
Disentangling the Diversity of the Labeobarbus Taxa (Cypriniformes: Cyprinidae) from the Epulu Basin (DR Congo, Africa)

Abstract
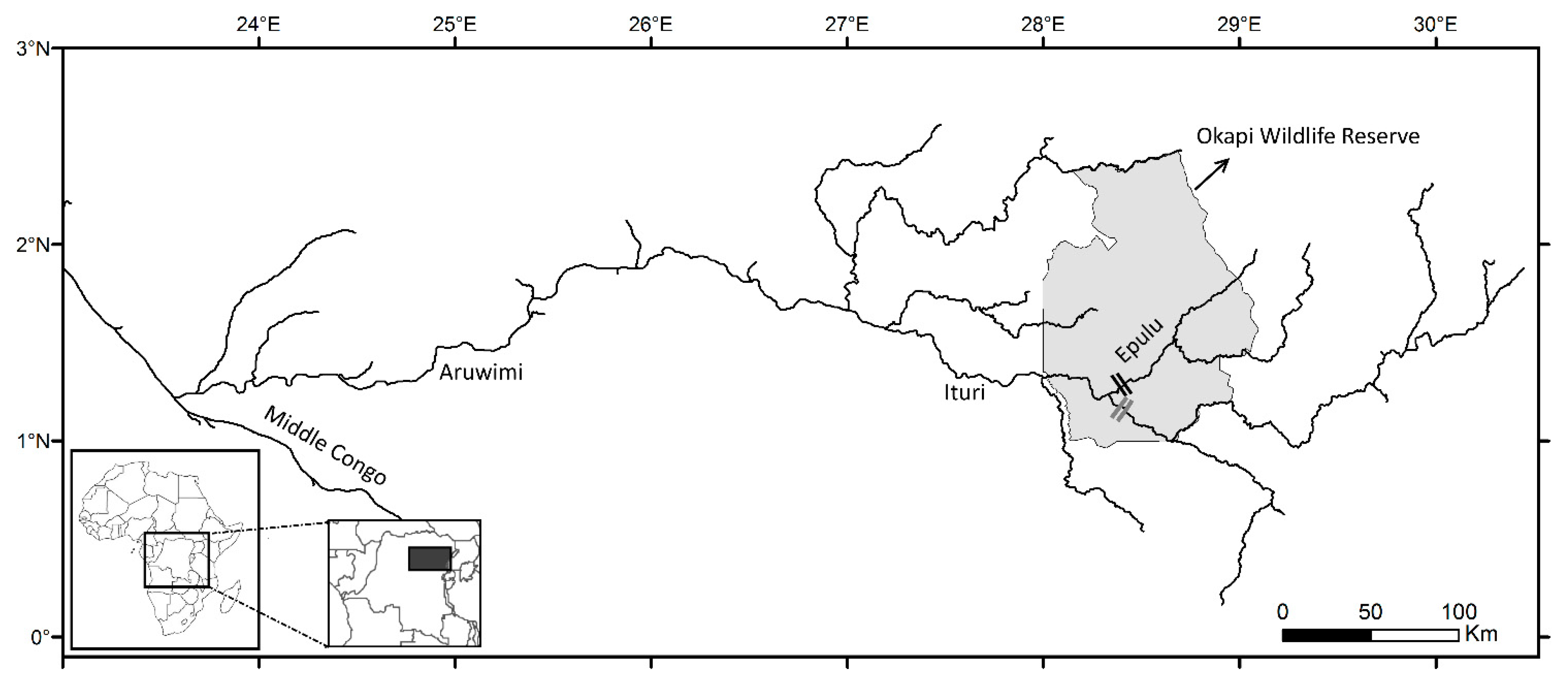
:1. Introduction
2. Materials and Methods
2.1. Morphology
2.2. Genetics
2.3. Abbreviations
3. Results
3.1. The Epulu Specimens: A Phenotypic Classification
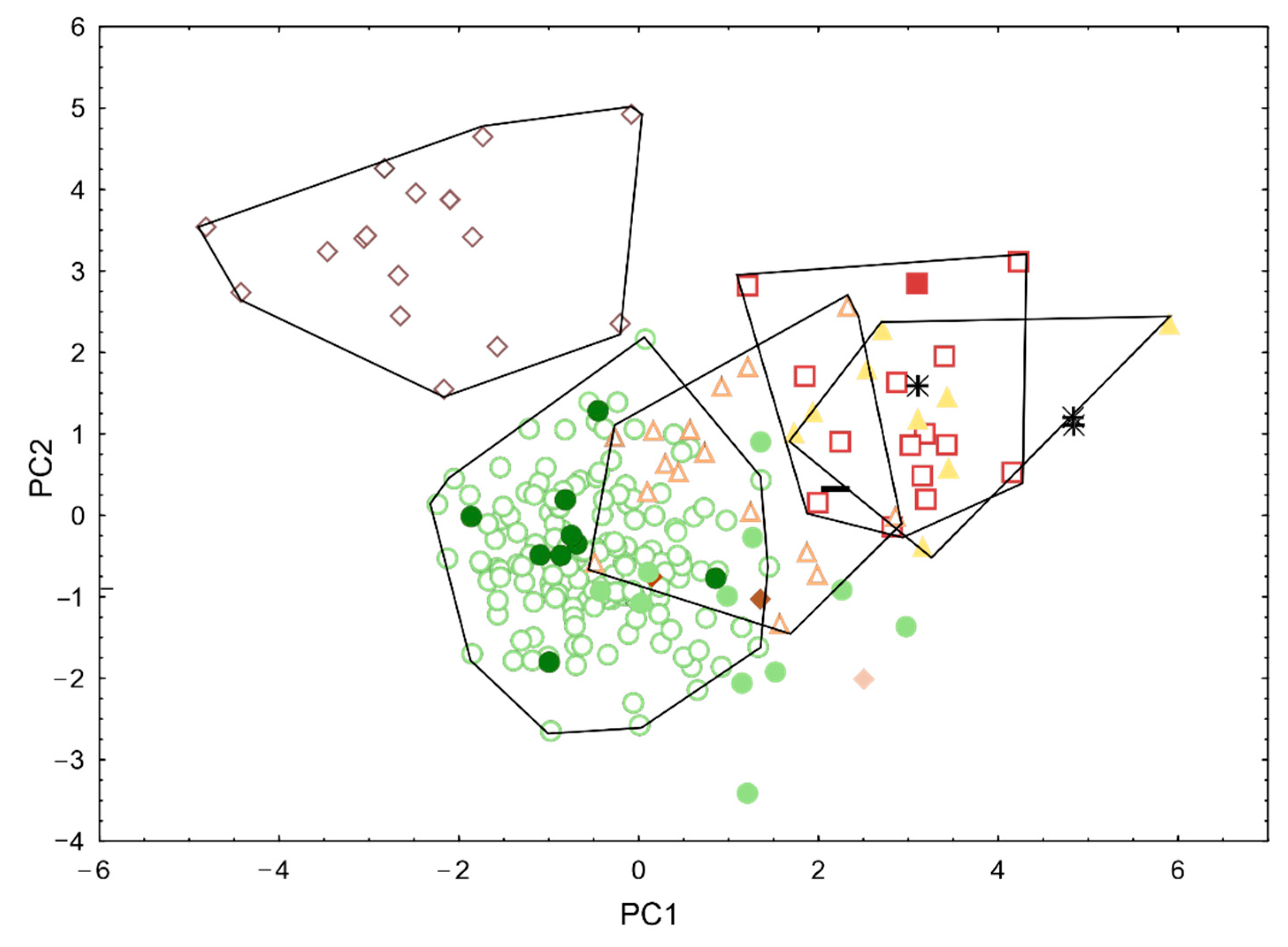
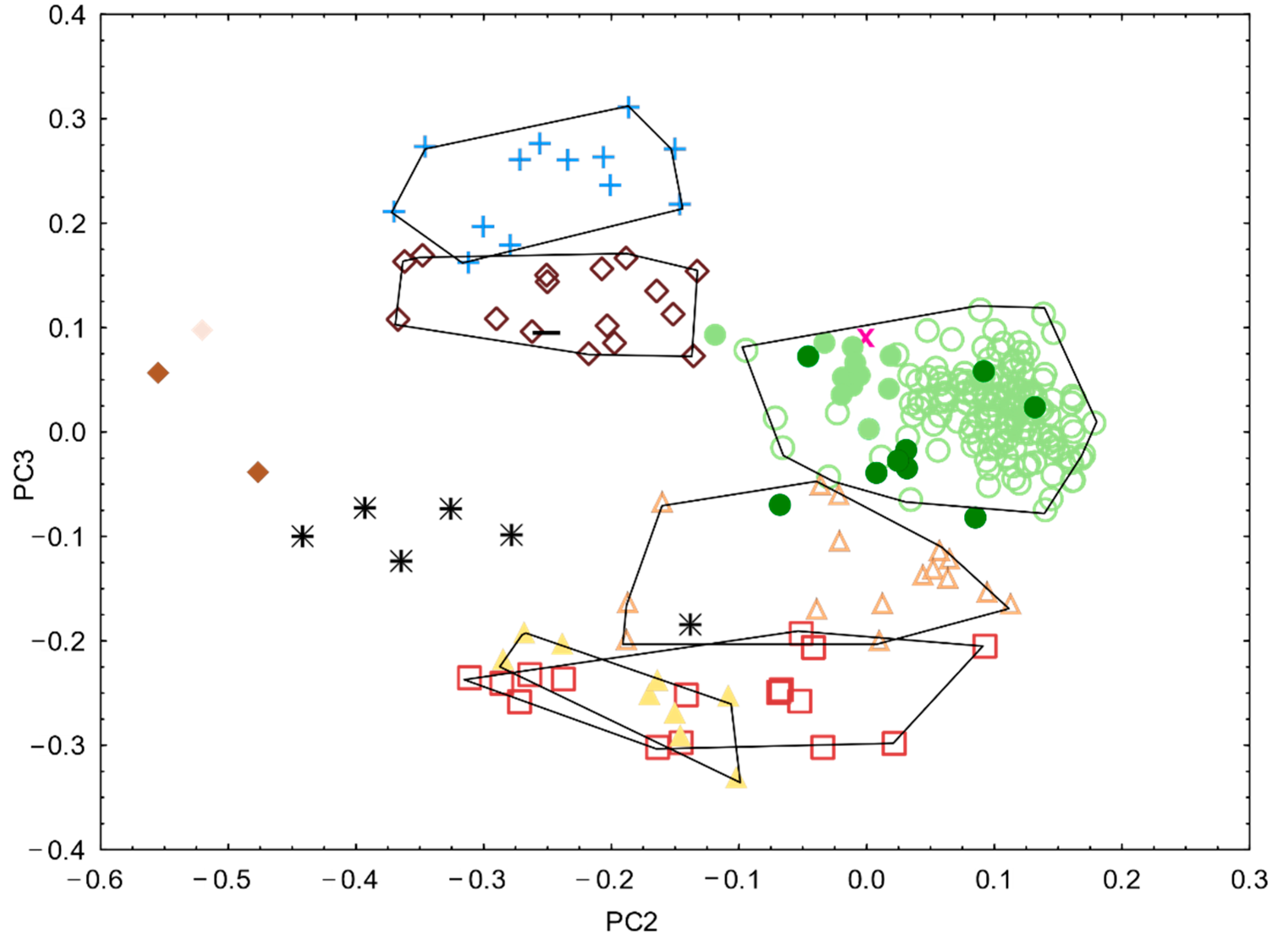
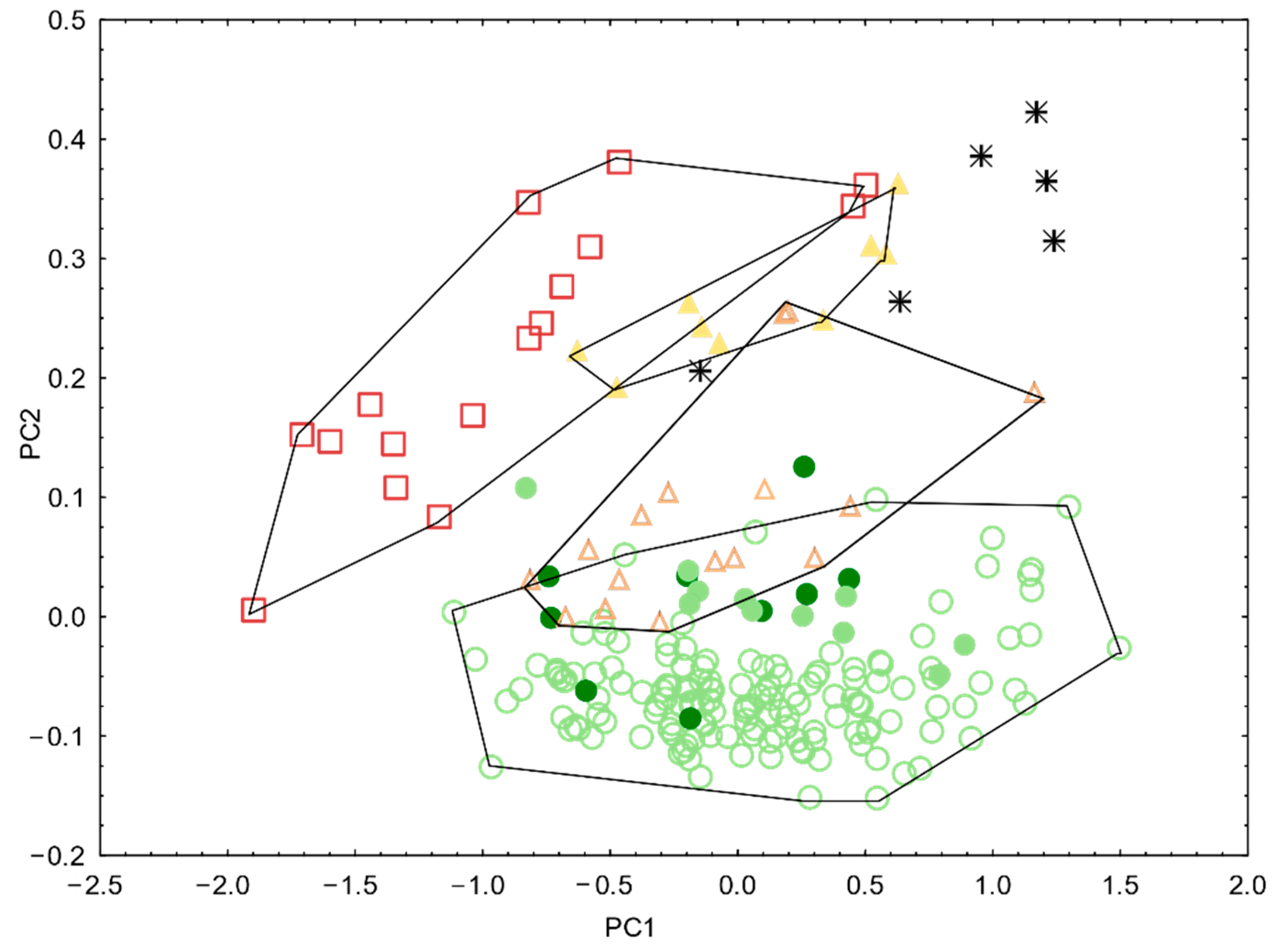
3.2. Morphological Analyses
3.2.1. Meristics
3.2.2. Measurements
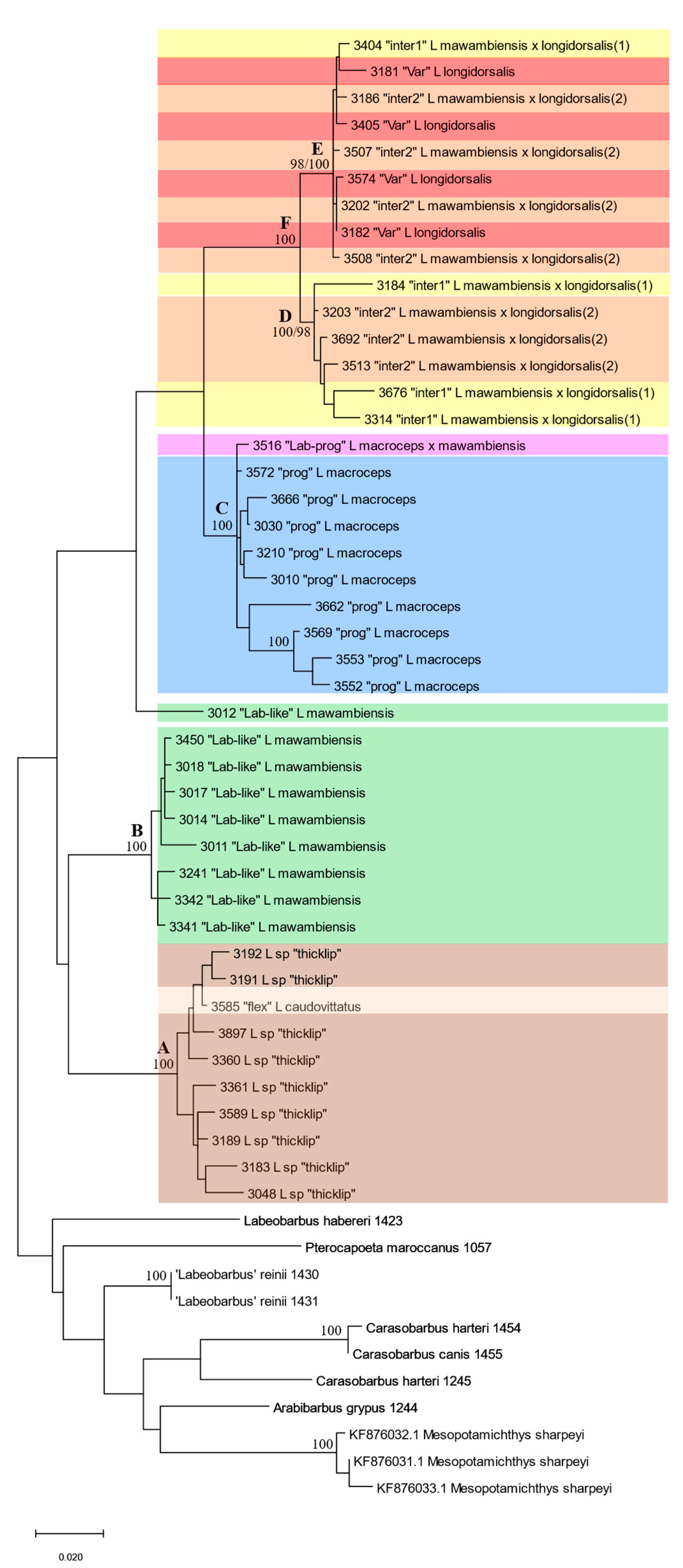
3.3. Genetics
3.4. Integrative Synthesis
4. Discussion
4.1. Which Labeobarbus Species Are Present in the Epulu Basin?
4.2. The Labeobarbus mawambiensis/longidorsalis Hybrid Complex
4.3. Additional Nomenclatorial Decisions
4.4. Hybridisation: A Widespread and Variable Phenomenon in Labeobarbus
4.5. Identification Key to the Labeobarbus Species and Possible Hybrids of the Epulu Basin
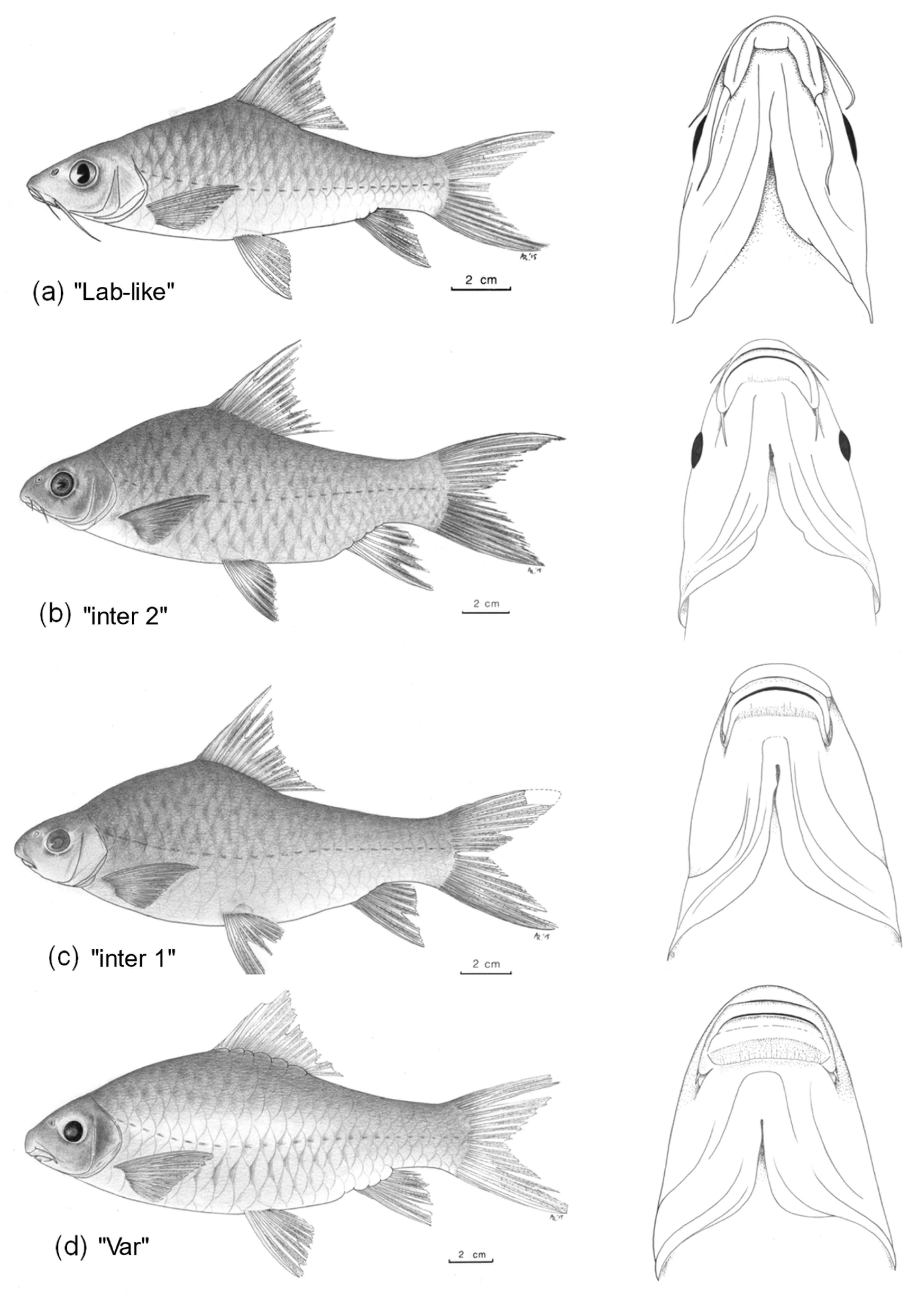
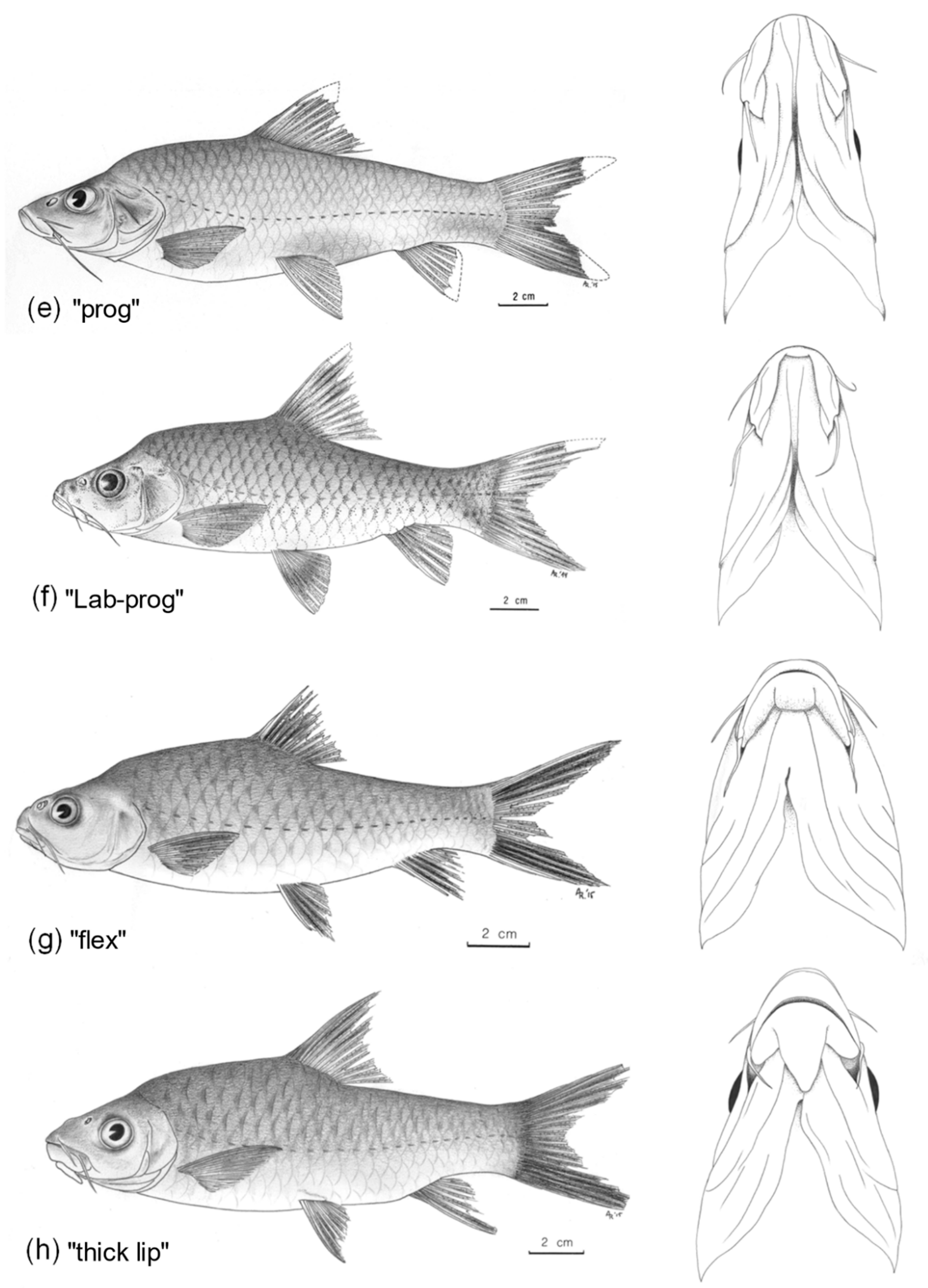
| (1) Well-defined keratinised cutting edge on lower jaw (Figure 2d) | L. longidorsalis |
| No keratinised cutting edge on lower jaw (Figure 2a–c,e–h) | 2 |
| (2) Lower jaw slightly to clearly prognathous (Figure 2e,f); 29–36 (median: 33) lateral line scales; 9–14 gill rakers on first gill arch | 3 |
| Mouth inferior; 21–29 (25) lateral line scales; 14–23 rakers on first gill arch | 4 |
| (3) 31–36 lateral line scales; 9–11 gill rakers on first gill arch; last unbranched dorsal fin ray flexible (38.0–61.6% of the dorsal fin height unsegmented) (Figure 2e) | L. macroceps |
| 29 lateral line scales; 14 gill rakers on first gill arch; last unbranched dorsal fin ray a well-ossified spine (66.8% of the dorsal fin height unsegmented; strongly ossified) (Figure 2f) | L. macroceps x mawambiensis hybrid |
| (4) 19–23 (median: 21) gill rakers on first gill arch; lower lip with a large, posteriorly detached, median lobe (Figure 2h) | L. sp. ‘thick lip’ |
| 14–19 (17) gill rakers on first arch; lower lip with or without a mental lobe; if present, mostly posteriorly attached (Figure 2a–c,g) | 5 |
| (5)One pair of short posterior barbels (Figure 2c) | putative L. longidorsalis x mawambiensis hybrid with one pair of barbels |
| Two pairs of barbels | 6 |
| (6) Barbels short; anterior barbels 7.8–12.6%HL and posterior barbels 5.1–17.1%HL; 10–12 (median: 11) branched dorsal fin rays; no mental lobe (Figure 2b) | putative L. longidorsalis x mawambiensis hybrid with two pairs of barbels |
| Barbels long; anterior barbels 15.8–40.7%HL, posterior barbels 19.6–42.9%HL; 8–11 (10) branched dorsal fin rays; mental lobe present, mostly posteriorly attached, sometimes free (Figure 2a,g) | 7 |
| (7) Last unbranched dorsal fin ray flexible (weakly ossified proximal part: 39.9%); dark grey to black band along the distal part of upper and lower caudal-fin lobes (Figure 2g) | L. caudovittatus |
| Last unbranched dorsal fin ray a well ossified spine (strongly ossified proximal part: 57.6–98.6%); upper and lower caudal-fin lobes uniform yellowish to grey (Figure 2a) | L. mawambiensis |
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A. Specimens Examined
- Type specimens
- Specimens from the Epulu River
References
- Yang, L.; Sado, T.; Hirt, M.V.; Pasco–Viel, E.; Arunachalam, M.; Li, J.; Wang, X.; Freyhof, J.; Saitoh, K.; Simons, A.M.; et al. Phylogeny and polyploidy: Resolving the classification of cyprinine fishes (Teleostei: Cypriniformes). Mol. Phylogenetics Evol. 2015, 85, 97–116. [Google Scholar] [CrossRef]
- Tsigenopoulos, C.S.; Kasapidis, P.; Berrebi, P. Phylogenetic relationships of hexaploid large–sized barbs (genus Labeobarbus, Cyprinidae) based on mtDNA data. Mol. Phylogenetics Evol. 2010, 56, 851–856. [Google Scholar] [CrossRef]
- Berrebi, P.; Chenuil, A.; Kotlik, P.; Machordom, A.; Tsigenopoulos, C.A. Disentangling the evolutionary history of the genus Barbus sensu latu, a twenty years adventure. Mus. Nac. De Hist. Nat. E Da Cienc. Lisb. 2014, 2013, 29–55. [Google Scholar]
- Vreven, E.; Musschoot, T.; Snoeks, J.; Schliewen, U.K. The African hexaploid barbines (Cypriniformes: Cyprinidae): Review of a tumultuous history. Zool. J. Linn. Soc. 2016, 177, 231–305. [Google Scholar] [CrossRef] [Green Version]
- Skelton, P.H. A Complete Guide to the Freshwater Fishes of Southern Africa, 2nd ed.; Struik: Cape Town, South Africa, 2001. [Google Scholar]
- Boulenger, G.A. Catalogue of the Fresh–Water Fishes of Africa in the British Museum (Natural History); British Museum (Natural History): London, UK, 1911; Volume 2. [Google Scholar]
- Banister, K.E. On the cyprinid fish Barbus alluaudi Pellegrin: A possible intergeneric hybrid from Africa. Studies on African Cyprinidae. Part I. Bull. Br. Mus. (Nat. Hist.) Zool. 1972, 24, 261–290. [Google Scholar]
- Banister, K.E. A possible intergeneric cyprinid hybrid from Lake Tanganyika. Bull. Br. Mus. (Nat. Hist.) Zool. 1976, 30, 171–189. [Google Scholar]
- Vreven, E.; Musschoot, T.; Decru, E.; Wamuini Lunkayilakio, S.; Obiero, K.; Cerwenka, A.F.; Schliewen, U.K. The complex origins of mouth polymorphism in the Labeobarbus (Cypriniformes: Cyprinidae) of the Inkisi River basin (Lower Congo, DRC, Africa): Insights from an integrative approach. Zool. J. Linn. Soc. 2019, 186, 414–482. [Google Scholar] [CrossRef]
- Snoeks, J.; Harrison, I.J.; Stiassny, M.L.J. The status and distribution of freshwater fishes. In The Diversity of Life in African Freshwaters: Underwater, under Threat. An Analysis of the Status and Distribution of Freshwater Species throughout Mainland Africa; Darwall, W., Smith, K., Allen, D., Holland, R., Harrison, I., Brooks, E., Eds.; IUCN: Gland, UK; Cambrigde, UK, 2011; pp. 42–91. [Google Scholar]
- Thieme, M.L.; Abell, R.; Stiassny, M.L.J.; Skelton, P.; Lehner, B.; Teugels, G.G.; Dinerstein, E.; Kamdem Toham, A.; Burgess, N.; Olson, D. Freshwater Ecoregions of Africa and Madagascar: A Conservation Assessment; Island Press: Washington, DC, USA, 2005. [Google Scholar]
- Reid, R.L. The River Aruwimi. Geogr. J. 1911, 38, 29–34. [Google Scholar] [CrossRef]
- De Vos, L. Données préliminaires sur l’Ichthyofaune de l’Epulu dans le futur Parc National de la fôret d’Ituri (Zaire). 1990; unpublished. [Google Scholar]
- Decru, E.; Vreven, E.; Danadu, C.; Walanga, A.; Mambo, T.; Snoeks, J. The ichthyofauna of the Itimbiri, Aruwimi and Lindi/Tshopo Rivers (Congo basin): Diversity and distribution patterns. Acta Ichthyol. Piscat. 2017, 47, 225–247. [Google Scholar] [CrossRef] [Green Version]
- Walanga, A.; Danadu, C.; Mambo, T.; Vreven, E. Rapport scientifique de l’expédition «Réserve de Faune à Okapis» (RFO) 2009 (République Démocratique du Congo). 2009; unpublished. [Google Scholar]
- UNESCO. World Heritage Centre—List of World Heritage in Danger. Available online: https://whc.unesco.org/en/danger/ (accessed on 15 February 2019).
- Banister, K.E. A revision of the large Barbus (Pisces, Cyprinidae) of the East and Central Africa. Studies on African Cyprinidae Part II. Bull. Br. Mus. Nat. Hist. Zool. 1973, 26, 1–148. [Google Scholar]
- Trewavas, E. The freshwater fishes of rivers Mungo and Meme and lakes Kotto, Mboandong and Soden, West Cameroon. Bull. Br. Mus. Nat. Hist. Zool. 1974, 26, 329–419. [Google Scholar] [CrossRef]
- de Quieroz, K. Species concepts and species delimitation. Syst. Biol. 2007, 56, 879–886. [Google Scholar] [CrossRef] [Green Version]
- Yeates, D.K.; Seago, A.; Nelson, L.; Cameron, S.L.; Joseph, L.E.O.; Trueman, J.W. Integrative taxonomy, or iterative taxonomy? Syst. Entomol. 2011, 36, 209–217. [Google Scholar] [CrossRef]
- Daget, J.; Gosse, J.P.; Thys van den Audenaerde, D.F.E. Check–List of the Freshwater Fishes of Africa; RMCA: Paris, France; ORSTOM: Brussels, Belgium, 1984; Volume 1, ISBN 9782871770022. [Google Scholar]
- Kullander, S.O.; Roberts, T.R. Out of Lake Tanganyika: Endemic lake fishes inhabit rapids of the Lukuga River. Ichthyol. Explor. Freshwat. 2012, 22, 355–376. [Google Scholar]
- Walanga, A.; Decru, E.; Vreven, E. Labeobarbus caudovittatus (Boulenger, 1902) (Cypriniformes: Cyprinidae) from the Congo basin: A morphological re-evaluation of its putative intraspecific variation. Royal Museum for Central Africa, Tervuren, Belgium. 2022; manuscript in preparation. [Google Scholar]
- Bookstein, F.L.; Chernoff, B.; Elder, R.L.; Humphries, J.M.; Smith, G.R.; Strauss, R.E. Morphometrics in Evolutionary Biology: The Geometry of Size and Shape Change, with Examples from Fishes; Academy of Natural Sciences of Philadelphia: Philadelphia, PA, USA, 1985. [Google Scholar]
- Snoeks, J. The Cichlid Diversity of Lake Malawi/Nyasa/Niassa: Identification, Distribution and Taxonomy; Cichlid Press: El Paso, TX, USA, 2004. [Google Scholar]
- Rice, W.R. Analyzing tables of statistical tests. Evolution 1989, 43, 223–225. [Google Scholar] [CrossRef]
- Briolay, J.; Galtier, N.; Brito, R.M.; Bouvet, Y. Molecular phylogeny of cyprinidae inferred from cytochrome b DNA Sequences. Mol. Phylogenetics Evol. 1998, 9, 100–108. [Google Scholar] [CrossRef]
- Esposti, D.M.; De Vries, S.; Crimi, M.; Ghelli, A.; Patarnello, T.; Meyer, A. Mitochondrial cytochrome b: Evolution and structure of the protein. Biochim. Biophys. Acta (BBA)–Bioenerg. 1993, 1143, 243–271. [Google Scholar] [CrossRef] [Green Version]
- Durand, J.D.; Tsigenopoulos, C.S.; Ünlü, E.; Berrebi, P. Phylogeny and biogeography of the family Cyprinidae in the Middle East inferred from cytochrome b DNA—Evolutionary significance of this region. Mol. Phylogenetics Evol. 2002, 22, 91–100. [Google Scholar] [CrossRef]
- Schwarzer, J.; Swartz, E.R.; Vreven, E.; Snoeks, J.; Cotterill, F.P.D.; Misof, B.; Schliewen, U.K. Repeated trans–watershed hybridization among haplochromine cichlids (Cichlidae) was triggered by Neogene landscape evolution. Proc. R. Soc. B Biol. Sci. 2012, 279, 4389–4398. [Google Scholar] [CrossRef]
- Hall, T.A. Bioedit: A user–friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucleic Acids Symp. Ser. 2012, 41, 95–98. [Google Scholar]
- Sabaj Perez, M.H. Standard Symbolic Codes for Institutional Resource Collections in Herpetology and Ichthyology: An Online Reference; Version 5.0; American Society of Ichthyologists and Herpetologists: Washington, DC, USA, 2014; Available online: http://www.asih.org/ (accessed on 15 August 2020)Version 5.0.
- USBGN. Republic of the Congo (Léopoldville): Official standard names approved by the United States Board on Geographic Names; Gazetteer no. 80; Office of Geography, Department of the Interior: Washington, DC, USA, 1964. [Google Scholar]
- Padial, J.M.; Miralles, A.; De la Riva, I.; Vences, M. The integrative future of taxonomy. Front. Zool. 2010, 7, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Kisekelwa, T.; Snoeks, J.; Vreven, E. An annotated checklist of the fish fauna of the river systems draining the Kahuzi-Biega National Park (Upper Congo: Eastern DR Congo). J. Fish Biol. 2020, 96, 700–721. [Google Scholar] [CrossRef] [PubMed]
- Levin, B.A.; Casal-López, M.; Simonov, E.; Dgebuadze, Y.Y.; Mugue, N.S.; Tiunov, A.V.; Doadrio Golubtsov, A.S. Adaptive radiation of barbs of the genus Labeobarbus (Cyprinidae) in an East African river. Freshw. Biol. 2019, 64, 1721–1736. [Google Scholar] [CrossRef]
- Levin, B.A.; Simonov, E.; Dgebuadze, Y.Y.; Levina, M.; Golubtsov, A.S. In the rivers: Multiple adaptive radiations of cyprinid fishes (Labeobarbus) in Ethiopian Highlands. Sci. Rep. 2020, 10, 1–13. [Google Scholar]
- Roberts, T.R.; Khaironizam, M.Z. Trophic polymorphism in the Malaysian fish Neolissochilus soroides and other old world barbs (Teleostei, Cyprinidae). Nat. Hist. Bull. Siam Soc. 2008, 56, 25–53. [Google Scholar]
- Lévêque, C.; Daget, J. Cyprinidae. In Check-List of the Freshwater Fishes of Africa (CLOFFA); Daget, J., Gosse, J.P., Thys van den Audenaerde, D.F.E., Eds.; ORSTOM: Paris, France; MRAC: Tervuren, Belgium, 1984; Volume 1, pp. 217–342. [Google Scholar]
- Banister, K.E.; Clarke, M.A. A revision of the large Barbus (Pisces, Cyprinidae) of Lake Malawi with a reconstruction of the history of the southern African Rift Valley lakes. J. Nat. Hist. 1980, 14, 483–542. [Google Scholar] [CrossRef]
- de Graaf, M.; Nentwich, D.; Osse, J.M.W.; Sibbing, F.A. Lacustrine spawning: Is this a new reproductive strategy among ‘Large’ African Cyprinid fishes? J. Fish Biol. 2005, 66, 1214–1236. [Google Scholar] [CrossRef]
- Anteneh, W.A.; Getahun, A.; Dejen, F.A.; Sibbing, L.A.J.; Nagelkerke, M.; De Graaf, T.; Wudneh, J.; Vijverberg, A.; Palstra, P. Spawning migrations of the endemic Labeobarbus (Cyprinidae, Teleostei) species of Lake Tana, Ethiopia: Status and threats. J. Fish Biol. 2012, 81, 750–765. [Google Scholar] [CrossRef]
- Abdissa, B.; Getahun, A.; Dejen, E. Emulating the natural spawning conditions of Labeobarbus brevicephalus (Pisces: Cyprinidae) from Lake Tana, Ethiopia. Aquac. Rep. 2022, 23, 294–306. [Google Scholar] [CrossRef]
- Nagelkerke, L.A.J.; Sibbing, F.A. Reproductive segregation among the Barbus intermedius complex of Lake Tana, Ethiopia. An example of intralacustrine speciation? J. Fish Biol. 1996, 49, 1244–1266. [Google Scholar] [CrossRef]
- Zworykin, D.D.; Budaev, S.V.; Darkov, A.A.; Dzerzhinskii, K.F.; Lyovin, B.A.; Mina, M.V. Assessment of the role of chemoreception in the mate choice in barbs of the Barbus intermedius complex from Lake Tana, Ethiopia. J. Ichthyol. 2006, 46, 661–667. [Google Scholar] [CrossRef]
- Steindachner, F. Zur Fischfauna des Dscha, eines sekundären Nebenflusses des Congo im Bezirke Molundu des südlichen Kamerun. Anz. Der Akad. Der Wiss. Wien 1912, 49, 443–449. [Google Scholar]
- Steindachner, F. Zur Fischfauna des Dscha, eines sekundären Niebenflusses des Kongo, im Bezirke Molundu, Kamerun. Denkschr. Der Math.—Nat. Cl. Der Kais. Akad. Der Wiss. Wien 1914, 89, 1–64. [Google Scholar]
- Fricke, R.; Eschmeyer, W.N.; van der Laan, R. (Eds.) Eschmeyer’s Catalog of Fishes: Genera, Species, References; 2022. Available online: http://www.calacademy.org/scientists/catalog-of-fishes-classification/ (accessed on 3 February 2022).
- Banister, K.E. Three new species of Varicorhinus (Pisces, Cyprinidae) from Africa. Bulletin Br. Mus. (Nat. Hist.) Zool. 1984, 47, 273–282. [Google Scholar] [CrossRef]
- Jones, F.C.; Brown, C.; Braithwaite, V.A. Lack of assortative mating between incipient species of stickleback from a hybrid zone. Behaviour 2008, 145, 463–484. [Google Scholar]
- Svardal, H.; Salzburger, W.; Malinsky, M. Genetic variation and hybridization in evolutionary radiations of cichlid fishes. Annu. Rev. Anim. Biosci. 2021, 9, 55–79. [Google Scholar] [CrossRef]
- Keller, I.; Wagner, C.E.; Greuter, L.; Mwaiko, S.; Selz, O.M.; Sivasundar, A.; Wittwer, S.; Seehausen, O. Population genomic signatures of divergent adaptation, gene flow and hybrid speciation in the rapid radiation of Lake Victoria cichlid fishes. Mol. Ecol. 2013, 22, 2848–2863. [Google Scholar] [CrossRef]
- Wagner, C.E.; Keller, I.; Wittwer, S.; Selz, O.M.; Mwaiko, S.; Greuter, L.; Sivasundar, A.; Seehausen, O. Genome–wide RAD sequence data provide unprecedented resolution of species boundaries and relationships in the Lake Victoria cichlid adaptive radiation. Mol. Ecol. 2013, 22, 787–798. [Google Scholar] [CrossRef]
- Malinsky, M.; Svardal, H.; Tyers, A.M.; Miska, E.A.; Genner, M.J.; Turner, G.F.; Durbin, R. Whole-genome sequences of Malawi cichlids reveal multiple radiations interconnected by gene flow. Nat. Ecol. Evol. 2018, 2, 1940–1955. [Google Scholar] [CrossRef] [Green Version]
- Nagelkerke, L.A.J.; Leon-Kloosterziel, K.M.; Megens, H.J.; De Graaf, M.; Diekmann, O.E.; Sibbing, F.A. Shallow genetic divergence and species delineations in the endemic Labeobarbus species flock of Lake Tana, Ethiopia. J. Fish Biol. 2015, 87, 1191–1208. [Google Scholar] [CrossRef]
- Seehausen, O. Hybridization and adaptive radiation. Trends Ecol. Evol. 2004, 19, 198–207. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, R.C.; Bart, H.L., Jr.; Nyingi, W.D. Multi-locus phylogeny reveals instances of mitochondrial introgression and unrecognized diversity in Kenyan barbs (Cyprininae: Smiliogastrini). Mol. Phylogenetics Evol. 2017, 111, 35–43. [Google Scholar] [CrossRef] [Green Version]
- Borkenhagen, K.; Krupp, F. Taxonomic revision of the genus Carasobarbus Karaman, 1971 (Actinopterygii, Cyprinidae). ZooKeys 2013, 339, 1–53. [Google Scholar] [CrossRef] [PubMed]
- Harrison, R.G.; Larson, E.L. Hybridization, introgression, and the nature of species boundaries. J. Hered. 2014, 105, 795–809. [Google Scholar] [CrossRef] [PubMed]



 : syntypes of L. macrolepidotus, ♦: syntypes of L. caudovittatus.
: syntypes of L. macrolepidotus, ♦: syntypes of L. caudovittatus.
: syntypes of L. macrolepidotus, ♦: syntypes of L. caudovittatus.
: syntypes of L. macrolepidotus, ♦: syntypes of L. caudovittatus. : syntypes of L. macrolepidotus, ♦: syntypes of L. caudovittatus.
: syntypes of L. macrolepidotus, ♦: syntypes of L. caudovittatus.
: syntypes of L. macrolepidotus, ♦: syntypes of L. caudovittatus.
: syntypes of L. macrolepidotus, ♦: syntypes of L. caudovittatus. : syntypes of L. macrolepidotus.
: syntypes of L. macrolepidotus.
: syntypes of L. macrolepidotus.
: syntypes of L. macrolepidotus.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| (a) | ||||||
| Mouth Phenotypes | Mouth Position | Barbels | Dorsal Spine | LLS | Identification | |
| Epulu morphotypes | ||||||
| “Lab-like” | f or a | inf | 2p | spine | 21–28 | L. mawambiensis |
| “inter2” | n | inf | 2p | inter | 24–28 | L. maw x L. long (2p) |
| “inter1” | n | inf | 1p | inter | 24–27 | L. maw x L. long (1p) |
| “Var” | n & ce | inf | 1p | inter | 23–27 | L. longidorsalis |
| “prog” | n | prog | 2p | flexible | 31–36 | L. macroceps |
| “Lab-prog” | n | supra | 2p | spine | 29 | L. mac x maw |
| “flex” | a | inf | 2p | flexible | 24 | L. caudovittatus |
| “thick lip” | h & f | inf | 2p | flexible | 24–27 | L. sp. ‘thick lip’ |
| (b) | ||||||
| Mouth Phenotypes | Mouth Position | Barbels | Dorsal Spine | LLS | Status | |
| types | ||||||
| L. caudovittatus | a | inf | 2p | flexible | 26 | valid |
| L. fasolt | a | inf | 2p | flexible | 26 | =L. caudovittatus |
| L. humphrii | a | inf | 2p | spine | 24–28 | valid |
| L. iturii | a | inf | 2p | flexible | 29 | valid |
| L. longidorsalis | n & ce | inf | 1p | inter | 29 | valid |
| L. macroceps | n | inf | 2p | flexible | 32 | valid |
| L. macrolepidotus | n | inf | 1p | flexible | 25–27 | valid |
| L. mawambi | n | inf | 2p | flexible | 28 | =L. mirabilis |
| L. mawambiensis | f or a | inf | 2p | spine | 23–26 | valid |
| L. mirabilis | n | inf | 2p | inter | 31 | valid |
| PCI (20.7%) | PCII (12.7%) | |
|---|---|---|
| Total number of lateral line scales | 0.141 | 0.250 |
| Number of predorsal scales | −0.127 | −0.069 |
| Number of scales above the lateral line | 0.203 | 0.101 |
| Lateral line-pelvic scales | 0.232 | 0.050 |
| Lateral line–ventral midline scales | 0.032 | −0.125 |
| Number of dorsal-fin base scales | 0.272 | 0.057 |
| Number of anal-fin base scales | −0.013 | 0.029 |
| Number of branched dorsal fin rays | 0.439 | 0.249 |
| Number of branched pectoral fin rays | −0.227 | −0.278 |
| Number of branched pelvic fin rays | 0.108 | 0.212 |
| Number of scales between dorsal and caudal fin | −0.136 | 0.328 |
| Number of lateral line scales between anterior dorsal- and pelvic-fin base | 0.397 | 0.145 |
| Number of gill rakers on lower branch of first gill arch | −0.204 | 0.591 |
| Number of gill rakers on upper branch of first gill arch | −0.404 | −0.044 |
| Total number of gill rakers on the first gill arch | −0.397 | 0.485 |
| (a) | (b) | |||||
|---|---|---|---|---|---|---|
| PC1 (91.1%) | PC2 (3.8%) | PC3 (2.1%) | PC1 (93.1%) | PC2 (4.0%) | PC3 (0.9%) | |
| Standard length | −0.189 | −0.039 | 0.038 | −0.231 | 0.002 | 0.180 |
| Body depth | −0.204 | −0.009 | −0.236 | −0.259 | 0.143 | −0.274 |
| Predorsal length | −0.191 | −0.007 | 0.140 | −0.231 | −0.094 | 0.166 |
| Dorsal fin base length | −0.189 | 0.051 | −0.326 | −0.247 | 0.177 | −0.186 |
| Dorsal fin height | −0.149 | 0.281 | −0.036 | −0.188 | −0.271 | −0.418 |
| Unsegmented dorsal fin height | −0.126 | 0.902 | 0.039 | −0.179 | −0.864 | −0.115 |
| Post-dorsal length | −0.194 | −0.064 | −0.016 | −0.241 | 0.039 | 0.327 |
| Dorsal-pelvic length | −0.211 | −0.022 | −0.257 | −0.271 | 0.163 | −0.263 |
| Prepectoral length | −0.179 | −0.015 | 0.264 | −0.208 | −0.158 | 0.276 |
| Pectoral fin length | −0.186 | 0.056 | −0.084 | −0.232 | 0.009 | −0.029 |
| Prepelvic length | −0.191 | −0.071 | 0.072 | −0.229 | 0.015 | 0.156 |
| Pelvic fin length | −0.183 | −0.010 | −0.138 | −0.229 | 0.094 | −0.111 |
| Anal fin base length | −0.203 | 0.002 | −0.195 | −0.257 | 0.133 | 0.178 |
| Anal fin height | −0.182 | 0.084 | −0.134 | −0.235 | −0.003 | −0.140 |
| Caudal peduncle length | −0.200 | −0.042 | 0.106 | −0.242 | −0.038 | 0.435 |
| Maximum caudal peduncle height | −0.195 | −0.056 | −0.182 | −0.246 | 0.153 | −0.201 |
| Minimum caudal peduncle height | −0.202 | −0.054 | −0.152 | −0.253 | 0.126 | −0.187 |
| Pre-anal length | −0.199 | −0.046 | 0.026 | −0.245 | 0.017 | 0.188 |
| Head length | −0.181 | −0.019 | 0.300 | - | - | - |
| Pre-operculum length | −0.179 | −0.002 | 0.313 | - | - | - |
| Head width | −0.199 | −0.005 | 0.029 | - | - | - |
| Inter-orbital distance | −0.230 | −0.117 | −0.179 | - | - | - |
| Lower jaw length | −0.195 | −0.043 | 0.443 | - | - | - |
| Mouth width | −0.238 | −0.176 | −0.011 | - | - | - |
| Eye diameter | −0.110 | 0.112 | 0.181 | - | - | - |
| Inter-nasal distance | −0.225 | −0.067 | −0.072 | - | - | - |
| Snout length | −0.217 | −0.099 | 0.259 | - | - | - |
| L. mawambiensis | L. longidorsalis | L. maw x long 2 | L. maw x long 1 | L. macroceps | L. mac x maw | L. sp. ‘thick lip’ | L. caudovittatus | |
|---|---|---|---|---|---|---|---|---|
| (a) | n = 149 | n = 16 | n = 16 | n = 9 | n = 13 | n = 1 | n = 16 | n = 1 |
| Standard length (mm) | 53.3–220.0 | 93.4–322.0 | 63.0–180.0 | 85.9–164.6 | 123.2–232.2 | 170.5 | 93.0–190.1 | 145.4 |
| Measurements in %SL | ||||||||
| Body depth | 22.7–37.8 | 33.4–39.6 | 31.2–39.4 | 32.4–37.0 | 24.4–29.5 | 30.1 | 25.3–31.2 | 30.3 |
| Predorsal length | 47.1–57.3 | 45.6–50.2 | 46.0–51.0 | 45.2–49.0 | 51.6–55.8 | 52.7 | 50.2–54.3 | 52.6 |
| Dorsal fin base length | 13.7–19.8 | 19.9–23.4 | 17.5–19.7 | 18.5–21.5 | 12.8–15.9 | 17.2 | 11.9–15.0 | 12.0 |
| Dorsal fin height | 24.8–43.9 | 21.8–33.0 | 25.9–34.3 | 25.2–28.2 | 18.9–24.2 | 28.3 | 25.6–30.0 | 17.7 |
| Unsegmented dorsal fin height | 17.7–35.8 | 10.4–19.2 | 14.4–24.2 | 12.3–17.0 | 8.8–13.9 | 18.9 | 10.0–15.7 | 7.1 |
| Segmented dorsal fin height | 7.5–34.0 | 26.0–46.1 | 9.9–45.5 | 24.1–37.5 | 28.6–48.8 | 37.7–58.6 | 35.0 | |
| Post-dorsal length | 28.3–38.8 | 29.2–38.6 | 34.9–39.0 | 31.8–38.7 | 27.6–35.8 | 34.5 | 33.0–42.1 | 37.0 |
| Dorsal-pelvic length | 24.6–35.7 | 32.3–37.4 | 28.2–36.4 | 30.3–36.8 | 23.2–28.8 | 29.6 | 23.3–30.9 | 26.1 |
| Pre-pectoral length | 25.1–33.1 | 21.4–25.3 | 23.2–28.6 | 22.2–25.4 | 27.7–31.8 | 29.1 | 26.7–31.6 | 30.3 |
| Pectoral fin length | 19.2–28.2 | 21.4–24.7 | 22.2–24.5 | 21.4–24.1 | 17.7–22.0 | 22.2 | 18.3–22.9 | 20.6 |
| Pre-pelvic length | 50.1–57.7 | 50.5–56.9 | 49.6–56.3 | 50.5–56.9 | 53.4–59.8 | 54.5 | 51.7–57.3 | 55.3 |
| Pelvic fin length | 16.8–22.9 | 19.1–24.0 | 19.4–22.4 | 19.9–23.3 | 16.0–19.1 | 19.8 | 16.8–20.1 | 17.1 |
| Anal fin base length | 5.4–9.7 | 7.2–11.1 | 6.8–8.3 | 7.2–9.0 | 6.1–8.1 | 7.6 | 5.7–7.2 | 6.6 |
| Anal fin height | 18.8–26.9 | 18.2–26.9 | 20.9–24.7 | 21.0–25.2 | 14.2–19.3 | 22.1 | 18.2–22.9 | 18.4 |
| Caudal peduncle length | 11.9–19.9 | 13.5–17.0 | 12.5–19.1 | 12.4–16.7 | 12.7–18.5 | 14.8 | 12.9–18.3 | 13.5 |
| Maximum caudal peduncle height | 12.7–17.9 | 15.4–18.3 | 14.9–17.6 | 15.9–17.8 | 12.1–14.4 | 15.6 | 13.0–16.1 | 15.2 |
| Minimum caudal peduncle height | 10.7–15.0 | 13.2–15.0 | 12.6–14.1 | 12.8–14.3 | 10.3–12.5 | 13.2 | 11.4–13.6 | 12.0 |
| Pre-anal length | 69.7–81.2 | 74.8–81.6 | 73.5–79.1 | 71.3–77.8 | 73.1–79.0 | 80.2 | 71.6–79.0 | 74.6 |
| Head length | 25.8–32.3 | 22.0–25.4 | 23.3–27.5 | 22.8–24.6 | 29.3–34.2 | 30.1 | 28.1–30.9 | 29.1 |
| Measurements in %HL | ||||||||
| Pre-operculum length | 69.7–80.6 | 63.8–77.8 | 71.6–79.3 | 70.5–75.4 | 69.5–74.9 | 73.9 | 69.9–83.9 | 71.2 |
| Head width | 49.7–62.8 | 61.1–73.9 | 53.2–63.0 | 55.8–67.6 | 40.4–47.8 | 49.8 | 46.0–61.4 | 57.9 |
| Inter-orbital distance | 22.9–35.7 | 36.4–57.7 | 30.7–41.0 | 35.0–45.5 | 19.8–23.7 | 29.2 | 27.2–38.8 | 35.9 |
| Lower jaw length | 30.4–42.7 | 24.7–48.8 | 30.9–40.9 | 27.7–35.6 | 43.1–48.7 | 44.2 | 31.4–45.5 | 36.2 |
| Mouth width | 14.2–30.0 | 21.6–43.8 | 16.9–29.2 | 20.4–33.2 | 17.0–26.1 | 18.5 | 15.7–27.8 | 24.8 |
| Eye diameter | 21.6–41.9 | 18.6–31.6 | 25.9–37.8 | 26.1–32.7 | 17.8–23.2 | 24.5 | 22.5–38.3 | 23.4 |
| Inter-nasal distance | 13.5–23.1 | 22.1–31.2 | 14.7–22.6 | 17.1–25.0 | 9.9–16.1 | 18.1 | 14.6–23.5 | 19.4 |
| Snout length | 26.8–43.5 | 32.0–47.7 | 31.4–44.0 | 31.1–39.4 | 30.5–35.7 | 34.2 | 37.3–45.2 | 36.9 |
| Anterior barbel length | 16.1–40.7 | 7.8–12.6 | 10.4–23.6 | 21.8 | 14.4–21.5 | 15.8 | ||
| Posterior barbel length | 19.9–42.9 | 1.5–7.2 | 5.1–17.1 | 3.6–9.2 | 15.0–27.0 | 27.0 | 17.1–25.4 | 19.6 |
| Premaxillary pedicel length | 9.0–23.6 | 15.3–32.4 | 15.8–22.4 | 20.2 | 20.7–31.3 | 21.0 | ||
| L. mawambiensis | L. longidorsalis | L. mawx long 2 | L. mawx long 1 | L. macroceps | L. macx maw | L.sp. ‘thick lip’ | L. caudovittatus | |
| (b) | n= 149 | n= 16 | n= 16 | n= 9 | n= 13 | n= 1 | n= 16 | n= 1 |
| Total number of lateral line scales | 21–28 | 23–27 | 24–28 | 24–27 | 31–36 | 29 | 24–27 | 24 |
| Number of predorsal scales | 7–11 | 8–10 | 8–11 | 8–9 | 10–14 | 10 | 7–10 | 8 |
| Number of scales above the lateral line | 3.5–5.5 | 4.5–5.5 | 4.5–5.5 | 4.5–5.5 | 5.5–5.5 | 5.5 | 3.5–4.5 | 4.5 |
| Lateral line-pelvic scales | 1.5–2.5 | 2–2.5 | 2–2 | 2–3 | 2–3 | 2.5 | 1.5–2 | 2.5 |
| Lateral line–ventral midline scales | 3.5–5.5 | 3.5–4.5 | 3.5–4.5 | 4.5–4.5 | 4.5–5.5 | 4.5 | 3.5–4.5 | 4.5 |
| Caudal peduncle scales | 12 | 12 | 12 | 12 | 12 | 12 | 12 | 12 |
| Number of dorsal-fin base scales | 5–10 | 6–10 | 6–10 | 7–11 | 6–10 | 8 | 5–8 | 8 |
| Number of anal-fin base scales | 1–5 | 2–4 | 3–4 | 3–4 | 3–5 | 4 | 3–5 | 3 |
| Number of unbranched dorsal fin rays | 4 | 4 | 4 | 4 | 4 | 4 | 4 | 4 |
| Number of branched dorsal fin rays | 8–11 | 11–12 | 10–12 | 11–12 | 10–11 | 10 | 9–10 | 10 |
| Number of unbranched anal fin rays | 3 | 3 | 3 | 3 | 3 | 3 | 3 | 3 |
| Number of branched anal fin rays | 6 | 6 | 6 | 6 | 6 | 6 | 6 | 6 |
| Number of branched pectoral fin rays | 14–17 | 13–15 | 14–16 | 13–15 | 13–15 | 14 | 14–16 | 15 |
| Number of branched pelvic fin rays | 7–9 | 8–9 | 8–9 | 8–9 | 8–8 | 8 | 8–9 | 8 |
| Number of caudal fin rays | 17 | 17 | 17 | 17 | 17 | 17 | 17 | 17 |
| Number of scales between dorsal and caudal fin | 9–15 | 11–13 | 11–14 | 10–13 | 12–18 | 13 | 11–15 | 11 |
| Number of lateral line scales between anterior dorsal- and pelvic-fin base | 0.5–2.5 | 2–3.5 | 1–2.5 | 2–2.5 | 1–2 | 1.5 | 1–2 | 2 |
| Number of gill rakers on lower branch of first gill arch | 9–15 | 10–14 | 10–13 | 10–13 | 5–7 | 9 | 13–17 | 9 |
| Number of gill rakers on upper branch of first gill arch | 2–6 | 1–4 | 2–6 | 2–4 | 2–3 | 4 | 3–6 | 4 |
| Total number of gill rakers on the first gill arch | 14–19 | 14–18 | 14–18 | 14–18 | 9–11 | 14 | 19–23 | 14 |
| L. caudovittatus | L. fasolt | L. humphrii | L. longidorsalis | L. macrolepidotus | L. mawambi | L. mawambiensis | L. mirabilis | |
|---|---|---|---|---|---|---|---|---|
| (a) | 2 syntypes | holotype | holotype & 10 paratypes | holotype | 6 syntypes | holotype | 9 syntypes | holotype |
| Standard length (mm) | 71.2–74.7 | 464.0 | 143.2 & 80.1–207.7 | 234.3 | 62.7–129.4 | 61.7 | 92.3–173.9 | 334.0 |
| Measurements (in % SL) | ||||||||
| Body depth | 25.3–26.5 | 31.8 | 27.2 & 22.6–29.6 | 34.2 | 29.0–34.8 | 28.6 | 28.9–34.4 | 31.7 |
| Predorsal length | 52.2–52.9 | 53.2 | 48.8 & 47.2–51.1 | 46.6 | 46.0–50.1 | 53.3 | 50.0–55.6 | 54.5 |
| Dorsal fin base length | 13.6–16.5 | 13.7 | 16.1 & 15.2–17.3 | 23.6 | 15.3–22.1 | 14.0 | 12.7–19.2 | 18.1 |
| Dorsal fin height | 19.8–20.5 | 18.9 | 22.1 & 18.3–29 | 23.5–28.6 | 20.6 | 23.6–32.0 | 16.0 | |
| Unsegmented dorsal fin height | 8.1–9.9 | 8.4 | 20.7 & 15–27.6 | 17.8 | 10.7–15.6 | 12.8 | 19.2–26.4 | 11.5 |
| Segmented dorsal fin height | 35.5 | 37.4–44.2 | 27.2 | 34.7–34.7 | 14.7 | |||
| Post-dorsal length | 34.8–36.3 | 35.3 | 39 & 31.9–41.3 | 35.5 | 29.8–34.9 | 33.7 | 31.4–37.8 | 31.3 |
| Dorsal-pelvic length | 24.6–27.8 | 29.3 | 26.5 & 22.1–26.4 | 32.0 | 25.0–32.3 | 28.4 | 28.2–33.0 | 32.1 |
| Pre-pectoral length | 27.4–28.9 | 30.1 | 26 & 26.3–29.8 | 20.7 | 24.6–30.0 | 29.0 | 26.8–32.4 | 28.0 |
| Pectoral fin length | 16.7–19.7 | 21.2 | 21.6 & 19.5–21.7 | 21.7 | 22.5–25.0 | 19.9 | 21.0–25.5 | 21.5 |
| Pre-pelvic length | 54.4–55.0 | 57.0 | 51.7 & 50.7–54 | 53.7 | 52.9–56.7 | 54.7 | 50.8–55.4 | 56.2 |
| Pelvic fin length | 18.7–19.5 | 17.0 | 18.7 & 16.2–22 | 21.8 | 19.0–23.3 | 19.0 | 17.8–21.5 | 17.1 |
| Anal fin base length | 6.2–7.4 | 8.4 | 8.4 & 6.8–8.3 | 10.0 | 6.9–8.3 | 8.5 | 6.4–8.9 | 7.2 |
| Anal fin height | 17.7–20.1 | 15.0 | 19.7 & 18.8–22 | 24.1 | 20.8–23.2 | 18.5 | 19.9–25.7 | 19.4 |
| Caudal peduncle length | 12.8–17.0 | 15.2 | 13.8 & 13.7–17.9 | 16.0 | 12.7–17.0 | 13.5 | 13.4–17.3 | 14.0 |
| Maximum caudal peduncle height | 11.8–16.1 | 13.2 | 13.8 & 12–13.4 | 15.8 | 14.5–15.9 | 15.1 | 13.3–18.1 | 14.2 |
| Minimum caudal peduncle height | 9.8–13.8 | 12.6 | 9.9 & 10–11.4 | 12.8 | 12.3–13.7 | 12.8 | 11.6–14.9 | 12.6 |
| Pre-anal length | 74.4–77.7 | 78.0 | 76.5 & 69–78.5 | 77.6 | 72.6–75.8 | 78.6 | 73.6–80.9 | 81.8 |
| Head length | 27.5–27.7 | 28.5 | 26.2 & 27.6–29.8 | 20.8 | 25.0–30.0 | 28.8 | 26.8–29.6 | 26.2 |
| Measurements (in % HL) | ||||||||
| Pre-operculum length | 72.0–74.0 | 73.0 | 72 & 69.4–74 | 71.0 | 72.5–78.9 | 75.5 | 70.8–76.8 | 74.2 |
| Head width | 50.7–53.6 | 61.7 | 55.2 & 50.3–55.8 | 73.8 | 50.0–62.2 | 46.8 | 51.3–58.8 | 59.2 |
| Inter-orbital distance | 32.1–33.8 | 46.1 | 30.4 & 26.3–34.2 | 53.0 | 31.4–37.2 | 25.3 | 26.5–36.0 | 41.2 |
| Lower jaw length | 34.3–35.7 | 34.6 | 35.2 & 32.3–39.1 | 36.1 | 29.6–34.1 | 42.4 | 32.8–38.2 | 36.3 |
| Mouth width | 20.8–22.4 | 39.9 | 19.2 & 17.9–21.9 | 44.6 | 20.7–26.9 | 21.1 | 19.1–23.7 | 29.2 |
| Eye diameter | 31.1–32.4 | 18.7 | 26.9 & 20.5–32.5 | 29.2 | 28.5–32.4 | 33.7 | 24.4–33.3 | 24.8 |
| Inter-nasal distance | 17.9–19.9 | 27.5 | 17.1 & 15.8–20 | 25.6 | 16.5–24.8 | 12.1 | 14.5–19.6 | 22.2 |
| Snout length | 30.6–32.9 | 36.2 | 34.4 & 21–38.5 | 32.3 | 32.4–38.2 | 31.7 | 28.8–35.7 | 35.0 |
| Anterior barbel length | 18.4–21.7 | 17.1 | 23.5 & 14.8–26.3 | 13.4 | 20.9–31.9 | 19.0 | ||
| Posterior barbel length | 15.5–28.1 | 22.1 | 28.8 & 24–30.7 | 3.3 | 3.7–9.4 | 21.6 | 22.1–37.4 | 22.3 |
| L. caudovittatus | L. fasolt | L. humphrii | L. longidorsalis | L. macrolepidotus | L. mawambi | L. mawambiensis | L.mirabilis | |
| (b) | 2 syntypes | holotype | holotype + 10 paratypes | holotype | 6 syntypes | holotype | 9 syntypes | holotype |
| Total number of lateral line scales | 26 | 26 | 26 & 24–28 | 29 | 25–27 | 28 | 23–26 | 31 |
| Number of predorsal scales | 10 | 8.5 | 8 & 8–10 | 10 | 7–8 | 10 | 8–10 | 14 |
| Number of scales above the lateral line | 4.5 | 4.5 | 4.5 & 4.5–4.5 | 4.5 | 4.5–5.5 | 5.5 | 4.5–4.5 | 5.5 |
| Lateral line–pelvic scales | 1.5–2.0 | 3 | 2 & 2–2.5 | 2 | 2.0–2.5 | 2.0 | 2.0 | 3.0 |
| Lateral line–ventral midline scales | 4.5 | 4.5 | 4.5 & 4.5–5.5 | 4.5 | 4.5 | 4.5 | 4.5–5.5 | 5.5 |
| Caudal peduncle scales | 12. | 12 | 12 & 12–12 | 12 | 12 | 12 | 12 | 12 |
| Number of dorsal-fin base scales | 7 | 6 | 8 & 5–9 | 6 | 6–11 | 5 | 5–8 | 7 |
| Number of anal-fin base scales | 2–3 | 3 | 5 & 3–4 | 4 | 3–4 | 4 | 3–4 | 4 |
| Number of unbranched dorsal fin rays | 4 | 4 | 4 | 4 | 4 | 4 | 4 | 4 |
| Number of branched dorsal fin rays | 10 | 10 | 10 & 9–10 | 13 | 10–12 | 11 | 10 | 11 |
| Number of unbranched anal fin rays | 3 | 3 | 3 | 3 | 3 | 3 | 3 | 3 |
| Number of branched anal fin rays | 6 | 6 | 6 & 6–6 | 6 | 6 | 6 | 6 | 6 |
| Number of branched pectoral fin rays | 14–15 | 15 | 16 & 15–17 | 14 | 13–16 | 15 | 14–17 | 15 |
| Number of branched pelvic fin rays | 8 | 8 | 8 & 8–8 | 8 | 8 | 8 | 8 | 8 |
| Number of caudal fin rays | 17 | 17 | 17 & 17–17 | 17 | 17 | 17 | 17 | 17 |
| Number of scales between dorsal and caudal fin | 10–14 | 14 | 13 & 9–14 | 11.5 | 10–12 | 13 | 12–13 | 12 |
| Number of lateral line scales between anterior dorsal- and pelvic-fin base | 2.0–2.5 | 1.5 | 2 & 1.5–2.5 | 3 | 2.5–3 | 1.5 | 1.0–2.5 | 1 |
| Number of gill rakers on lower branch of first gill arch | 9–11 | 11 | 11 & 9–12 | 13 | 12–13 | 8 | 11–13 | 10 |
| Number of gill rakers on upper branch of first gill arch | 4 | 3 | 3 & 2–4 | 3 | 2–3 | 3 | 3–4 | 2 |
| Total number of gill rakers on the first gill arch | 14–16 | 15 | 15 & 13–16 | 17 | 15–16 | 12 | 15–18 | 13 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Decru, E.; Snoeks, J.; Walanga, A.; Vreven, E.J.W.M.N. Disentangling the Diversity of the Labeobarbus Taxa (Cypriniformes: Cyprinidae) from the Epulu Basin (DR Congo, Africa). Diversity 2022, 14, 1022. https://doi.org/10.3390/d14121022
Decru E, Snoeks J, Walanga A, Vreven EJWMN. Disentangling the Diversity of the Labeobarbus Taxa (Cypriniformes: Cyprinidae) from the Epulu Basin (DR Congo, Africa). Diversity. 2022; 14(12):1022. https://doi.org/10.3390/d14121022
Chicago/Turabian StyleDecru, Eva, Jos Snoeks, Albert Walanga, and Emmanuel J. W. M. N. Vreven. 2022. "Disentangling the Diversity of the Labeobarbus Taxa (Cypriniformes: Cyprinidae) from the Epulu Basin (DR Congo, Africa)" Diversity 14, no. 12: 1022. https://doi.org/10.3390/d14121022