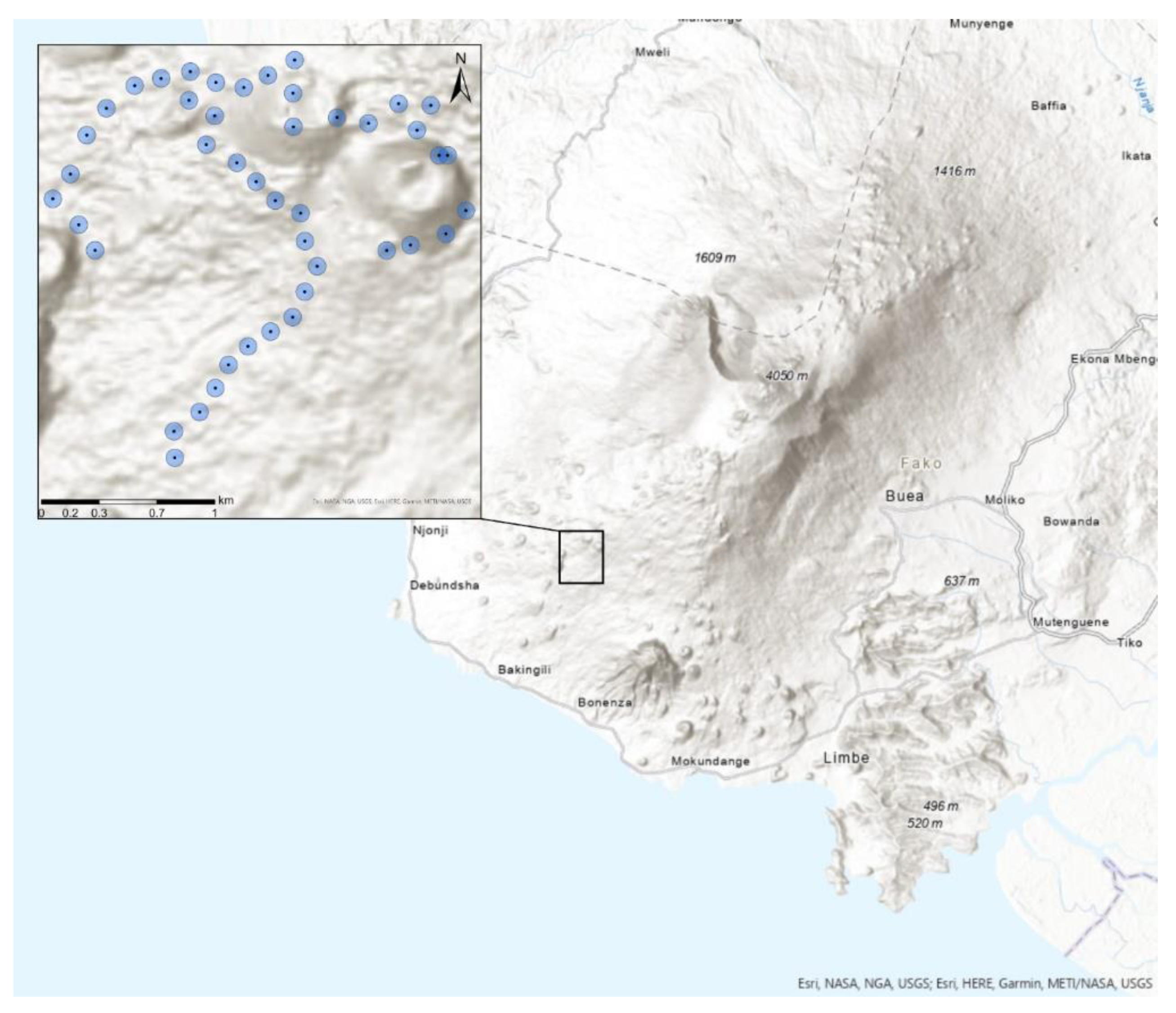
2.1. Study Area
2.2. Bird Diversity Survey
2.2.1. Vegetation Sampling
2.2.2. Statistical Analyses
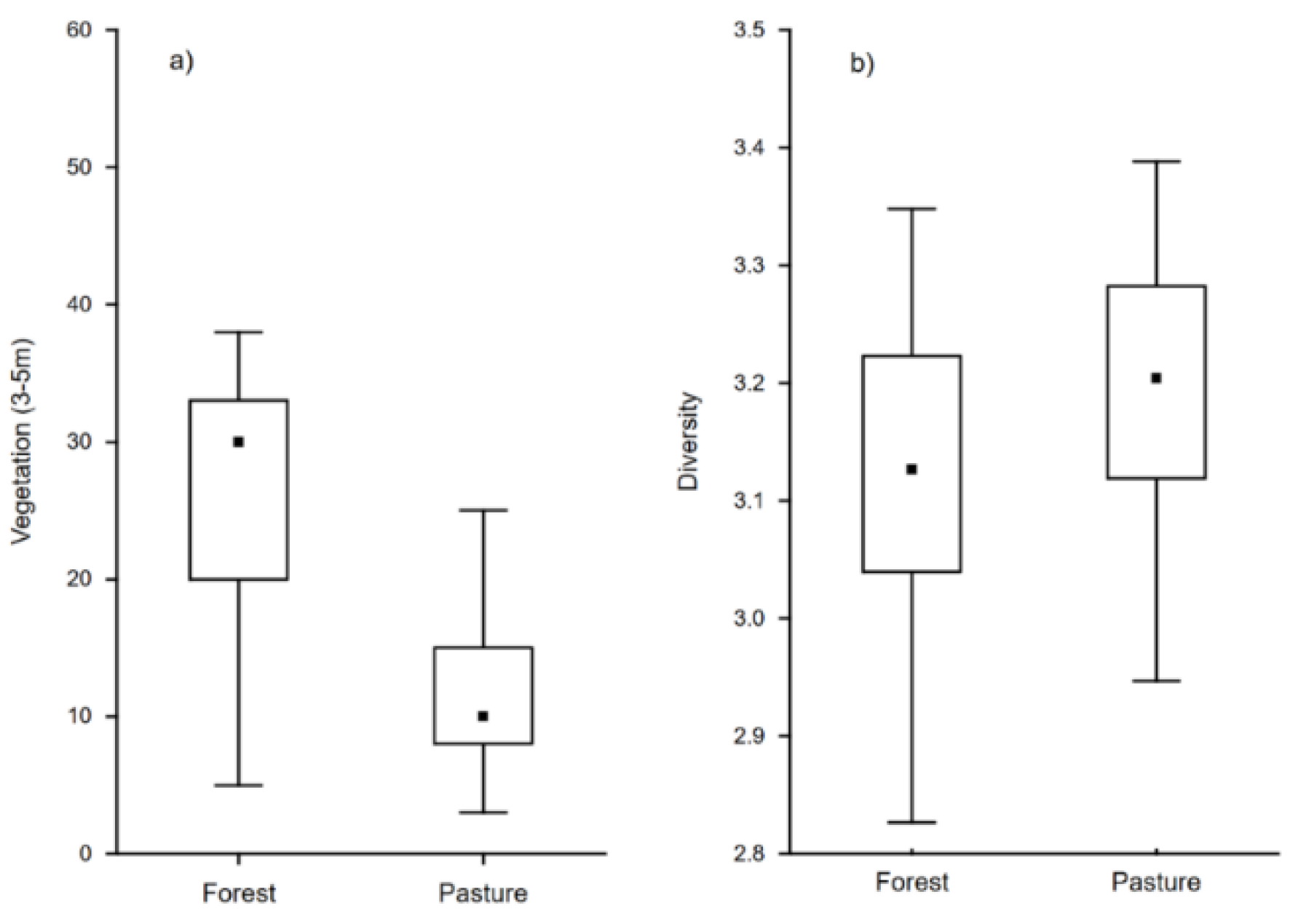
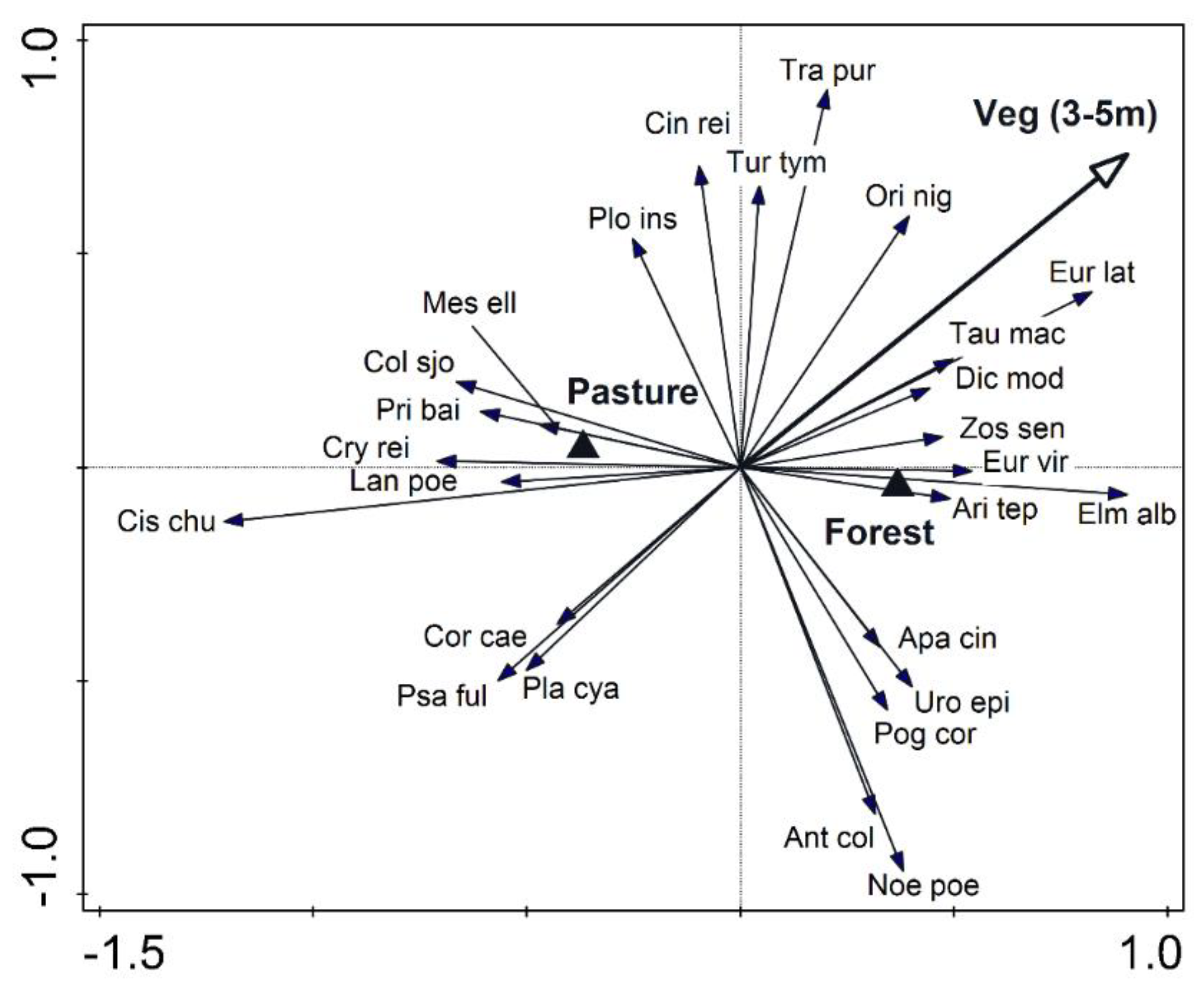
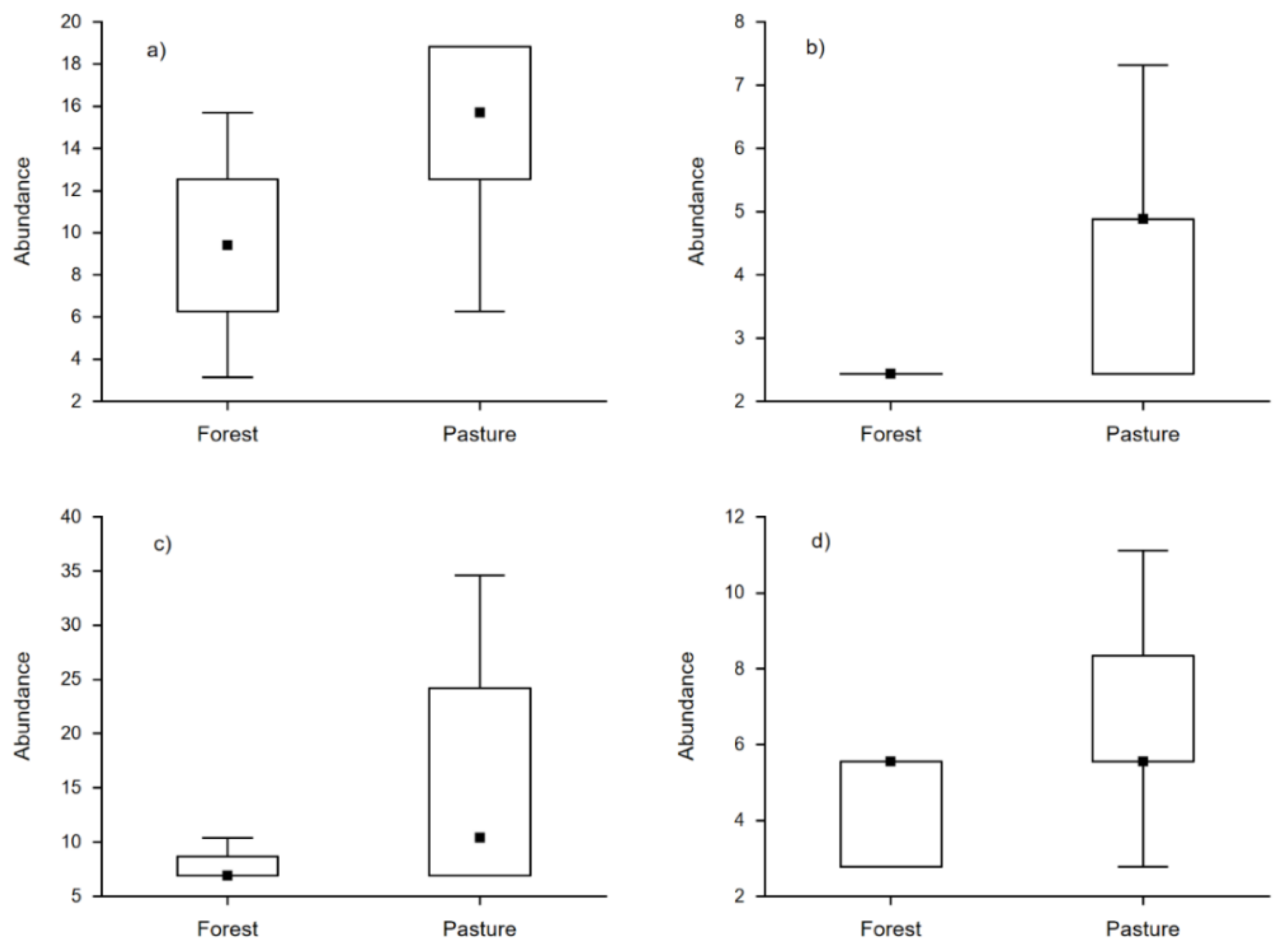
3. Results
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Foley, J.A.; DeFries, R.; Asner, G.P.; Barford, C.; Bonan, G.; Carpenter, S.R.; Snyder, P.K. Global consequences of land use. Science 2005, 309, 570–574. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brown, J.H. Why are there so many species in the tropics? J. Biogeogr. 2014, 41, 8–22. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cuni-Sanchez, A.; Sullivan, M.J.; Platts, P.J.; Lewis, S.L.; Marchant, R.; Imani, G.; Hubau, W.; Abiem, I.; Adhikari, H.; Albrecht, T.; et al. High aboveground carbon stock of African tropical montane forests. Nature 2021, 596, 536–542. [Google Scholar] [CrossRef] [PubMed]
- Zakaria, M.; Rajpar, M.N.; Ozdemir, I.; Rosli, Z. Tropical Forests-the Challenges of Maintaining Ecosystem Services While Managing the Landscape, 3rd ed.; InTech: Janeza Tedine, Croatia, 2016; pp. 11–49. [Google Scholar]
- Godfray, H.C.J.; Beddington, J.R.; Crute, I.R.; Haddad, L.; Lawrence, D.; Muir, J.F.; Toulmin, C. Food security: The challenge of feeding 9 billion people. Science 2010, 327, 812–818. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Laurance, W.F. Habitat destruction: Death by a thousand cuts. In Conservation Biology for All; Sodhi, N.S., Ehrlich, P.R., Eds.; Oxford Scholarship Online; Oxford University Press: Oxford, UK, 2010; pp. 73–88. [Google Scholar]
- Margono, B.A.; Potapov, P.V.; Turubanova, S.; Stolle, F.; Hansen, M.C. Primary Forest cover loss in Indonesia over 2000–2012. Nat. Clim. Chang. 2014, 4, 730–735. [Google Scholar] [CrossRef]
- Achard, F.; Eva, H.D.; Stibig, H.J.; Mayaux, P.; Gallego, J.; Richards, T.; Malingreau, J.P. Determination of deforestation rates of the world’s humid tropical forests. Science 2002, 297, 999–1002. [Google Scholar] [CrossRef] [Green Version]
- Simberloff, D. Global climate change and introduced species in United States forests. Sci. Total Environ. 2000, 262, 253–261. [Google Scholar] [CrossRef]
- Ibarra, J.T.; Martin, K. Biotic homogenization: Loss of avian functional richness and habitat specialists in disturbed Andean temperate forests. Biol. Conserv. 2015, 192, 418–427. [Google Scholar] [CrossRef]
- Shahabuddin, G.; Kumar, R. Influence of anthropogenic disturbance on bird communities in a tropical dry forest: Role of vegetation structure. Anim. Conserv. 2006, 9, 404–413. [Google Scholar] [CrossRef]
- Flynn, D.F.; Gogol-Prokurat, M.; Nogeire, T.; Molinari, N.; Richers, B.T.; Lin, B.B.; Simpson, N.; Mayfield, M.M.; DeClerck, F. Loss of functional diversity under land use intensification across multiple taxa. Ecol. Lett. 2009, 12, 22–33. [Google Scholar] [CrossRef]
- Sheta, B.; Orabi, G.; Bedir, M.; El-bokl, M.; Habbk, L. Impact of Some Anthropogenic Activities on the Diversity of Resident Bird Species at Damietta Region, Egypt. Catrina 2011, 6, 59–74. [Google Scholar]
- Altaf, M.; Javid, A.; Khan, A.M.; Khan, M.S.H.; Umair, M.; Ali, Z. Anthropogenic impact on the distribution of the birds in the tropical thorn forest, Punjab, Pakistan. J. Asia-Pac. Biodivers. 2018, 11, 229–236. [Google Scholar] [CrossRef]
- Matuoka, M.A.; Benchimol, M.; de Almeida-Rocha, J.M.; Morante-Filho, J.C. Effects of anthropogenic disturbances on bird functional diversity: A global meta-analysis. Ecol. Indic. 2020, 116, 106471. [Google Scholar] [CrossRef]
- Mohd-Taib, F.S.; Mohd-Saleh, W.; Asyikha, R.; Mansor, M.S.; Ahmad-Mustapha, M.; Mustafa-Bakray, N.A.; Sulaiman, N. Effects of anthropogenic disturbance on the species assemblages of birds in the back mangrove forests. Wetl. Ecol. Manag. 2020, 28, 479–494. [Google Scholar] [CrossRef]
- Sousa, W.P. The role of disturbance in natural communities. Annual Review of Ecology and Systematics 1984, 15, 353–391. [Google Scholar] [CrossRef]
- Parminter, J. Natural disturbance ecology. In Conservation Biology Principles for Forested Landscapes; Voller, J., Harrison, S., Eds.; UBC Press: Vancouver, BC, Canada, 1998; pp. 3–41. [Google Scholar]
- Campos-Arceiz, A.; Blake, S. Megagardeners of the forest–the role of elephants in seed dispersal. Acta Oecol. 2011, 37, 542–553. [Google Scholar] [CrossRef]
- Beaune, D.; Fruth, B.; Bollache, L.; Hohmann, G.; Bretagnolle, F. Doom of the elephant-dependent trees in a Congo tropical forest. For. Ecol. Manag. 2013, 295, 109–117. [Google Scholar] [CrossRef]
- Parker, D.M. The Effects of Elephants at Low Densities and after Short Occupation Time on the Ecosystems of the Eastern Cape Province, South Africa. Ph.D. Thesis, Rhodes University, Grahamstown, South Africa, 2008. [Google Scholar]
- White, L.J.; Tutin, C.E.; Fernandez, M. Group composition and diet of forest elephants, Loxodonta africana cyclotis Matschie 1900, in the Lopé Reserve, Gabon. Afr. J. Ecol. 1993, 3, 181–199. [Google Scholar] [CrossRef]
- Blake, S.; Deem, S.L.; Mossimbo, E.; Maisels, F.; Walsh, P. Forest elephants: Tree planters of the Congo. Biotropica 2009, 41, 459–468. [Google Scholar] [CrossRef]
- Wiafe, E.; Dakwa, K.; Yeboah, S. Assemblages of avian communities in forest elephant (Loxodonta cyclotis) range in Ghana. Pachyderm 2010, 48, 41–47. [Google Scholar]
- Campbell, D.G. Gap formation in tropical forest canopy by elephants, Oveng, Gabon, Central Africa. Biotropica 1991, 23, 195–196. [Google Scholar] [CrossRef]
- Guldemond, R.A.; Purdon, A.; Van Aarde, R.J. A systematic review of elephant impact across Africa. PLoS ONE 2017, 12, e0178935. [Google Scholar] [CrossRef] [PubMed]
- Kerley, G.I.; Landman, M. The impacts of elephants on biodiversity in the Eastern Cape Subtropical Thickets: Elephant conservation. S. Afr. J. Sci. 2006, 102, 395–402. [Google Scholar]
- Okosodo, E.F.; Orimaye, J.O.; Ogunyemi, O.O. Habitat effects on avian species abundance and diversity in Idanre forest reserve, south western Nigeria. Int. J. Plant Anim. Envi. Sci. 2016, 6, 1–18. [Google Scholar]
- Herremans, M. Effects of woodland modification by African elephant Loxodonta Africana on bird diversity in northern Botswana. Ecography 1995, 18, 440–454. [Google Scholar] [CrossRef]
- Déruelle, B.; N’ni, J.; Kambou, R. Mount Cameroon: An active volcano of the Cameroon Line. J. Afr. Earth Sci. 1987, 6, 197–214. [Google Scholar] [CrossRef]
- MINFOF. The Management Plan of the Mount Cameroon National Park and Its Peripheral Zone; Action plan; MINFOF: Yaounde, Cameroon, 2014. [Google Scholar]
- Cheek, M.; Cable, S.; Hepper, F.N.; Ndam, N.; Watts, J. The Biodiversity of African Plants, 3rd ed.; Kluwer Academic Publishers: Wageningen, The Netherlands, 1996; pp. 110–120. [Google Scholar]
- IUCN. The IUCN Red List of Threatened Species. Version 2021-3; Available online: https://www.iucnredlist.org (accessed on 10 November 2021).
- Nkengfack, S.N. Biodiversity Conservation, Ecotourism and Rural Livelihoods in Protected Areas. Case Study: The Mount Cameroon National Park. Master’s Thesis, Norwegian University of Life Sciences, Ås, Norway, 2012. [Google Scholar]
- Dudley, J.P.; Mensah-Ntiamoah, A.Y.; Kpelle, D.G. Forest elephants in a rainforest fragment: Preliminary findings from a wildlife conservation project in southern Ghana. Afr. J. Ecol. 1992, 30, 116–126. [Google Scholar] [CrossRef]
- Ekobo, A. Survey of Large Mammals in the Mount Cameroon Forest; Unpublished Report; GTZ Mount Cameroon Project, Environmental Information and Documentation Centre: Buea, Cameroon, 2003. [Google Scholar]
- White, A.M.; Goodman, P.S. Differences in woody vegetation are unrelated to use by African elephants (Loxodonta africana) in Mkhuze Game Reserve, South Africa. Afr. J. Ecol. 2009, 48, 215–223. [Google Scholar] [CrossRef]
- Shannon, G.; Druce, D.J.; Page, B.R.; Eckhardt, H.C.; Grant, R.; Slotow, R. The utilization of large savanna trees by elephant in southern Kruger National Park. J. Trop. Ecol. 2008, 24, 281–289. [Google Scholar] [CrossRef]
- Monadjem, A.; Garcelon, D.K. Nesting distribution of vultures in relation to land use in Swaziland. Biodivers. Conserv. 2005, 14, 2079–2093. [Google Scholar] [CrossRef]
- Chmel, K.; Kamga, S.M.; Awa, T., II; Ewome, F.L.; Uceda-Gómez, G.; Hořák, D.; Mlíkovský, J.; Molua, L.L.; Riegert, J.; Janeček, Š. Vertical stratification and seasonal changes of the avian community in Mount Cameroon lowland rainforest. Afr. J. Ecol. 2021, 59, 655–666. [Google Scholar] [CrossRef]
- Chmel, K.; Riegert, J.; Paul, L.; Novotný, V. Vertical stratification of an avian community in New Guinean tropical rainforest. Popul. Ecol. 2016, 58, 535–547. [Google Scholar] [CrossRef]
- Ayonghe, A. Rural communities and protected area management in the Mount Cameroon Region of West Africa. Afr. J. Econ. Sustain. Dev. 2012, 1, 146–165. [Google Scholar] [CrossRef]
- Ndam, N.; Acworth, J.; Kenfack, D.; Tchouto, M.G.P.; Hall, J. Plant Diversity Assessment on Mount Cameroon: Surveys from 1990 to 2000. Sys. Geogr. Plants 2001, 71, 1017–1022. [Google Scholar] [CrossRef]
- Fonge, B.A.; Yinda, G.S.; Focho, D.A.; Fongod, A.G.N.; Bussmann, R.W. Vegetation and soil status on an 80 year old lava flow of Mt. Cameroon, West Africa. Lyonia 2005, 8, 17–39. [Google Scholar]
- Payton, R.W. Ecology, Altitudinal Zonation and Conservation of Tropical Rainforest of Mt Cameroon; Report to the Overseas Development Administration: London, UK, 1993; p. 251. [Google Scholar]
- Tynsong, H.; Tiwari, B.K. Diversity of plant species in arecanut agroforests of south Meghalaya, north-east India. J. For. Res. 2010, 21, 281–286. [Google Scholar] [CrossRef]
- Bibby, C.J.; Burgess, N.D.; Hillis, D.M.; Hill, D.A.; Mustoe, S. Bird Census Techniques, 2nd ed.; Academic Press: London, UK, 2000. [Google Scholar]
- Johnson, D.H. Point counts of birds: What are we estimating? In Monitoring Bird Populations by Point Counts; Ralph, C.J., Sauer, J.R., Droege, S., Eds.; General Technical Reports PSW-GTR-149; US Department of Agriculture, Forest Service, Pacific Southwest Research Station: Albany, CA, USA, 1995; pp. 117–124, 149. [Google Scholar]
- Sutherland, W.J.; Newton, I.; Green, R.E. Bird Ecology and Conservation: A Handbook of Techniques, 3rd ed.; Oxford University Press: New York, NY, USA, 2004; p. 386. [Google Scholar]
- TIBCO Software Inc. Statistica (Data Analysis Software System), Version 13. 2017. Available online: http://statistica.io (accessed on 7 November 2021).
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2020; Available online: https://www.R-project.org/ (accessed on 7 November 2021).
- McDonald, T.; Nielson, R.; McDonald, A.; Nielson, R.; Augustine, B.; Griswald, J.; McKann, P.; Jeroue, L.; Hoffman, A.; Klainsausser, M.; et al. Package ‘Rdistance’. Microsoft R Application Network. 2015. Available online: https://mran.microsoft.com/snapshot/2018-09-09/web/packages/Rdistance/Rdistance.pdf (accessed on 18 August 2021).
- Simpson, E.H. Measurement of diversity. Nature 1949, 163, 688. [Google Scholar] [CrossRef]
- Jaccard, P. Gesetze der Pflanzenverteilung in der alpinen Region. Flora 1902, 90, 349–377. [Google Scholar]
- Shannon, C.E. A mathematical theory of communication. Bell Syst. Tech. J. 1948, 27, 379–423. [Google Scholar] [CrossRef] [Green Version]
- ter Braak, C.J.; Smilauer, P. Canoco Reference Manual and User’s Guide: Software for Ordination; Version 5.0; Microcomputer Power: Ithaca, NY, USA, 2012. [Google Scholar]
- Marrot, P.; Garant, D.; Charmantier, A. Spatial autocorrelation in fitness affects the estimation of natural selection in the wild. Methods Ecol. Evol. 2015, 6, 1474–1483. [Google Scholar] [CrossRef] [Green Version]
- Legendre, P.; Legendre, L. Numerical Ecology, 3rd ed.; Elsevier: Amsterdam, The Netherlands, 2012. [Google Scholar]
- Šmilauer, P.; Lepš, J. Multivariate Analysis of Ecological Data Using CANOCO 5; Cambridge University Press: Cambridge, UK, 2014. [Google Scholar]
- Okland, B. Unlogged forests: Important sites for preserving the diversity of mycetophilids (Diptera: Sciaroidea). Biol. Conserv. 1996, 76, 297–310. [Google Scholar] [CrossRef]
- Murcia, C. Edge effects in fragmented forests: Implications for conservation. Trends Ecol. Evol. 1995, 10, 58–62. [Google Scholar] [CrossRef]
- Steffan-Dewenter, I.; Tscharntke, T. Butterfly community structure in fragmented habitats. Ecol. Lett. 2000, 3, 449–456. [Google Scholar] [CrossRef]
- Ralph, C.J. Habitat association patterns of forest and steppe birds of Northern Patagonia, Argentina. Condor 1985, 87, 471–483. [Google Scholar] [CrossRef]
- Sullivan, T.P.; Sullivan, D.S. Influence of variable retention harvests on forest ecosystems. II. Diversity and population dynamics of small mammals. J. Appl. Ecol. 2001, 38, 1234–1252. [Google Scholar] [CrossRef]
- Davidowitz, G.; Rosenzweig, M.L. The latitudinal gradient of species diversity among North American grasshoppers within a single habitat: A test of the spatial heterogeneity hypothesis. J. Biogeogr. 1998, 25, 553–560. [Google Scholar] [CrossRef]
- Tews, J.; Brose, U.; Grimm, V.; Tielbörger, K.; Wichmann, M.C.; Schwager, M.; Jeltsch, F. Animal species diversity driven by habitat heterogeneity/diversity: The importance of keystone structures. J. Biogeogr. 2004, 31, 79–92. [Google Scholar] [CrossRef] [Green Version]
- Greenberg, C.H.; Lanham, J.D. Breeding bird assemblages of hurricane-created gaps and adjacent closed canopy forest in the southern Appalachians. For. Ecol. Manag. 2001, 154, 251–260. [Google Scholar] [CrossRef] [Green Version]
- Bazzaz, F.A. Plant species diversity in old-field successional ecosystems in southern Illinois. Ecology 1975, 56, 485–488. [Google Scholar] [CrossRef]
- Scherber, C.; Vockenhuber, E.A.; Stark, A.; Meyer, H.; Tscharntke, T. Effects of tree and herb biodiversity on Diptera, a hyperdiverse insect order. Oecologia 2014, 174, 1387–1400. [Google Scholar] [CrossRef]
- Hassan, S.N.; Salum, A.R.; Rija, A.A.; Modest, R.; Kideghesho, J.R.; Malata, P.F. Human-Induced Disturbances Influence on Bird Communities of Coastal Forests in Eastern Tanzania. 2013. Available online: http://www.suaire.sua.ac.tz/handle/123456789/303 (accessed on 9 November 2021).
- Stuart, S.N. Conservation of Cameroon Montane Forests. International Council for Bird Preservation: Cambridge, UK, 1986.
- Reif, J.; Sedláček, O.; Hořák, D.; Riegert, J.; Pešata, M.; Hrázský, Z.; Janeček, Š. Habitat preferences of birds in a montane forest mosaic in the Bamenda Highlands, Cameroon. Ostrich 2007, 78, 31–36. [Google Scholar] [CrossRef]
- Borrow, N.; Demey, R. Birds of Western Africa, 2nd ed.; Bloomsbury Publishing Plc: London, UK, 2014; p. 592. [Google Scholar]
- Reif, J.; Hořák, D.; Sedláček, O.; Riegert, J.; Pešata, M.; Hrázský, Z.; Janeček, Š.; Storch, D. Unusual abundance-range size relationship in an Afromontane bird community: The effect of geographic isolation? J. Biogeogr. 2006, 33, 1959–1968. [Google Scholar] [CrossRef]
- MacArthur, R.H.; MacArthur, J.W. On bird species diversity. Ecology 1961, 42, 594–598. [Google Scholar] [CrossRef]
- Johnston, V.R. Breeding birds of the forest edge in Illinois. Condor 1947, 49, 45–53. [Google Scholar] [CrossRef]
- Batáry, P.; Fronczek, S.; Normann, C.; Scherber, C.; Tscharntke, T. How do edge effect and tree species diversity change bird diversity and avian nest survival in Germany’s largest deciduous forest? For. Ecol. Manag. 2014, 319, 44–50. [Google Scholar] [CrossRef]
- Sisk, T.D.; Battin, J. Habitat edges and avian ecology: Geographic patterns and insights for western landscapes. J. Avian Biol. 2002, 25, 30–48. [Google Scholar]
- Robinson, S.K.; Thompson, F.R., III; Donovan, T.M.; Whitehead, D.R.; Faaborg, J. Regional Forest fragmentation and the nesting success of migratory birds. Science 1995, 267, 1987–1990. [Google Scholar] [CrossRef] [Green Version]
- Batáry, P.; Baldi, A. Evidence of an edge effect on avian nest success. Conserv. Biol. 2004, 18, 389–400. [Google Scholar] [CrossRef]
- Djomo, N.E.; Sedláček, O.; Doležal, J.; Dančák, M.; Altman, J.; Svoboda, M.; Hořák, D. Relationship between survival rate of avian artificial nests and forest vegetation structure along a tropical altitudinal gradient on Mount Cameroon. Biotropica 2015, 47, 758–764. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variable | U | p |
|---|---|---|
| Layer 0–1 m | 21.5 | 0.001 |
| Layer 1–3 m | 233.5 | 0.851 |
| Layer 3–5 m | 62.5 | 0.001 |
| Layer 5–10 m | 63.5 | 0.001 |
| Layer >10 m | 48.5 | 0.001 |
| % of bushes | 194 | 0.264 |
| Variables | Layer 2 | Layer 3 | Layer 4 | Layer 5 | % of Bushes |
|---|---|---|---|---|---|
| Layer 1 | −0.17 | −0.56 | −0.56 | −0.60 | 0.33 |
| Layer 2 | 0.44 | 0.18 | −0.03 | 0.13 | |
| Layer 3 | 0.74 | 0.49 | −0.07 | ||
| Layer 4 | 0.76 | −0.06 | |||
| Layer 5 | −0.17 |
| Forest | Pasture | |
|---|---|---|
| Species richness | 22.64 ± 3.87 (15–29) | 27.00 ± 3.16 (21–32) |
| Simpson index | 0.07 ± 0.01 (0.05–0.10) | 0.07 ± 0.01 (0.05–0.11) |
| Jaccard index | 0.43 ± 0.11 (0.24–0.83) | 0.54 ± 0.08 (0.31–0.77) |
| Contribution % | Pseudo-F | p | |
|---|---|---|---|
| Vegetation layer (3–5 m) | 12.3 | 2.6 | 0.006 |
| Habitat | 7.5 | 1.6 | 0.036 |
| PCO.3 | 10.7 | 2.9 | 0.006 |
| PCO.4 | 8.8 | 2.5 | 0.002 |
| PCO.7 | 7.1 | 2 | 0.008 |
| PCO.6 | 6.3 | 1.8 | 0.016 |
| Bird Species | U | p |
|---|---|---|
| Cisticola chubbi | 56.5 | 0.003 |
| Laniarus poensis | 36 | 0.013 |
| Onychognathus walleri | 55 | 0.017 |
| Pseudoalcippe abyssinica | 123.5 | 0.044 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kamga, S.M.; Tamungang, S.A.; Awa, T., II; Ewome, F.L.; Motombi, F.N.; Hořák, D.; Riegert, J. The Importance of Forest Elephants for Vegetation Structure Modification and Its Influence on the Bird Community of a Mid-Elevation Forest on Mount Cameroon, West-Central Africa. Diversity 2022, 14, 227. https://doi.org/10.3390/d14030227
Kamga SM, Tamungang SA, Awa T II, Ewome FL, Motombi FN, Hořák D, Riegert J. The Importance of Forest Elephants for Vegetation Structure Modification and Its Influence on the Bird Community of a Mid-Elevation Forest on Mount Cameroon, West-Central Africa. Diversity. 2022; 14(3):227. https://doi.org/10.3390/d14030227
Chicago/Turabian StyleKamga, Solange Mekuate, Simon Awafor Tamungang, Taku Awa, II, Francis Luma Ewome, Francis Njie Motombi, David Hořák, and Jan Riegert. 2022. "The Importance of Forest Elephants for Vegetation Structure Modification and Its Influence on the Bird Community of a Mid-Elevation Forest on Mount Cameroon, West-Central Africa" Diversity 14, no. 3: 227. https://doi.org/10.3390/d14030227