
Morphological Variability of Alveolophora antiqua from a Freshwater Early Miocene Paleolake in the Barguzin Valley (Baikal Rift Zone)
 , ,
, ,  , and
, and
Abstract
:1. Introduction
2. Materials and Methods
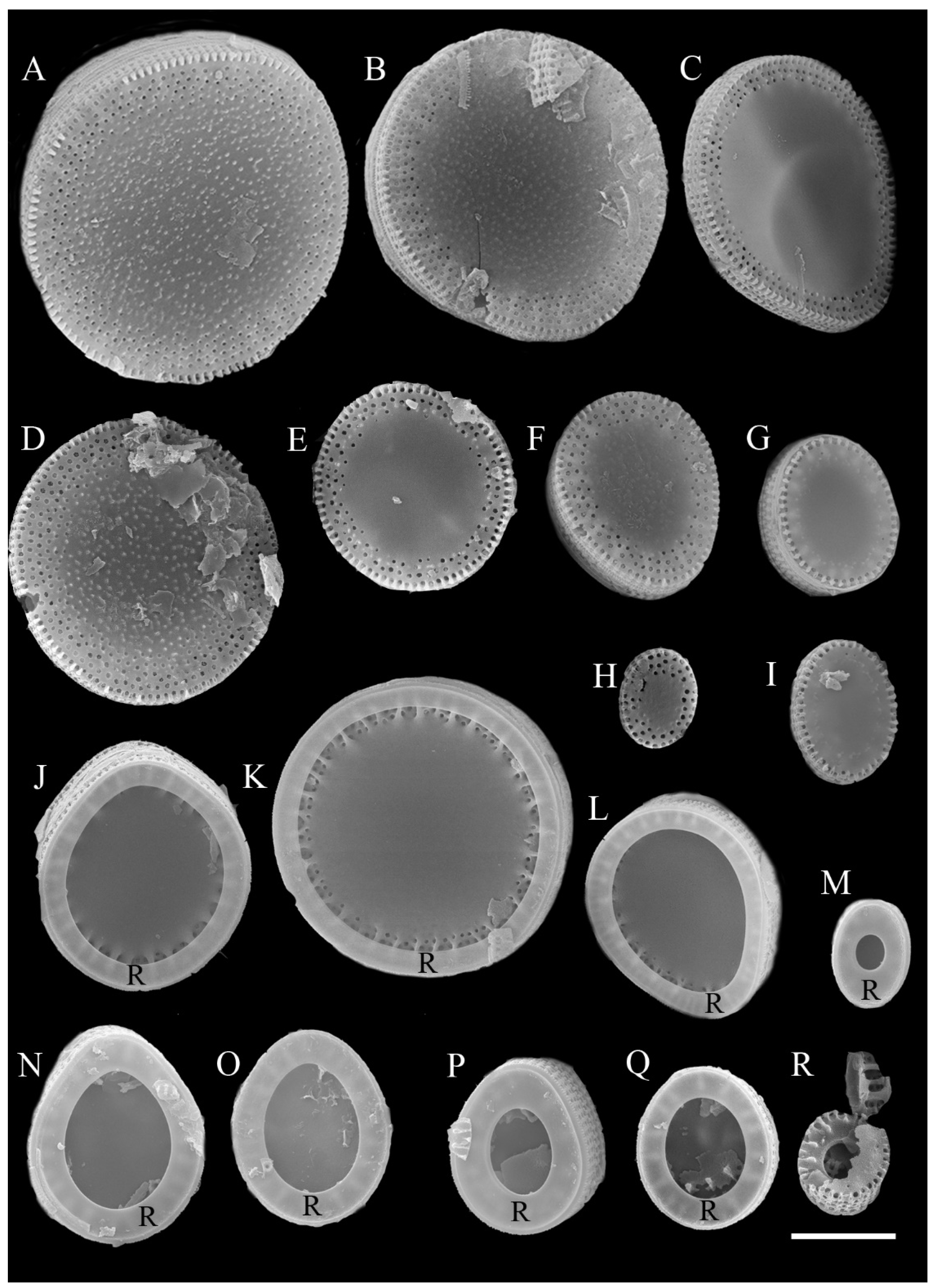
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hassan, A.; Usoltseva, M.V.; Rasskazov, S.V.; Chuvashova, I.S.; Titova, L.A. First study of fossil diatom flora from Middle Miocene-Lower Pliocene lacustrine sediments in Barguzin Valley, Baikal Rift Zone. Quart. Int. 2019, 524, 24–30. [Google Scholar] [CrossRef]
- Usoltseva, M.V.; Titova, L.A.; Hassan, A.; Chuvashova, I.S.; Rasskazov, S.V. Centric diatoms from paleolakes of the Baikal Rift Zone, Russia. Issues Mod. Algol. 2019, 2, 279–284. (In Russian) [Google Scholar] [CrossRef]
- Usoltseva, M.V.; Hassan, A.; Rodionova, E.V.; Titova, L.A.; Chuvashova, I.S.; Rasskazov, S.V. The first finding of diatoms from the Early Miocene lacustrine deposits of the Barguzin Valley (Baikal Rift Zone). Limnol. Freshw. Biol. 2020, 4, 752–754. [Google Scholar] [CrossRef]
- Moisseeva, A.I.; Nevretdinova, T.L. The new family and genus of fresh water diatom algae (Bacillariophyta). Bot. Zhurnal 1990, 75, 539–544. (In Russian) [Google Scholar]
- Glezer, S.I.; Makarova, I.V.; Moisseeva, A.I.; Nikolaev, V.A. The Diatoms of the USSR Fossil and Recent; Nauka St.-Petersburg Branch: St. Petersburg, Russia, 1992; Volume 2, Fasc. 2; p. 126. (In Russian) [Google Scholar]
- Kuz’min, M.I.; Khursevich, G.K.; Prokopenko, A.A.; Sedenya, S.A.; Karabanov, E.B. Late Cenozoic Centric Diatoms of Lake Baikal; Academic Publishing House Geo: Novosibirsk, Russia, 2009; p. 370. (In Russian) [Google Scholar]
- Hassan, A.I.; Rasskazov, S.V.; Chuvashova, I.S.; Reshetova, S.A.; Rubcova, M.N.; Yasnygina, T.A.; Budaev, R.C.; Al’Hamud, A.; Titova, L.A.; Rodionova, E.V.; et al. Cenozoic sediments from the Selenga-Vitim Downwarp and Baikal Rift Zone spatially combined in the Uro section. Litosfera 2022, 6. in press (In Russian) [Google Scholar]
- Grachev, M.A.; Vorobyova, S.S.; Likhoshway, Y.V.; Goldberg, E.L.; Ziborova, G.A.; Levina, O.V.; Khlystov, O.M. A high resolution diatom record of the palaeoclimates of East Siberia for the last 2.5 my from Lake Baikal. Quat. Sci. Rev. 1998, 17, 1101–1106. [Google Scholar] [CrossRef]
- Hunter, J.D. Matplotlib: A 2D graphics environment. Comput. Sci. Eng. 2007, 9, 90–95. [Google Scholar] [CrossRef]
- Jouse, A.P. The Bacillariophyta, Class Centricae. In Fundamentals of Palaeontology, A Reference Book for the Palaeontologists and Geologists of the USSR; Orlov, U.A., Ed.; Academy of Sciences of the USSR: Moscow, Russia, 1963; pp. 55–121. (In Russian) [Google Scholar]
- Jouse, A.P. Diatomeen in Seesedimenten. Arch. Fur Hydrobiol. Beihandlungen Ergeb. Limnol. 1966, 4, 1–32. (In Russian) [Google Scholar]
- Moiseeva, A.I. Revision of the systematic position of freshwater species of the genus Melosira Ag. s. l. (Bacillariophyta). In Sbornik Nauchnykh Trudov. Aktual’nye Voprosy Sovremennoi Paleoalgologii; Sokolov, B.S., Ed.; Naukova Dumka: Kiev, Ukraine, 1986; pp. 70–73. (In Russian) [Google Scholar]
- Kozyrenko, T.; Strelnikova, N.; Khursevich, G.; Tsoy, I.; Yakovschikova, T.; Mukhina, V.; Olshtynskaja, A.; Semina, G. The Diatoms of Russia and Adjacent Countries Fossil and Recent II; Strelnikova, N., Tsoy, I., Eds.; St. Petersburg University Press: St. Petersburg, Russia, 2008; p. 171. (In Russian) [Google Scholar]
- Usoltseva, M.V. Morphological variability of Alveolophora jouseana (Bacillariophyta) from type material. Phytotaxa 2022, 533, 194–204. [Google Scholar] [CrossRef]
- Khursevich, G.K.; Fedenya, S.A. Morphology of new fossil species of Concentrodiscus and Alveolophora (Bacillariophyta) from bottom sediment of Lake Baikal. Int. J. Algae 2006, 8, 78–90. [Google Scholar] [CrossRef]
- Titova, L.A.; Hassan, A.; Mikhailov, I.S.; Rodionova, E.V.; Rasskazov, S.V.; Usoltseva, M.V. Diversity and Ecology of Diatoms in Pliocene Deposits of the Tunka Valley (Baikal Rift Zone). Diversity 2021, 13, 479. [Google Scholar] [CrossRef]
- Servant-Vildary, S.; Paicheler, J.C.; Semelin, B. Miocene lacustrine Diatom from Turkey, Proceedings of the 9th International Diatom Symposium, Bristol, UK, 24–30 August 1986; Round, F.E., Ed.; Biopress, Ltd.: Bristol, UK; Koeltz, Koenigstein: Bristol, UK, 1986; pp. 165–180. [Google Scholar]
- Krammer, K. Die Cymbelloiden Diatomeen, Teil 2. Encyonema Part. Encyonopsis und Cymbellopsis, Bibliotheca Diamotologica Bd. 37; Cramer, J., Ed.; Schweizerbart Science Publishers: Berlin, Germany; Stuttgart, Germany, 1997; p. 469. [Google Scholar]
- Usoltseva, M.V.; Tsoy, I.B. Elliptical species of the freshwater genus Aulacoseira in Miocene sediments from Yamato Rise (Sea of Japan). Diatom Res. 2010, 25, 397–415. [Google Scholar] [CrossRef]
- Tanaka, H.; Nagumo., T.; Akiba, F. Aulacoseira hachiyaensis sp. nov., A new Early Miocene freshwater fossil diatom from Hachiya Formation, Japan. In Proceedings of the 19th International Diatom Symposium, Irkutsk, Russia, 28 August–3 September 2006; Biopress: Bristol, UK, 2008; pp. 115–123. [Google Scholar]
- Tanaka, H.; Nagumo, T. Aulacoseira iwakiensis Sp. Nov., A New Elliptical Aulacoseira Species, From an Early Miocene Sediment, Japan. Diatom 2011, 27, 1–8. [Google Scholar]
- Iwahashi, Y. Freshwater Centricae in Japan (Iv). J. Jpn. Bot. 1935, 11, 768–771. (In Japanese) [Google Scholar]
- Titova, L.A.; Hassan, A.; Usoltseva, M. Aulacoseira capitalina sp. nov. (Bacillariophyta) from the Middle Miocene sediments of the Barguzin Valley, Baikal Rift Zone (Russia). Acta Biol. Sibirica 2022, 8, 571–582. (In Russian) [Google Scholar] [CrossRef]
- Schrader, H.J. Quaternary through Neogene history of the Black Sea, deduced from the paleoecology of diatoms, silicoflagellates, ebridians, and chrysomonads. In Initial Reports of the Deep-Sea Drilling Project; Ross, D.A., Neprochnov, Y.P., Eds.; U.S. Government Printing Office: Washington, DC, USA, 1978; Volume 42, pp. 789–901. [Google Scholar]
- Jouse, A.P.; Mukhina, V.V. Diatoms units and the paleogeography of the Black Sea in the late Cenozoic. DSDP. In Initial Reports of the Deep-Sea Drilling Project; Ross, D.A., Neprochnov, Y.P., Eds.; U.S. Government Printing Office: Washington, DC, USA, 1978; pp. 903–950. [Google Scholar]
- Connolly, J.A.; Oliver, M.J.; Beaulieu, J.M.; Knight, C.A.; Tomanek, L.; Moline, M.A. Correlated evolution of genome size and cell volume in diatoms (Bacillariophyceae). J. Phycol. 2008, 44, 124–131. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Parks, M.B.; Nakov, T.; Ruck, E.C.; Wickett, N.J.; Alverson, A.J. Phylogenomics reveals an extensive history of genome duplication in diatoms (Bacillariophyta). Am. J. Bot. 2018, 105, 330–347. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koester, J.A.; Swalwell, J.E.; Von Dassow, P.; Armbrust, E. Genome size differentiates co-occurring populations of the planktonic diatom Ditylum brightwellii (Bacillariophyta). BMC Evol. Biol. 2010, 10, 1. [Google Scholar] [CrossRef] [Green Version]
- Round, F.E.; Crawford, R.M.; Mann, D.G. The Diatoms: Biology and Morphology of the Genera; Cambridge University Press: Cambridge, UK, 1990; p. 747. [Google Scholar]
- Mann, D.G.; Chepurnov, V.A.; Droop, S.J.M. Sexuality, incompatibility, size variation, and preferential polyandry in natural populations and clones of Sellaphora pupula (Bacillariophyceae). J. Phycol. 1999, 35, 152–170. [Google Scholar] [CrossRef]
- Winder, M.; Reuter, J.E.; Schlado, S.G. Lake warming favours small-sized planktonic diatom species. Proc. R. Soc. 2008, 276, 427–435. [Google Scholar] [CrossRef]
- Wimpenny, R. The Size of Diatoms. I. The Diameter Variation of Rhizosolenia Styliformis Brightw. and R. Alata Brightw. in Particular and of Pelagic Marine Diatoms in General. J. Mar. Biol. Assoc. U. K. 1936, 21, 29–60. [Google Scholar] [CrossRef] [Green Version]
- Pinto, R.; Mortágua, A.; Almeida, S.F.P.; Serra, S.; Feio, M.J. Diatom size plasticity at regional and global scales. Limnetica 2020, 39, 387–403. [Google Scholar] [CrossRef]
- Berven, K.A. The genetic basis of altitudinal variation in the wood frog Rana sylvatica. I. An experimental analysis of life history traits. Evolution 1982, 36, 962–983. [Google Scholar] [CrossRef]
- Zhong, M.; Yu, X.; Liao, W. A Review for Life-history Traits Variation in Frogs Especially for Anurans in China. Asian Herpatol. Res. 2018, 9, 165–174. [Google Scholar] [CrossRef]
- Irie, T.; Fischer, K. Ectotherms with a calcareous exoskeleton follow the tempera_ture-size rule-evidence from field survey. Mar. Ecol. Prog. Ser. 2009, 385, 33–37. [Google Scholar] [CrossRef]
- Daufresne, M.; Lengfellner, K.; Sommer, U. Global warming benefits the small in aquatic ecosystems. Proc. Natl. Acad. Sci. USA 2009, 106, 12788–12793. [Google Scholar] [CrossRef] [Green Version]
- Desai, A.S.; Singh, R.K. The effects of water temperature and ration size on growth and body composition of fry of common carp, Cyprinus carpio. J. Therm. Biol. 2009, 34, 276–280. [Google Scholar] [CrossRef]
- Sahin, T. Effect of water temperature on growth of hatchery reared Black Sea turbot, Scophthalmus maximus (Linnaeus, 1758). Turk. J. Zool. 2001, 25, 183–186. [Google Scholar]
- Stillwell, R.C.; Fox, C.W. Geographic variation in body size, sexual size dimorphism and fitness components of a seed beetle: Local adaptation versus phenotypic plasticity. Oikos 2009, 118, 703–712. [Google Scholar] [CrossRef]
- Brehm, G.; Zeuss, D.; Colwell, R.K. Moth body size increases with elevation along a complete tropical elevational gradient for two hyperdiverse clades. Ecography 2018, 42, 632–642. [Google Scholar] [CrossRef] [Green Version]
- Bizer, J.R. Growth rates and size at metamorphosis of high elevation populations of Ambystoma tigrinum. Oecologia 1978, 34, 175–184. [Google Scholar] [CrossRef] [PubMed]
- Schäuble, C.S. Variation in body size and sexual dimorphism across geographical and environmental space in the frogs Limnodynastes tasmaniensis and L. peronei. Biol. J. Linn. Soc. 2004, 82, 39–56. [Google Scholar] [CrossRef]













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Taxon | A. jouseana | A. baicalensis | A. antiqua | A. antiqua | A. antiqua | A. antiqua | A. antiqua |
|---|---|---|---|---|---|---|---|
| Valve form | round | round | triangular–oval and triangular with widely rounded corners | triangular and triangular with widely rounded corners | ovoid | oval | round |
| Height of valves | 2.0–7.0 | 1.0–5.0 | 4.0–8.0 | 2.3–3.0 | 2.3–2.7 | 3.2–3.8 | 2.5–3.0 |
| Diameter of valve (μm) | 8.0–31.0 | 6.0–25.0 | – | – | 9.0–27.0 a.a 6.5–24.0 t.a | 5.0–22.0 a.a 4.0–16.0 t.a | 13.0–33.0 |
| Length of one side of the valve | – | – | 8.0–40.0 | 8.0–53.0 | – | – | – |
| Number of areolar rows in 10 μm | 8–12 straight rows, each 2–4 rows of areolae alternate with costae | 10–14 straight rows, each 2–4 rows of areolae alternate with costae | 7–9 straight rows, each 2–4 rows of areolae alternate with costae | 11–15 straight rows, each 2–4 rows of areolae alternate with costae | 12–14 straight rows, each 2–4 rows of areolae alternate with costae | 12–14 | 12–14 straight rows, each 1–3 rows of areolae alternate with costae |
| Number of areolae in a row | 3–5 | 2–5 | 2–4 | 2–4 | 3 | 2–4 | |
| Areolae on the valve face | over the entire surface, along the periphery in 1–3 areolae rows or completely absent. Sometimes has granules at the center or one ring of granules at the periphery. | areolae in the submarginal zone of the valve face in short radial rows alternating with rib-like thickenings | without areolae or only on the periphery. Sometimes has granules at the center. | without areolae or only on the periphery 1–4 rows. Valve surface smooth or with granules. Sometimes, one ring of granules at the periphery. | without areolae or only on the periphery 1–4 rows. Valve surface smooth or with granules | without areolae or only on the periphery 1–4 rows. Valve surface smooth | without areolae or only on the peripheral 1–4 rows. Valve surface smooth or with granules. Sometimes, one ring of granules at the periphery |
| Linking spines | short, spathulate–widened | spathulate | small | spathulate | spathulate | spathulate | spathulate |
| Separating spines | pointed | – | – | elongated petaloid | - | - | – |
| Ringleist | narrow | narrow | narrow–wide | narrow–wide | narrow–wide | narrow–wide | narrow–wide |
| Shape and diameter (μm) of ringleist aperture | big round or medium | big round or medium | triangular or oval | triangular with widely rounded corners 5.0–6.0 a.a 4.7 t.a | ovoid or oval, 2.3–16.0 a.a 2.0–12.0 t.a | oval, 3.5–8.0 a.a 2.7–5.0 t.a | round, 6.0–9.0 |
| Rimoportulae | small, sessile, located at the base of the costae or pseudoalveolae | sessile, located at the base of pseudoalveolae | – | sessile, located at the base of alveolae, every 2–4 costae | sessile, located at the base of alveolae, every 2–4 costae | sessile, located at the base of alveolae, every 2–4 costae | – |
| Alveolar character | pseudoalveolae and micro-pseudoalveolae | pseudoalveolae | – | alveolae | alveolae | alveolae | alveolae |
| Number of costae in 10 µm | 3–10 | 4–8 (12) | – | 3–6 | 3–8 | 4–6 | 3–8 |
| Location | Far East, Primorye, Lake Tony | Lake Baikal, | Transbaikalia, Vitim Plateau | Barguzin Valley, core 517 | Barguzin Valley, core 517 | Barguzin Valley, core 517 | Barguzin Valley, core 517 |
| Age | Miocene | Late Miocene | Early Miocene | Early Miocene | Early Miocene | Early Miocene | Early Miocene |
| References | [14] | [15] | [4] | present data | present data | present data | present data |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Usoltseva, M.; Titova, L.; Hassan, A.; Rasskazov, S.; Morozov, A. Morphological Variability of Alveolophora antiqua from a Freshwater Early Miocene Paleolake in the Barguzin Valley (Baikal Rift Zone). Diversity 2022, 14, 1075. https://doi.org/10.3390/d14121075
Usoltseva M, Titova L, Hassan A, Rasskazov S, Morozov A. Morphological Variability of Alveolophora antiqua from a Freshwater Early Miocene Paleolake in the Barguzin Valley (Baikal Rift Zone). Diversity. 2022; 14(12):1075. https://doi.org/10.3390/d14121075
Chicago/Turabian StyleUsoltseva, Marina, Lyubov’ Titova, Abdulmonem Hassan, Sergei Rasskazov, and Alexey Morozov. 2022. "Morphological Variability of Alveolophora antiqua from a Freshwater Early Miocene Paleolake in the Barguzin Valley (Baikal Rift Zone)" Diversity 14, no. 12: 1075. https://doi.org/10.3390/d14121075