
Evaluating Potential Distribution and Niche Divergence among Populations of the World’s Largest Living Damselfly, Megaloprepus caerulatus (Drury, 1782)


{kind=link}
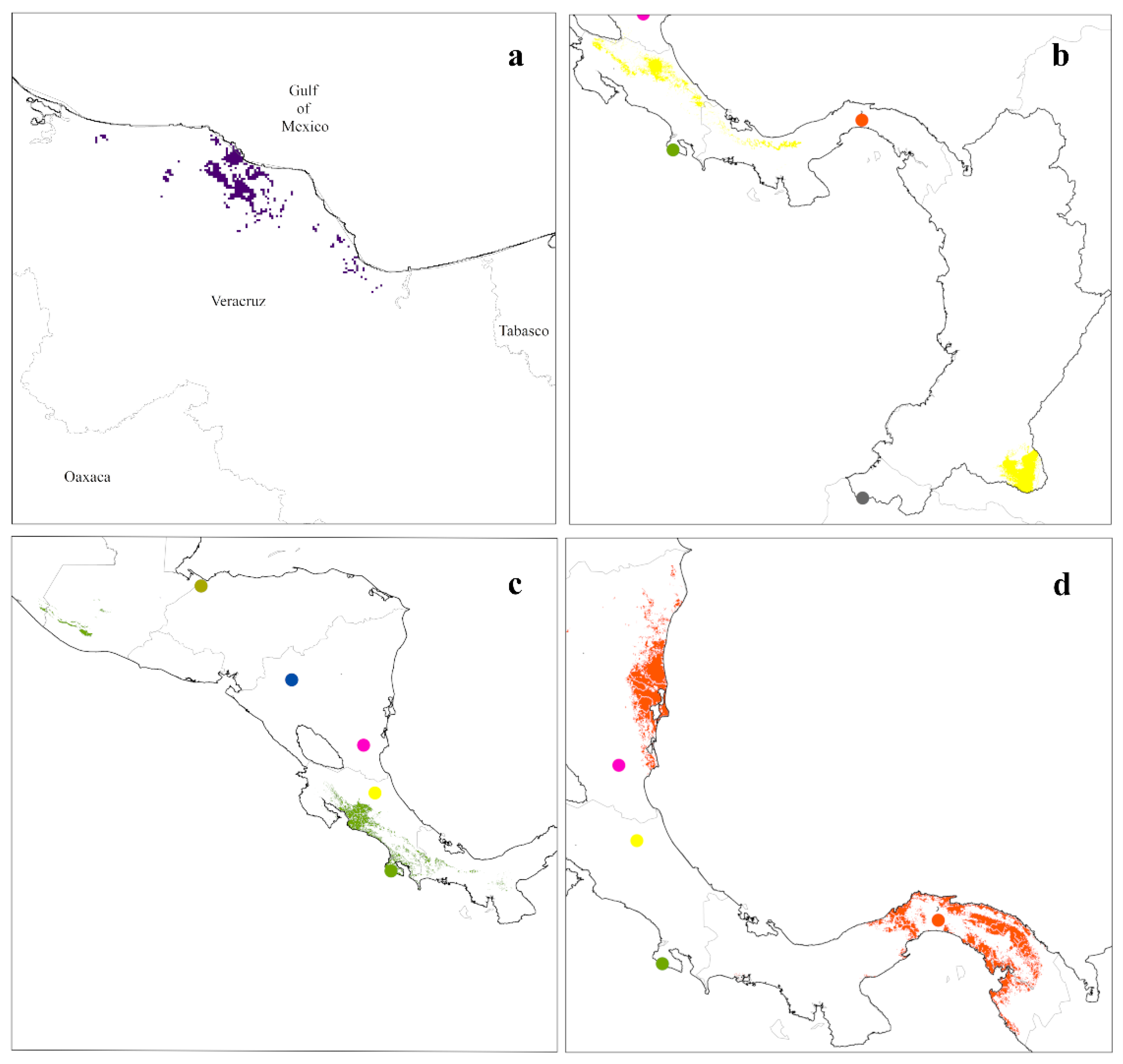{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Biological and Environmental Data
2.2. Ecological Niche Modelling
2.3. Niche Similarity Tests
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Grinnell, J. The niche-relationships of the California Thrasher. Auk 1917, 34, 427–433. [Google Scholar] [CrossRef]
- Soberón, J.; Peterson, A.T. Interpretation of models of fundamental ecological niches and species’ distributional areas. Biodivers. Inform. 2005, 2, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Soberón, J. Grinnellian and Eltonian niches and geographic distributions of species. Ecol. Lett. 2007, 10, 1115–1123. [Google Scholar] [CrossRef]
- Barve, N.; Martin, C.; Brunsell, N.A.; Peterson, A.T. The role of physiological optima in shaping the geographic distribution of Spanish moss. Glob. Ecol. Biogeogr. 2014, 23, 633–645. [Google Scholar] [CrossRef]
- Peterson, A.T.; Soberón, J.; Pearson, R.G.; Anderson, R.P.; Martínez-Meyer, E.; Nakamura, M.; Araújo, M.B. Ecological Niches and Geographic Distributions; Princeton University Press: Princeton, NJ, USA, 2011; p. 314. [Google Scholar]
- Jackson, S.T.; Overpeck, J.T. Responses of plant populations and communities to environmental changes of the late Quaternary. Paleobiology 2000, 26, 194–220. [Google Scholar] [CrossRef]
- Peterson, A.T. Predicting species’ geographic distributions based on ecological niche modeling. Condor 2001, 103, 599–605. [Google Scholar] [CrossRef]
- Ortega-Huerta, M.A.; Peterson, A.T. Modeling ecological niches and predicting geographic distributions: A test of six presence-only methods. Rev. Mex. Biodivers. 2008, 79, 205–216. [Google Scholar]
- Elith, J.; Leathwick, J.R. Species distribution models: Ecological explanation and prediction across space and time. Annu. Rev. Ecol. Evol. Syst. 2009, 40, 677–697. [Google Scholar] [CrossRef]
- Smith, A.B.; Godsoe, W.; Rodríguez-Sánchez, F.; Wang, H.H.; Warren, D. Niche estimation above and below the species level. Trends Ecol. Evol. 2019, 34, 260–273. [Google Scholar] [CrossRef]
- Collins, S.D.; McIntyre, N.E. Modeling the distribution of odonates: A review. Freshw. Sci. 2015, 34, 1144–1158. [Google Scholar] [CrossRef]
- Costa Bastos, R.; Schlemmer Brasil, L.; Oliveira-Junior, J.M.B.; Geraldo Carvalho, F.; Lennox, G.D.; Barlow, J.; Juen, L. Morphological and phylogenetic factors structure the distribution of damselfly and dragonfly species (Odonata) along an environmental gradient in Amazonian streams. Ecol. Indic. 2021, 122, 107257. [Google Scholar] [CrossRef]
- Rodríguez-Tapia, G.; Rocha-Ortega, M.; Córdoba-Aguilar, A. An index to estimate the vulnerability of damselflies and dragonflies (Insecta: Odonata) to land use changes using niche modeling. Aquat. Insects 2020, 41, 254–272. [Google Scholar] [CrossRef]
- Boys, W.A.; Siepielski, A.M.; Smith, B.D.; Patten, M.A.; Bried, J.T. Predicting the distributions of regional endemic dragonflies using a combined model approach. Insect Conserv. Divers. 2021, 14, 52–66. [Google Scholar] [CrossRef]
- Ackerly, D.D. Community assembly, niche conservatism, and adaptive evolution in changing environments. Int. J. Plant Sci. 2003, 164, S165–S184. [Google Scholar] [CrossRef]
- Balanyá, J.; Oller, J.M.; Huey, R.B.; Gilchrist, G.W.; Serra, L. Global genetic change tracks global climate warming in Drosophila subobscura. Science 2006, 313, 1773–1775. [Google Scholar] [CrossRef] [PubMed]
- Holt, R.D. The microevolutionary consequences of climate change. Trends Ecol. Evol. 1990, 5, 311–315. [Google Scholar] [CrossRef]
- Peterson, A.T.; Soberón, J.; Sánchez-Cordero, V. Conservatism of ecological niches in evolutionary time. Science 1999, 285, 1265–1267. [Google Scholar] [CrossRef]
- Martínez-Meyer, E.; Peterson, A.T.; Hargrove, W.W. Ecological niches as stable distributional constraints on mammal species, with implications for Pleistocene extinctions and climate change projections for biodiversity. Glob. Ecol. Biogeogr. 2004, 13, 305–314. [Google Scholar] [CrossRef]
- Jakob, S.S.; Ihlow, A.; Blattner, F.R. Combined ecological niche modelling and molecular phylogeography revealed the evolutionary history of Hordeum marinum (Poaceae)—Niche differentiation, loss of genetic diversity, and speciation in Mediterranean Quaternary refugia. Mol. Ecol. 2007, 16, 1713–1727. [Google Scholar] [CrossRef]
- Pearman, P.B.; Guisan, A.; Broennimann, O.; Randin, C.F. Niche dynamics in space and time. Trends Ecol. Evol. 2008, 23, 149–158. [Google Scholar] [CrossRef]
- Pearman, P.B.; D’Amen, M.; Graham, C.H.; Thuiller, W.; Zimmermann, N.E. Within-taxon niche structure: Niche conservatism, divergence and predicted effects of climate change. Ecography 2010, 33, 990–1003. [Google Scholar] [CrossRef]
- Brown, J.L.; Carnaval, A.C. A tale of two niches: Methods, concepts, and evolution. Front. Biogeogr. 2019, 11, e44158. [Google Scholar] [CrossRef] [Green Version]
- Encarnación-Luévano, A.; Peterson, A.T.; Rojas-Soto, O.R. Burrowing habit in Smilisca frogs as an adaptive response to ecological niche constraints in seasonally dry environments. Front. Biogeogr. 2021, 13, e50517. [Google Scholar] [CrossRef]
- Serra-Varela, M.J.; Grivet, D.; Vincenot, L.; Broennimann, O.; Gonzalo-Jiménez, J.; Zimmermann, N.E. Does phylogeographical structure relate to climatic niche divergence? A test using maritime pine (Pinus pinaster Ait.). Glob. Ecol. Biogeogr. 2015, 24, 1302–1313. [Google Scholar] [CrossRef]
- Banta, J.A.; Ehrenreich, I.M.; Gerard, S.; Chou, L.; Wilczek, A.; Schmitt, J.; Kover, P.X.; Purugganan, M.D. Climate envelope modelling reveals intraspecific relationships among flowering phenology, niche breadth and potential range size in Arabidopsis thaliana. Ecol. Lett. 2012, 15, 769–777. [Google Scholar] [CrossRef] [PubMed]
- Marcer, A.; Méndez-Vigo, B.; Alonso-Blanco, C.; Picó, F.X. Tackling intraspecific genetic structure in distribution models better reflects species geographical range. Ecol. Evol. 2016, 6, 2084–2097. [Google Scholar] [CrossRef] [Green Version]
- Mota-Vargas, C.; Rojas-Soto, O.R. Taxonomy and ecological niche modeling: Implications for the conservation of wood partridges (genus Dendrortyx). J. Nat. Conserv. 2016, 29, 1–13. [Google Scholar] [CrossRef]
- Bried, J.T.; Siepielski, A.M. Predator driven niches vary spatially among co-occurring damselfly species. Evol. Ecol. 2019, 33, 243–256. [Google Scholar] [CrossRef]
- Wiens, J.J.; Ackerly, D.D.; Allen, A.P.; Anacker, B.L.; Buckley, L.B.; Cornell, H.V.; Damschen, E.I.; Jonathan Davies, T.; Grytnes, J.A.; Harrison, S.P.; et al. Niche conservatism as an emerging principle in ecology and conservation biology. Ecol. Lett. 2010, 13, 1310–1324. [Google Scholar] [CrossRef]
- Fincke, O.M.; Xu, M.; Khazan, E.S.; Wilson, M.; Ware, J.L. Tests of hypotheses for morphological and genetic divergence in Megaloprepus damselflies across Neotropical forests. Biol. J. Linn. Soc. 2018, 125, 844–861. [Google Scholar] [CrossRef]
- Eaton, A.E.; Calvert, P.P. Biologia Centrali-Americana: Insecta, Neuroptera, Ephemeridæ & Odonata; Taylor & Francis: London, UK, 1892–1908; p. 420. [Google Scholar]
- Hedström, I.; Sahlén, G. A key to the adult Costa Rican “helicopter” damselflies (Odonata: Pseudostigmatidae) with notes on their phenology and life zone preferences. Rev. Biol. Trop. 2001, 48, 1037–1056. [Google Scholar]
- Fincke, O.M.; Hedström, I. Differences in forest use and colonization by Neotropical tree-hole damselflies (Odonata: Pseudostigmatidae): Implications for forest conversion. Stud. Neotrop. Fauna Environ. 2008, 43, 35–45. [Google Scholar] [CrossRef]
- Feindt, W.; Fincke, O.; Hadrys, H. Still a one species genus? Strong genetic diversification in the world’s largest living odonate, the Neotropical damselfly Megaloprepus caerulatus. Conserv. Genet. 2014, 15, 469–481. [Google Scholar] [CrossRef]
- Young, A.M. Feeding and oviposition in the Giant Tropical Damselfly Megaloprepus coerulatus (Drury) in Costa Rica. Biotropica 1980, 12, 237. [Google Scholar] [CrossRef]
- Fincke, O.M. Giant damselflies in a tropical forest: Reproductive biology of Megaloprepus caerulatus with notes on Mecistogaster (Zygoptera: Pseudostigmatidae). Adv. Odonatol. 1984, 2, 13–27. [Google Scholar]
- Fincke, O.M. Interspecific competition for tree holes: Consequences for mating systems and coexistence in Neotropical damselflies. Am. Nat. 1992, 139, 80–101. [Google Scholar] [CrossRef]
- Fincke, O.M. Use of forest and tree species, and dispersal by giant damselflies (Pseudostigmatidae): Their prospects in fragmented forests. In Forests and Dragonflies, Proceedings of the 4th WDA International Symposium of Odonatology, Pontevedra, Spain, 26–30 July 2005; Cordero-Rivera, A., Ed.; Pensoft Publishers: Sofia, Bulgaria, 2006; pp. 103–125. [Google Scholar]
- Schultz, T.D.; Fincke, O.M. Structural colours create a flashing cue for sexual recognition and male quality in a Neotropical giant damselfly. Funct. Ecol. 2009, 23, 724–732. [Google Scholar] [CrossRef]
- Xu, M.; Fincke, O.M. Ultraviolet wing signal affects territorial contest outcome in a sexually dimorphic damselfly. Anim. Behav. 2015, 101, 67–74. [Google Scholar] [CrossRef]
- de Selys Longchamps, M. Révision du synopsis des Agrionines. Première partie comprenant les légions Pseudostigma—Podagrion—Platycnemis et Protoneura. Mem. Couronnés Académie R. Belg. 1886, 38, 1–233. [Google Scholar]
- Escoto-Moreno, J.A.; Hernández-Hernández, A.; Hernández-Hernández, J.A.; Márquez, J.; Silva-Briano, M.; Novelo-Gutiérrez, R. El registro más septentrional de la libélula gigante neotropical Megaloprepus caerulatus (Drury, 1782) (Odonata: Coenagrionidae) en el continente Americano. Gayana 2018, 82, 90–93. [Google Scholar] [CrossRef] [Green Version]
- Measey, G.J. Some Odonata from Belize, Central America. Not. Odonatol. 1994, 4, 40–46. [Google Scholar]
- González-Soriano, E.; Dirzo, R.; Vogt, R.C. Historia Natural de los Tuxtlas; Instituto de Biología, UNAM: Mexico City, Mexico, 1997; p. 647. [Google Scholar]
- Machado, J. Inventario y Estudio Comparativo de la Fauna de Odonata en tres Áreas de Honduras. Bachelor Thesis, Universidad Zamorano, San Antonio de Oriente, Honduras, November 2001. [Google Scholar]
- Cuevas-Yáñez, K. Los odonatos (Insecta: Odonata) de la Hidroeléctrica de Patla (El Pozo) y del Río Tecpatlán, Zihuateutla, Puebla, México. Dugesiana 2007, 14, 83–91. [Google Scholar]
- González-Soriano, E.; Paulson, D.R. Los odonatos de Chiapas. In Chiapas: Estudios Sobre su Diversidad Biológica; Álvarez-Noguera, F., Ed.; Instituto de Biología, UNAM: Mexico City, Mexico, 2011; pp. 299–314. [Google Scholar]
- Esquivel, C. Las libélulas de la zona de El Rodeo, cantón de Mora, San José, Costa Rica. Brenesia 2012, 77, 329–342. [Google Scholar]
- GBIF. GBIF Occurrence Download. Available online: https://doi.org/10.15468/dl.dqiufw (accessed on 19 August 2019).
- Olson, D.M.; Dinerstein, E.; Wikramanayake, E.D.; Burgess, N.D.; Powell, G.V.N.; Underwood, E.C.; D’amico, J.A.; Itoua, I.; Strand, H.E.; Morrison, J.C.; et al. Terrestrial ecoregions of the World: A new Map of Life on Earth: A new global map of terrestrial ecoregions provides an innovative tool for conserving biodiversity. Bioscience 2001, 51, 933–938. [Google Scholar] [CrossRef]
- Corbet, P.S. Dragonflies: Behaviour and Ecology of Odonata (Revised Edition), 2nd ed.; Comstock Publishing Associates: New York, NY, USA, 2004; p. 829. [Google Scholar]
- Corbet, P.S.; Brooks, S.J. Dragonflies; Collins: London, UK, 2008; p. 454. [Google Scholar]
- Cuevas-Yáñez, K.; Rivas, M.; Muñoz, J.; Córdoba-Aguilar, A. Conservation status assessment of Paraphlebia damselflies in Mexico. Insect Conserv. Divers. 2015, 8, 517–524. [Google Scholar] [CrossRef]
- Rangel-Sánchez, L.; Nava-Bolaños, A.; Palacino-Rodríguez, F.; Córdoba-Aguilar, A. Estimating distribution area in six Argia damselflies (Insecta: Odonata: Coenagrionidae) including A. garrisoni, a threatened species. Rev. Mex. Biodivers. 2018, 89, 921–926. [Google Scholar] [CrossRef]
- Hijmans, R.J.; Cameron, S.E.; Parra, J.L.; Jones, P.G.; Jarvis, A. Very high resolution interpolated climate surfaces for global land areas. Int. J. Climatol. 2005, 25, 1965–1978. [Google Scholar] [CrossRef]
- Tuanmu, M.N.; Jetz, W. A global 1-km consensus land-cover product for biodiversity and ecosystem modelling. Glob. Ecol. Biogeogr. 2014, 23, 1031–1045. [Google Scholar] [CrossRef]
- Trabucco, A.; Zomer, R.J. Global High-Resolution Soil-Water Balance. Figshare Fileset. Available online: https://figshare.com/articles/Global_High-Resolution_Soil-Water_Balance/7707605/3 (accessed on 18 November 2019).
- Danielson, J.J.; Gesch, D.B. Global Multi-Resolution Terrain Elevation Data 2010 (GMTED2010): U.S. Geological Survey Open-File Report 2011–1073; U.S. Geological Survey: Reston, VA, USA, 2011; p. 26. [Google Scholar]
- Lehner, B.; Verdin, K.; Jarvis, A. New global hydrography derived from spaceborne elevation data. Eos 2008, 89, 93–94. [Google Scholar] [CrossRef]
- Harrell, F.E. Hmisc: Harrell Miscellaneous. R Package Version 4.3-0. Available online: https://CRAN.R-project.org/package=Hmisc (accessed on 13 September 2019).
- R Core Team. R: A Language and Environment for Statistical Computing. R Foundation for Statistical Computing, Vienna, Austria. Available online: http://www.R-project.org/ (accessed on 30 May 2020).
- Phillips, S.J.; Anderson, R.P.; Schapire, R.E. Maximum entropy modeling of species geographic distributions. Ecol. Modell. 2006, 190, 231–259. [Google Scholar] [CrossRef] [Green Version]
- Peterson, A.T.; Soberón, J. Species distribution modeling and ecological niche modeling: Getting the Concepts Right. Nat. Conserv. 2012, 10, 102–107. [Google Scholar] [CrossRef]
- Barve, N.; Barve, V.; Jiménez-Valverde, A.; Lira-Noriega, A.; Maher, S.P.; Peterson, A.T.; Soberón, J.; Villalobos, F. The crucial role of the accessible area in ecological niche modeling and species distribution modeling. Ecol. Modell. 2011, 222, 1810–1819. [Google Scholar] [CrossRef]
- Warren, D.L.; Glor, R.E.; Turelli, M. Environmental niche equivalency versus conservatism: Quantitative approaches to niche evolution. Evolution 2008, 62, 2868–2883. [Google Scholar] [CrossRef] [PubMed]
- Peterson, A.T. Ecological niche conservatism: A time-structured review of evidence. J. Biogeogr. 2011, 38, 817–827. [Google Scholar] [CrossRef]
- Broennimann, O.; Fitzpatrick, M.C.; Pearman, P.B.; Petitpierre, B.; Pellissier, L.; Yoccoz, N.G.; Thuiller, W.; Fortin, M.J.; Randin, C.; Zimmermann, N.E.; et al. Measuring ecological niche overlap from occurrence and spatial environmental data. Glob. Ecol. Biogeogr. 2012, 21, 481–497. [Google Scholar] [CrossRef] [Green Version]
- Owens, H.L.; Campbell, L.P.; Dornak, L.L.; Saupe, E.E.; Barve, N.; Soberón, J.; Ingenloff, K.; Lira-Noriega, A.; Hensz, C.M.; Myers, C.E.; et al. Constraints on interpretation of ecological niche models by limited environmental ranges on calibration areas. Ecol. Modell. 2013, 263, 10–18. [Google Scholar] [CrossRef]
- Liu, C.; White, M.; Newell, G. Selecting thresholds for the prediction of species occurrence with presence-only data. J. Biogeogr. 2013, 40, 778–789. [Google Scholar] [CrossRef]
- Lehner, B.; Grill, G. Global river hydrography and network routing: Baseline data and new approaches to study the world’s large river systems. Hydrol. Process. 2013, 27, 2171–2186. [Google Scholar] [CrossRef]
- Peterson, A.T.; Papeş, M.; Soberón, J. Rethinking receiver operating characteristic analysis applications in ecological niche modeling. Ecol. Modell. 2008, 213, 63–72. [Google Scholar] [CrossRef]
- Osorio-Olvera, L.; Lira-Noriega, A.; Soberón, J.; Peterson, A.T.; Falconi, M.; Contreras-Díaz, R.G.; Martínez-Meyer, E.; Barve, V.; Barve, N. ntbox: An r package with graphical user interface for modelling and evaluating multidimensional ecological niches. Methods Ecol. Evol. 2020, 11, 1199–1206. [Google Scholar] [CrossRef]
- Pearson, R.G.; Raxworthy, C.J.; Nakamura, M.; Townsend Peterson, A. Predicting species distributions from small numbers of occurrence records: A test case using cryptic geckos in Madagascar. J. Biogeogr. 2007, 34, 102–117. [Google Scholar] [CrossRef]
- Schoener, T.W. Nonsynchronous spatial overlap of lizards in patchy habitats. Ecology 1970, 51, 408–418. [Google Scholar] [CrossRef] [Green Version]
- Aguirre-Gutiérrez, J.; Serna-Chavez, H.M.; Villalobos-Arambula, A.R.; Pérez de la Rosa, J.A.; Raes, N. Similar but not equivalent: Ecological niche comparison across closely-related Mexican white pines. Divers. Distrib. 2015, 21, 245–257. [Google Scholar] [CrossRef]
- Di Cola, V.; Broennimann, O.; Petitpierre, B.; Breiner, F.T.; D’Amen, M.; Randin, C.; Engler, R.; Pottier, J.; Pio, D.; Dubuis, A.; et al. ecospat: An R package to support spatial analyses and modeling of species niches and distributions. Ecography 2017, 40, 774–787. [Google Scholar] [CrossRef]
- Halffter, G.; Morrone, J.J. An analytical review of Halffter’s Mexican transition zone, and its relevance for evolutionary biogeography, ecology and biogeographical regionalization. Zootaxa 2017, 4226, 1–46. [Google Scholar] [CrossRef]
- Morrone, J.J. Biogeographic regionalization and biotic evolution of Mexico: Biodiversity’s crossroads of the New World. Rev. Mex. Biodivers. 2019, 90, 1–68. [Google Scholar] [CrossRef]
- Rzedowski, J. El endemismo en la flora fanerogámica mexicana: Una apreciación analítica preliminar. Acta Bot. Mex. 1991, 47–64. [Google Scholar] [CrossRef] [Green Version]
- Dinerstein, E.; Olson, D.; Joshi, A.; Vynne, C.; Burgess, N.D.; Wikramanayake, E.; Hahn, N.; Palminteri, S.; Hedao, P.; Noss, R.; et al. An ecoregion-based approach to protecting half the terrestrial realm. Bioscience 2017, 67, 534–545. [Google Scholar] [CrossRef]
- Morrone, J.J. Biogeografía de América Latina y el Caribe, M&T–Manuales & Tesis SEA, Vol. 3; CYTED, UNESCO-ORCYT & SEA: Zaragoza, Spain, 2001; 148p. [Google Scholar]
- Avise, J.C.; Walker, D. Pleistocene phylogeographic effects on avian populations and the speciation process. Proc. R. Soc. B Biol. Sci. 1998, 265, 457–463. [Google Scholar] [CrossRef] [Green Version]
- Morrone, J.J. Biogeographical regionalisation of the neotropical region. Zootaxa 2014, 3782, 1–110. [Google Scholar] [CrossRef] [Green Version]
- Haffer, J. Speciation in amazonian forest birds. Science 1969, 165, 131–137. [Google Scholar] [CrossRef] [PubMed]
- Bush, M.B.; de Oliveira, P.E. The rise and fall of the Refugial Hypothesis of Amazonian speciation: A paleoecological perspective. Biota Neotrop. 2006, 6, bn00106012006. [Google Scholar] [CrossRef] [Green Version]
- Fincke, O.M. Consequences of larval ecology for territoriality and reproductive success of a neotropical damselfly. Ecology 1992, 73, 449–462. [Google Scholar] [CrossRef]
- Eaton, M.D.; Soberón, J.; Peterson, A.T. Phylogenetic perspective on ecological niche evolution in american blackbirds (Family Icteridae). Biol. J. Linn. Soc. 2008, 94, 869–878. [Google Scholar] [CrossRef] [Green Version]
- Petitpierre, B.; Kueffer, C.; Broennimann, O.; Randin, C.; Daehler, C.; Guisan, A. Climatic niche shifts are rare among terrestrial plant invaders. Science 2012, 335, 1344–1348. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Toussaint, E.F.A.; Bybee, S.M.; Erickson, R.J.; Condamine, F.L. Forest giants on different evolutionary branches: Ecomorphological convergence in helicopter damselflies. Evolution 2019, 73, 1045–1054. [Google Scholar] [CrossRef]
- Ingley, S.J.; Bybee, S.M.; Tennessen, K.J.; Whiting, M.F.; Branham, M.A. Life on the fly: Phylogenetics and evolution of the helicopter damselflies (Odonata, Pseudostigmatidae). Zool. Scr. 2012, 41, 637–650. [Google Scholar] [CrossRef]
- Paulson, D.R. Middle American Odonata By Country. Slater Museum of Natural History, University of Puget Sound. Available online: https://www.pugetsound.edu/academics/academic-resources/slater-museum/biodiversity-resources/dragonflies/middle-american-odonata/ (accessed on 3 September 2020).
- Marshall, L.G.; Webb, S.D.; Sepkoski, J.J.; Raup, D.M. Mammalian evolution and the great American interchange. Science 1982, 215, 1351–1357. [Google Scholar] [CrossRef]
- Rich, P.V.; Rich, T.H. The Central American dispersal route: Biotic history and palaeogeography. In Costa Rican Natural History; Janzen, D.H., Ed.; The University of Chicago Press: Chicago, IL, USA, 1983; pp. 12–34. [Google Scholar]
- Montes, C.; Cardona, A.; Jaramillo, C.; Pardo, A.; Silva, J.C.; Valencia, V.; Ayala, C.; Pérez-Angel, L.C.; Rodriguez-Parra, L.A.; Ramirez, V.; et al. Middle Miocene closure of the Central American Seaway. Science 2015, 348, 226–229. [Google Scholar] [CrossRef] [Green Version]
- Hoorn, C.; Flantua, S. An early start for the Panama land bridge. Science 2015, 348, 186–187. [Google Scholar] [CrossRef]
- Halffter, G. Biogeography of the Montane Entomofauna of Mexico and Central America. Annu. Rev. Entomol. 1987, 32, 95–114. [Google Scholar] [CrossRef]
- Morrone, J.J. Homología Biogeográfica: Las Coordenadas Espaciales de la Vida. Cuadernos del Instituto de Biología 37; Instituto de Biología, UNAM: Mexico City, Mexico, 2004; p. 199. [Google Scholar]
- Brown, K.S. Areas where humid tropical forest probably persisted. In Biogeography and Quaternary History in Tropical America; Whitmore, T.C., Prance, G.T., Eds.; Clarendon Press: Oxford, UK, 1987; pp. 44–45. [Google Scholar]
- Hiebeler, D. Populations on fragmented landscapes with spatially structured heterogeneities: Landscape generation and local dispersal. Ecology 2000, 81, 1629. [Google Scholar] [CrossRef]
- Suárez-Tovar, C.M.; Rocha-Ortega, M.; González-Voyer, A.; González-Tokman, D.; Córdoba-Aguilar, A. The larger the damselfly, the more likely to be threatened: A sexual selection approach. J. Insect Conserv. 2019, 23, 535–545. [Google Scholar] [CrossRef]
- Rocha-Ortega, M.; Rodríguez, P.; Bried, J.; Abbott, J.; Córdoba-Aguilar, A. Why do bugs perish? Range size and local vulnerability traits as surrogates of Odonata extinction risk. Proc. R. Soc. B Biol. Sci. 2020, 287, 20192645. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leiva, F.P.; Calosi, P.; Verberk, W.C.E.P. Scaling of thermal tolerance with body mass and genome size in ectotherms: A comparison between water- and air-breathers. Philos. Trans. R. Soc. B Biol. Sci. 2019, 374, 20190035. [Google Scholar] [CrossRef] [Green Version]
- Rocha-Ortega, M.; Rodríguez, P.; Córdoba-Aguilar, A. Can dragonfly and damselfly communities be used as bioindicators of land use intensification? Ecol. Indic. 2019, 107, 105553. [Google Scholar] [CrossRef]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Encarnación-Luévano, A.; Escoto-Moreno, J.A.; Villalobos-Jiménez, G. Evaluating Potential Distribution and Niche Divergence among Populations of the World’s Largest Living Damselfly, Megaloprepus caerulatus (Drury, 1782). Diversity 2022, 14, 84. https://doi.org/10.3390/d14020084
Encarnación-Luévano A, Escoto-Moreno JA, Villalobos-Jiménez G. Evaluating Potential Distribution and Niche Divergence among Populations of the World’s Largest Living Damselfly, Megaloprepus caerulatus (Drury, 1782). Diversity. 2022; 14(2):84. https://doi.org/10.3390/d14020084
Chicago/Turabian StyleEncarnación-Luévano, Alondra, Jaime Antonio Escoto-Moreno, and Giovanna Villalobos-Jiménez. 2022. "Evaluating Potential Distribution and Niche Divergence among Populations of the World’s Largest Living Damselfly, Megaloprepus caerulatus (Drury, 1782)" Diversity 14, no. 2: 84. https://doi.org/10.3390/d14020084