
The Utilityof 28S rDNA for Barcoding of Freshwater Sponges (Porifera, Spongillida)

{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
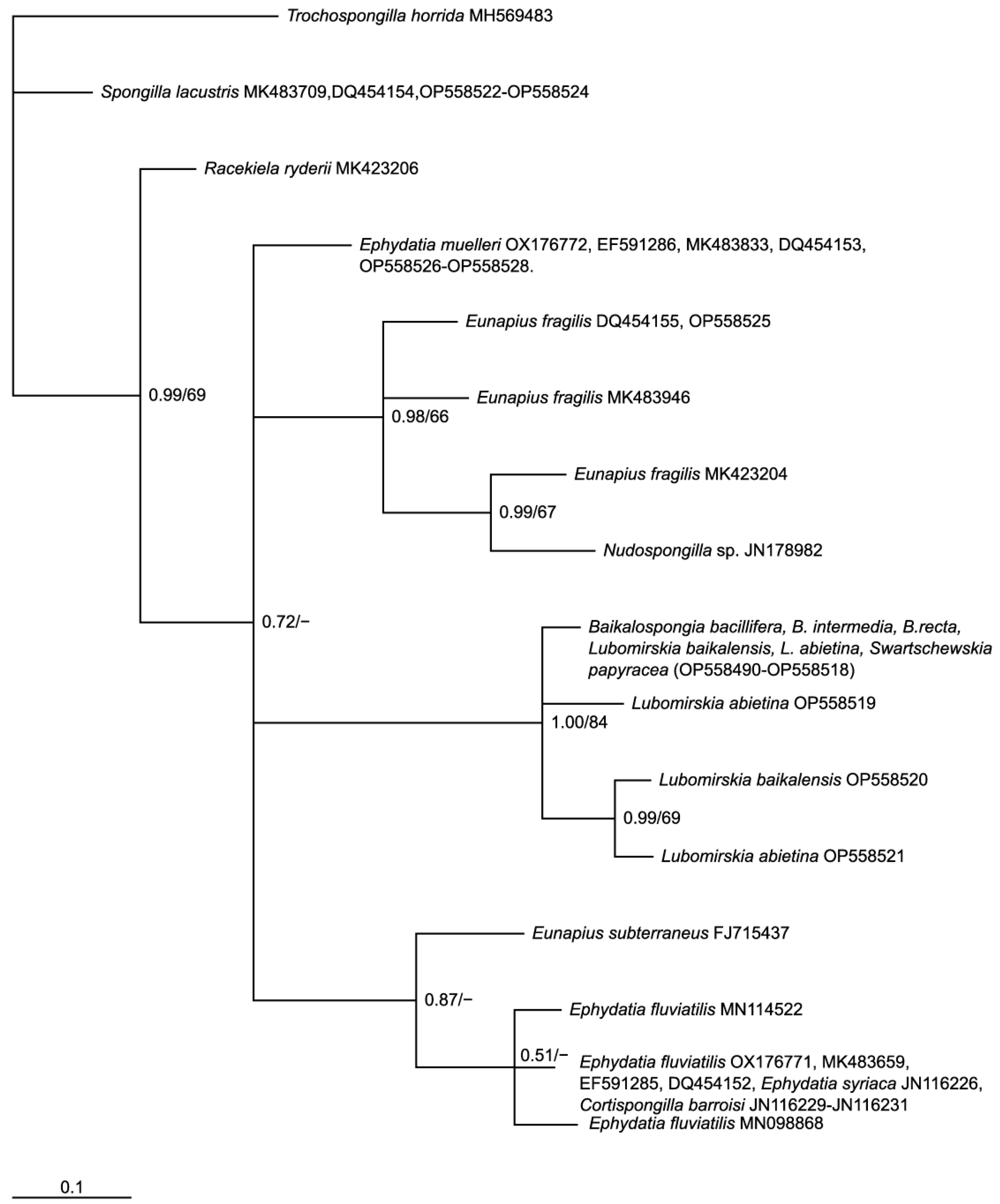
2. Materials and Methods
3. Results and Discussion
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Timoshkin, O.A. Lake Baikal: Fauna diversity, problems of its immiscibility and origin, ecology and exotic communities. In Index of Animal Species Inhabiting Lake Baikal and Its Catchment Area; Timoshkin, O.A., Ed.; Nauka: Lake Baikal, Russia, 2001; Volume 1, pp. 74–116. [Google Scholar]
- Timoshkin, O.A.; Samsonov, D.P.; Yamamuro, M.; Moore, M.V.; Belykh, O.I.; Malnik, V.V.; Sakirko, M.V.; Shirokaya, A.A.; Bondarenko, N.A.; Domysheva, V.M.; et al. Rapid ecological change in the coastal zone of Lake Baikal (East Siberia): Is the site of the world’s greatest freshwater biodiversity in danger? J. Great Lakes Res. 2016, 42, 487–497. [Google Scholar] [CrossRef] [Green Version]
- Khanaev, I.V.; Kravtsova, L.S.; Maikova, O.; Bukshuk, N.; Sakirko, M.V.; Kulakova, N.V.; Butina, T.V.; Nebesnykh, I.A.; Belikov, S.I. Current state of the sponge fauna (Porifera: Lubomirskiidae) of Lake Baikal: Sponge disease and the problem of conservation of diversity. J. Great Lakes Res. 2018, 44, 77–85. [Google Scholar] [CrossRef]
- Itskovich, V.B.; Shigarova, A.M.; Glyzina, O.Y.; Kaluzhnaya, O.V.; Borovskii, G.B. Heat shock protein 70 (Hsp70) response to elevated temperatures in the endemic Baikal sponge Lubomirskiabaicalensis. Ecol. Indic. 2018, 88, 1–7. [Google Scholar] [CrossRef]
- Kaluzhnaya, O.V.; Itskovich, V.B. Features of diversity of polyketide synthase genes in the community of freshwater sponge Baikalospongia fungiformis. Russ. J. Genet. 2022, 58, 336–346. [Google Scholar] [CrossRef]
- Efremova, S.M. Sponges (Porifera). In Index of Animal Species Inhabiting Lake Baikal and Its Catchment Area; Timoshkin, O.A., Ed.; Nauka: Lake Baikal, Russia, 2001; Volume 1, pp. 182–192. [Google Scholar]
- Efremova, S.M. New genus and new species of sponges from family LubomirskiidaeRezvoj, 1936. In Index of Animal Species Inhabiting Lake Baikal and Its Catchment Area; Timoshkin, O.A., Ed.; Nauka: Lake Baikal, Russia, 2004; Volume 1, pp. 1261–1278. [Google Scholar]
- Meixner, M.J.; Luter, C.; Eckert, C.; Itskovich, V.; Janussen, D.; von Rintelen, T.; Bohne, A.V.; Meixner, J.M.; Hess, W.R. Phylogenetic analysis of freshwater sponges provide evidence for endemism and radiation in ancient lakes. Mol. Phyl. Evol. 2007, 45, 875–886. [Google Scholar] [CrossRef] [PubMed]
- Itskovich, V.; Gontcharov, A.; Masuda, Y.; Nohno, T.; Belikov, S.; Efremova, S.; Meixner, M.; Janussen, D. Ribosomal ITS sequences allow resolution of freshwater sponge phylogeny with alignments guided by secondary structure prediction. J. Mol. Evol. 2008, 67, 608–620. [Google Scholar] [CrossRef]
- Itskovich, V.B.; Kaluzhnaya, O.V.; Belikov, S.I. Investigation of nuclear and mitochondrial DNA polymorphism in closely related species of endemic Baikal sponges. Russ. J. Genet. 2013, 49, 966–974. [Google Scholar] [CrossRef]
- Erpenbeck, D.; Steiner, M.; Schuster, A.; Genner, M.J.; Manconi, R.; Pronzato, R.; Ruthensteiner, B.; van den Spiegel, D.; van Soest, R.W.M.; Worheide, G. Minimalist barcodes for sponges: A case study classifying African freshwater Spongillida. Genome 2019, 62, 1–10. [Google Scholar] [CrossRef]
- Kenny, N.J.; Plese, B.; Riesgo, A.; Itskovich, V.B. Symbiosis, selection and novelty: Freshwater adaptation in the unique sponges of Lake Baikal. Mol. Biol. Evol. 2019, 36, 2462–2480. [Google Scholar] [CrossRef]
- Kenny, N.J.; Itskovich, V.B. Phylogenomic inference of the interrelationships of Lake Baikal sponges. Syst. Biodivers. 2021, 19, 209–217. [Google Scholar] [CrossRef]
- Manconi, R.; Pronzato, R. Suborder Spongillinasubord. nov.: Freshwater sponges. In Systema Porifera. A Guide to the Classification of Sponges; Hooper, J.N.A., Van Soest, R.W.M., Eds.; Kluwer Academic/Plenum Publishers: New York, NY, USA; Boston, MA, USA; Dordrecht, The Netherland; London, UK; Moscow, Russia, 2002; pp. 921–1020. [Google Scholar] [CrossRef]
- Masuda, Y. Studies on the taxonomy and distribution of freshwater sponges in Lake Baikal. In Biosilica in Evolution, Morphogenesis and Nanobiotechnology; Müller, W.E.G., Grachev, M.A., Eds.; Springer: Berlin/Heidelberg, Germany, 2009; pp. 81–110. [Google Scholar] [CrossRef]
- Itskovich, V.; Glyzina, O.; Kaluzhnaya, O. Intraspecific and interspecific sequence variability in the ITS region of the rDNA of freshwater sponges of Lake Baikal and East Siberia. Inland Water 2017, 7, 259–266. [Google Scholar] [CrossRef]
- Itskovich, V.B.; Kaluzhnaya, O.V.; Veynberg, E.; Erpenbeck, D. Endemic Lake Baikal sponges from deep water. 1: Potential cryptic speciation and discovery of living species known only from fossils. Zootaxa 2015, 3990, 123–137. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schroder, H.C.; Efremova, S.M.; Itskovich, V.B.; Belikov, S.; Masuda, Y.; Krasko, A.; Muller, I.M.; M€uller, W.E.G. Molecular phylogeny of the freshwater sponges in Lake Baikal. J. Zool. Syst. Evol. Res. 2003, 41, 80–86. [Google Scholar] [CrossRef]
- Erpenbeck, D.; Weier, T.; De Voogd, N.J.; Worheide, G.; Sutcliffe, P.; Todd, J.A.; Michel, E. Insights into the evolution of freshwater sponges (Porifera: Demospongiae: Spongillina): Barcoding and phylogenetic data from Lake Tanganyika endemics indicate multiple invasions and unsettle existing taxonomy. Mol. Phyl. Evol. 2011, 61, 231–236. [Google Scholar] [CrossRef] [PubMed]
- Yakhnenko, A.S.; Itskovich, V.B. Analysis of the variability of mtDNA in closely related species of Baikalian sponges for the development of new barcoding markers. Limnology 2020, 21, 49–57. [Google Scholar] [CrossRef]
- Sokolova, A.M.; Palatov, D.M.; Itskovich, V.B. Genetic analysis confirms the freshwater origin of the endemic Caspian sponges (Demospongiae, Spongillida, Metschnikowiidae). ZooKeys 2020, 915, 1–16. [Google Scholar] [CrossRef] [Green Version]
- Carballo, J.L.; Cruz-Barraza, J.A.; Yanez, B.; Gomez, P. Taxonomy and molecular systematic position of freshwater genus Racekiela (Porifera: Spongillida) with the description of a new species from North-west Mexico. Syst. Biodivers. 2018, 16, 160–170. [Google Scholar] [CrossRef]
- Itskovich, V.B. Intragenomic variation of rDNA internal transcribed spacers in the endemic Baikal sponge Lubomirskiabaicalensis: Implications for Porifera barcoding. J. Great Lakes Res. 2020, 46, 62–66. [Google Scholar] [CrossRef]
- Naumenko, S.A.; Logacheva, M.D.; Popova, N.V.; Klepikova, A.V.; Penin, A.A.; Bazykin, G.A.; Etingova, A.E.; Mugue, N.S.; Kondrashov, A.S.; Yampolsky, L.Y. Transcriptome-based phylogeny of endemic Lake Baikal amphipod species flock: Fast speciation accompanied by frequent episodes of positive selection. Mol. Ecol. 2017, 26, 536–553. [Google Scholar] [CrossRef]
- Adamowicz, S.J.; Marinone, M.C.; Menu-Marque, S.; Martin, J.W.; Allen, D.C.; Pyle, M.N.; De los Ríos, P.; Sobel, C.N.; Ibáñez, C.; Pinto, J.; et al. The Hyalella (Crustacea: Amphipoda) species cloud of the ancient Lake Titicaca originated from multiple colonizations. Mol. Phyl. Evol. 2018, 125, 232–242. [Google Scholar] [CrossRef] [Green Version]
- Fassio, G.; Modica, M.V.; Alvaro, M.C.; Buge, B.; Salvi, D.; Oliverio, M.; Schiaparelli, S. An Antarctic flock under the Thorson’s rule: Diversity and larval development of Antarctic Velutinidae (Mollusca: Gastropoda). Mol. Phyl. Evol. 2019, 132, 1–13. [Google Scholar] [CrossRef]
- Mindell, D.; Honeycutt, R. Ribosomal RNA in Vertebrates: Evolution and phylogenetic applications. Annu. Rev. Ecol. Syst. 1990, 21, 541–566. [Google Scholar] [CrossRef]
- McInerney, J.O.; Adams, C.I.; Kelly, M. Phylogenetic resolution potential of 18S and 28S rRNA genes within the lithistid Astrophorida. Mem. Queensl. Mus. 1999, 44, 343–351. [Google Scholar]
- McCormack, G.P.; Erpenbeck, D.; van Soest, R.W.M. Major discrepancy between phylogenetic hypotheses based on molecular and morphological criteria within the Order Haplosclerida (Phylum Porifera: Class Demospongiae). J. Zool. Syst. Evol. Res. 2002, 40, 237–240. [Google Scholar] [CrossRef]
- Redmond, N.E.; Raleigh, J.; van Soest, R.W.; Kelly, M.; Travers, S.A.; Bradshaw, B.; Vartia, S.; Stephens, K.M.; McCormack, G.P. Phylogenetic relationships of the marine Haplosclerida (Phylum Porifera) employing ribosomal (28S rRNA) and mitochondrial (cox1, nad1) gene sequence data. PLoS ONE 2011, 6, e24344. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morrow, C.; Allcock, L.A.; Mccormack, G. A new species of Hymeraphia Bowerbank, 1864 (Axinellida: Raspailiidae) from a deep-water canyon southwest off Ireland. Zootaxa 2018, 4466, 61–68. [Google Scholar] [CrossRef]
- Erpenbeck, D.; Breeuwer, J.A.J.; van Soest, R.W.M. Implications from a 28S rRNA gene fragment for the phylogenetic relationships of halichondrid sponges (Porifera: Demospongiae). J. Zool. Syst. Evol. Res. 2005, 43, 93–99. [Google Scholar] [CrossRef]
- Erpenbeck, D.; Hooper, J.N.A.; List-Armitage, S.E.; Degnan, B.M.; Wörheide, G.; van Soest, R.W.M. Affinities of the family Sollasellidae (Porifera, Demospongiae). II. Molecular evidence. Contrib. Zool. 2007, 76, 95–102. [Google Scholar] [CrossRef] [Green Version]
- Lopp, A.; Reintamm, T.; Vallmann, K.; Päri, M.; Mikli, V.; Richelle-Maurer, E.; Kelve, M. Molecular identificati on characterization and distribution of freshwater sponges (Porifera: Spongillidae) in Estonia. Fundam. Appl. Limnol. (Archiv für Hydrobiologie) 2007, 168, 93–103. [Google Scholar] [CrossRef]
- Roovere, T.; Lopp, A.; Reintamm, T.; Kuusksalu, A.; Richelle-Maurer, E.; Kelve, M. Freshwater sponges in Estonia: Genetic and morphological identification. Proc. Estonian Acad. Sci. Biol. Ecol. 2006, 55, 216–227. [Google Scholar] [CrossRef]
- Hall, T.A. BioEdit: A user-friendly biological sequence alignment editor and analysis program for Windows 95/98/ NT. Nucl. Acids Symp. Ser. 1999, 41, 95–98. [Google Scholar] [CrossRef]
- Thompson, J.D.; Higgins, D.G.; Gibson, T.J. CLUSTAL W: Improving the sensitivity of progressive multiple sequence alignment through sequence weighting, positions-specific gap penalties and weight matrix choice. Nucl. Acids Res. 1994, 22, 4673–4680. [Google Scholar] [CrossRef] [Green Version]
- Tamura, K.; Peterson, D.; Peterson, N.; Stecher, G.; Nei, M.; Kumar, S. MEGA5: Molecular evolutionary genetics analysis using maximum likelihood, evolutionary distance, and maximum parsimony methods. Mol. Biol. Evol. 2011, 28, 2731–2739. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Felsenstein, J. Confidence limits on phylogenies: An approach using the bootstrap. Evolution 1985, 39, 783–791. [Google Scholar] [CrossRef]
- Ronquist, F.; Huelsenbeck, J.P. MrBayes 3: Bayesian phylogenetic inference under mixed models. Bioinformatics 2003, 19, 1572–1574. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huelsenbeck, J.P.; Rannala, B. Frequentist properties of Bayesian posterior probabilities of phylogenetic trees under simple and complex substitution models. Syst. Biol. 2004, 53, 904–913. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Addis, J.S.; Peterson, K.J. Phylogenetic relationships of freshwater sponges (Porifera, Spongillina) inferred from analyses of 18S rDNA, COI mtDNA, and ITS2 rDNA sequences. Zool. Scr. 2005, 34, 549–557. [Google Scholar] [CrossRef]
- Harcet, M.; Bilandžija, H.; Bruvo-Mađarić, B.; Ćetković, H. Taxonomic position of Eunapiussubterraneus (Porifera, Spongillidae) inferred from molecular data—A revised classification needed? Mol. Phyl. Evol. 2010, 54, 1021–1027. [Google Scholar] [CrossRef]
- Itskovich, V.; Kaluzhnaya, O.; Ostrovsky, I.; McCormack, G. The number of endemic species of freshwater sponges (Malawispongiidae; Spongillina; Porifera) from Lake Kinneret is overestimated. J. Zool. Syst. Evol. Res. 2013, 51, 252–257. [Google Scholar] [CrossRef]
- Erpenbeck, D.; Galitz, A.; Wörheide, G.; Albrecht, C.; Pronzato, R.; Manconi, R. Having the balls to colonize—The Ephydatiafluviatilis group and the origin of (ancient) lake “endemic” sponge lineages. J. Great Lakes Res. 2020, 46, 1140–1145. [Google Scholar] [CrossRef]
- Itskovich, V.; Kaluzhnaya, O.; Veynberg, Y.; Erpenbeck, D. Endemic Lake Baikal sponges from deep water. 2: Taxonomy and bathymetric distribution. Zootaxa 2017, 4236, 335–342. [Google Scholar] [CrossRef] [PubMed]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Itskovich, V.; Kaluzhnaya, O.; Glyzina, O. The Utilityof 28S rDNA for Barcoding of Freshwater Sponges (Porifera, Spongillida). Diversity 2022, 14, 1126. https://doi.org/10.3390/d14121126
Itskovich V, Kaluzhnaya O, Glyzina O. The Utilityof 28S rDNA for Barcoding of Freshwater Sponges (Porifera, Spongillida). Diversity. 2022; 14(12):1126. https://doi.org/10.3390/d14121126
Chicago/Turabian StyleItskovich, Valeria, Oxana Kaluzhnaya, and Olga Glyzina. 2022. "The Utilityof 28S rDNA for Barcoding of Freshwater Sponges (Porifera, Spongillida)" Diversity 14, no. 12: 1126. https://doi.org/10.3390/d14121126