

Morphology of Three Sphyrion (Copepoda: Siphonostomatoida: Sphyriidae) Species Infecting Teleost Fishes off South Africa with the First Description of Males of Two Species


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
3. Results
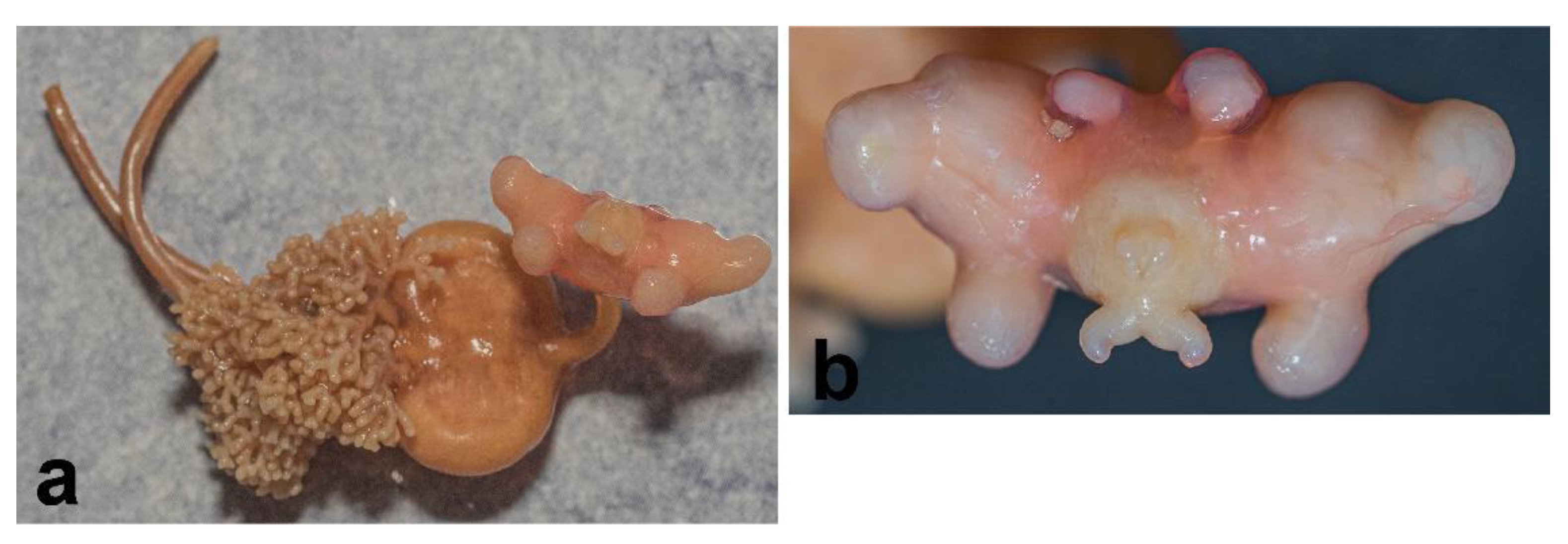
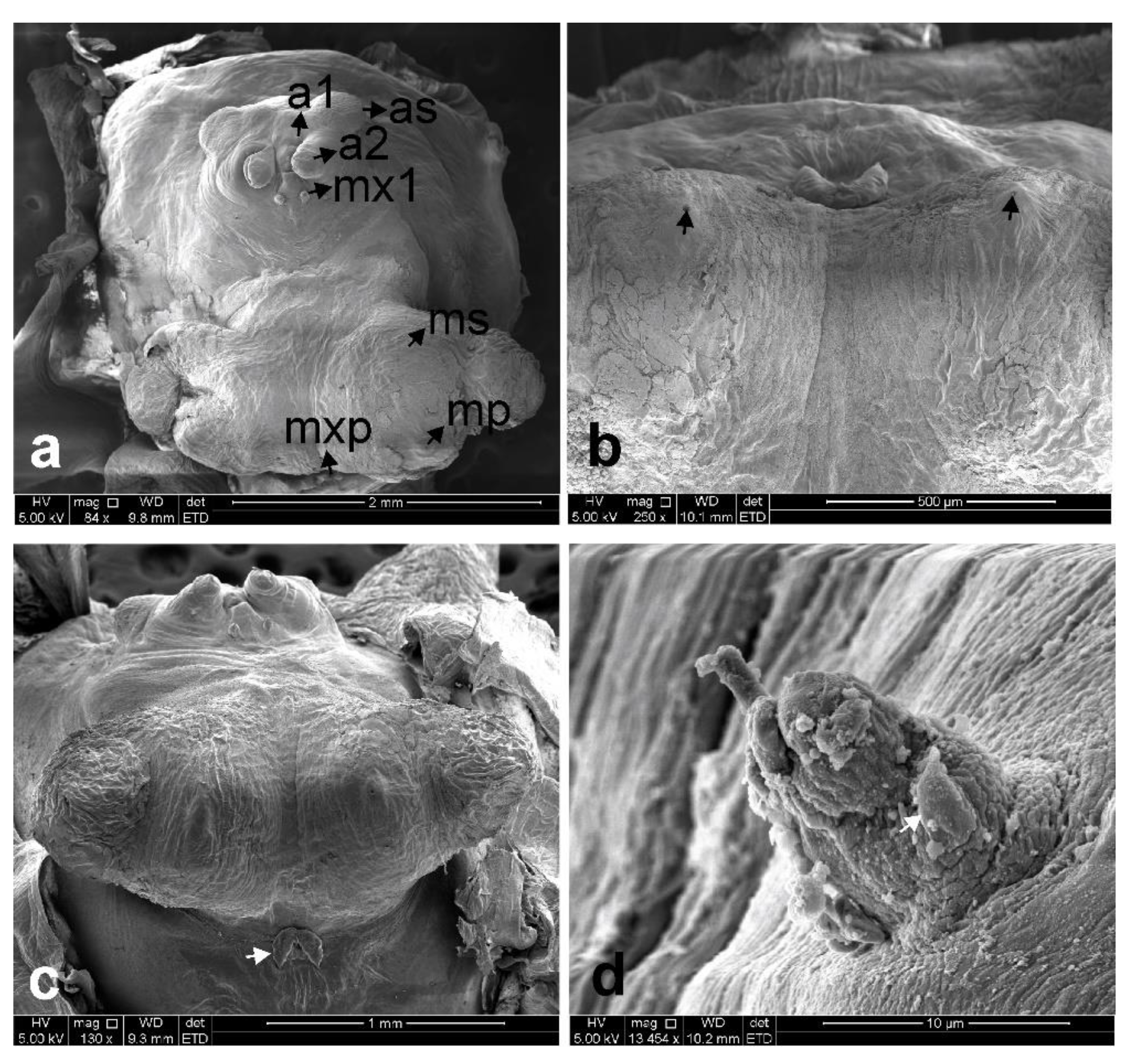
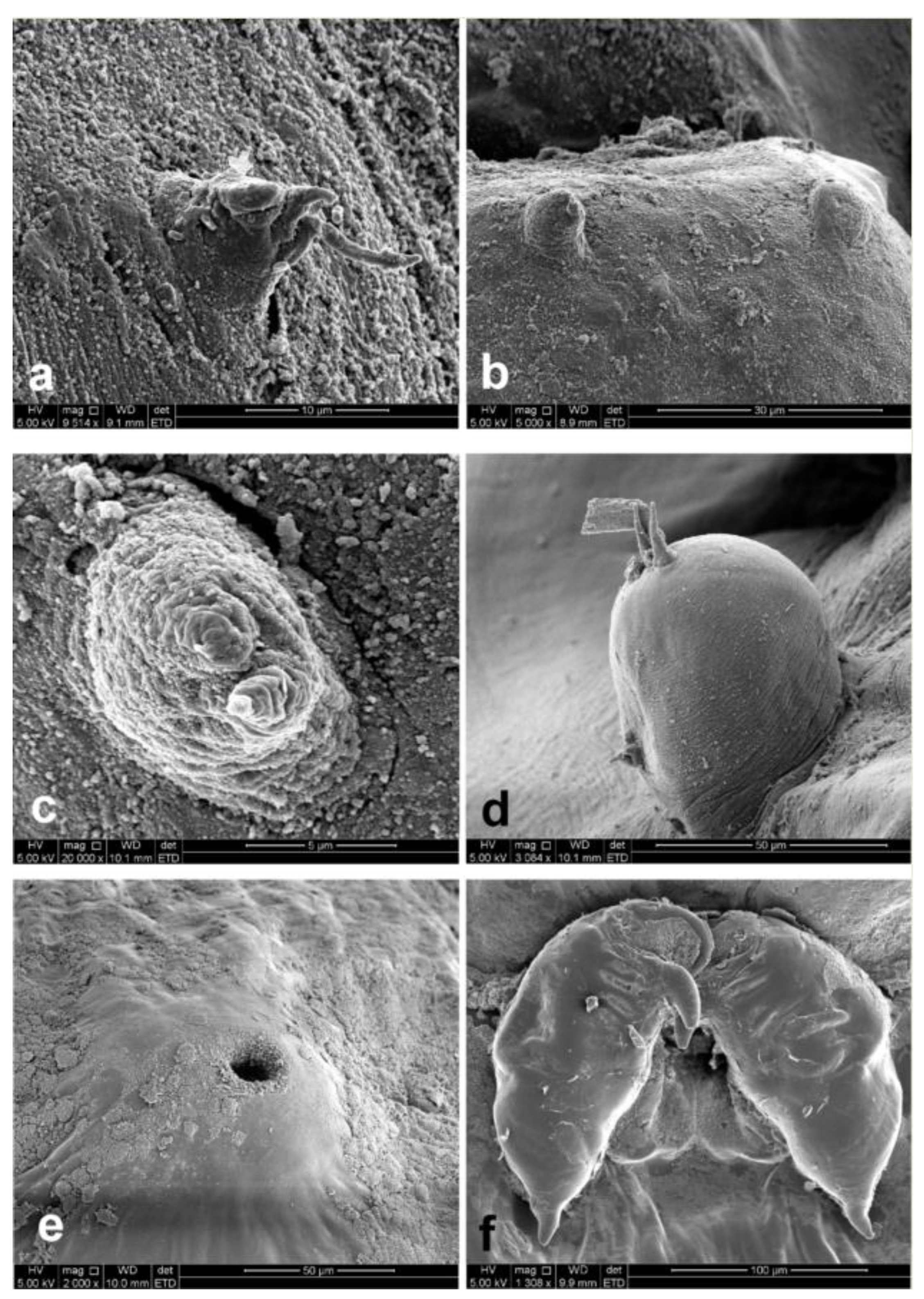
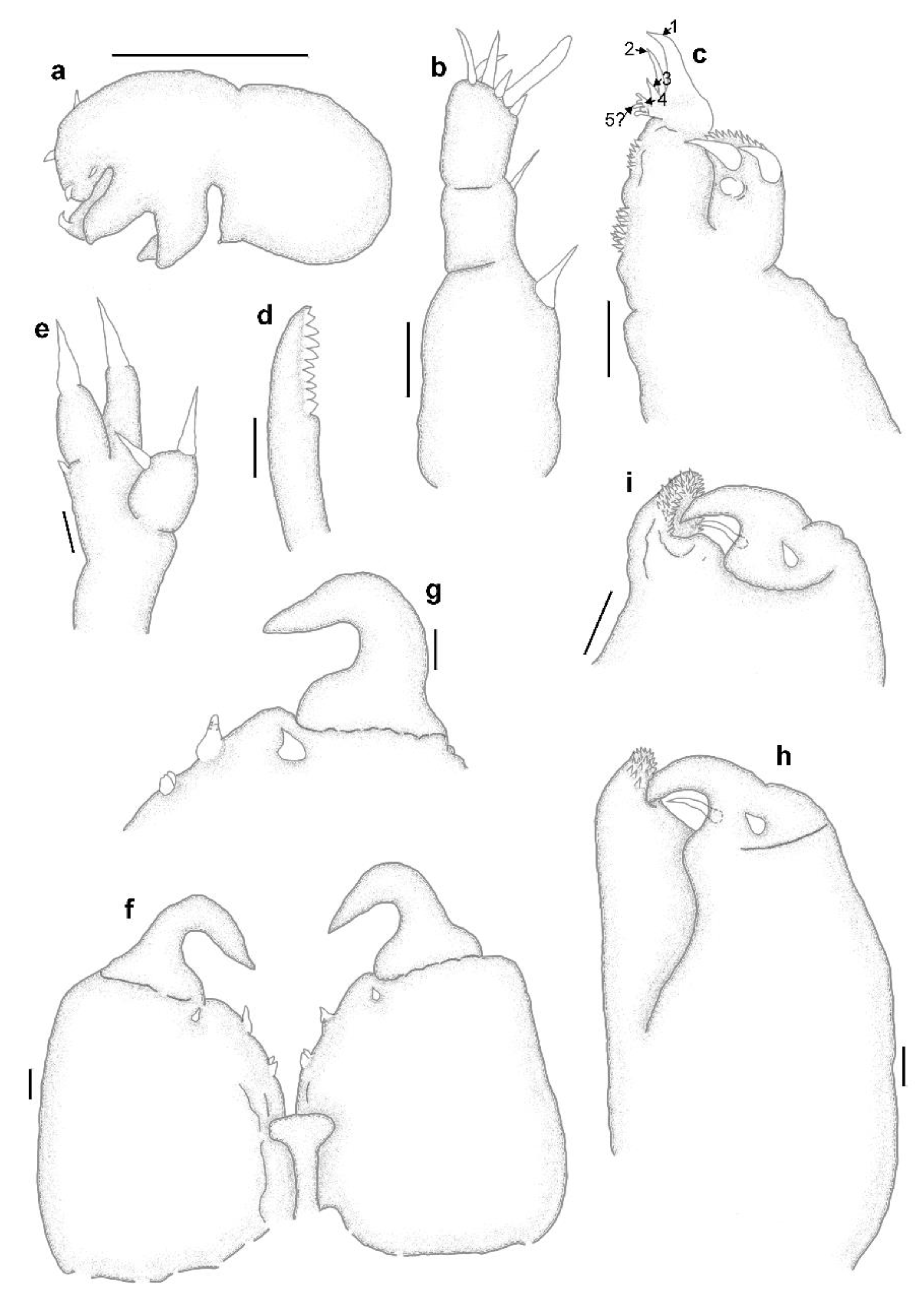
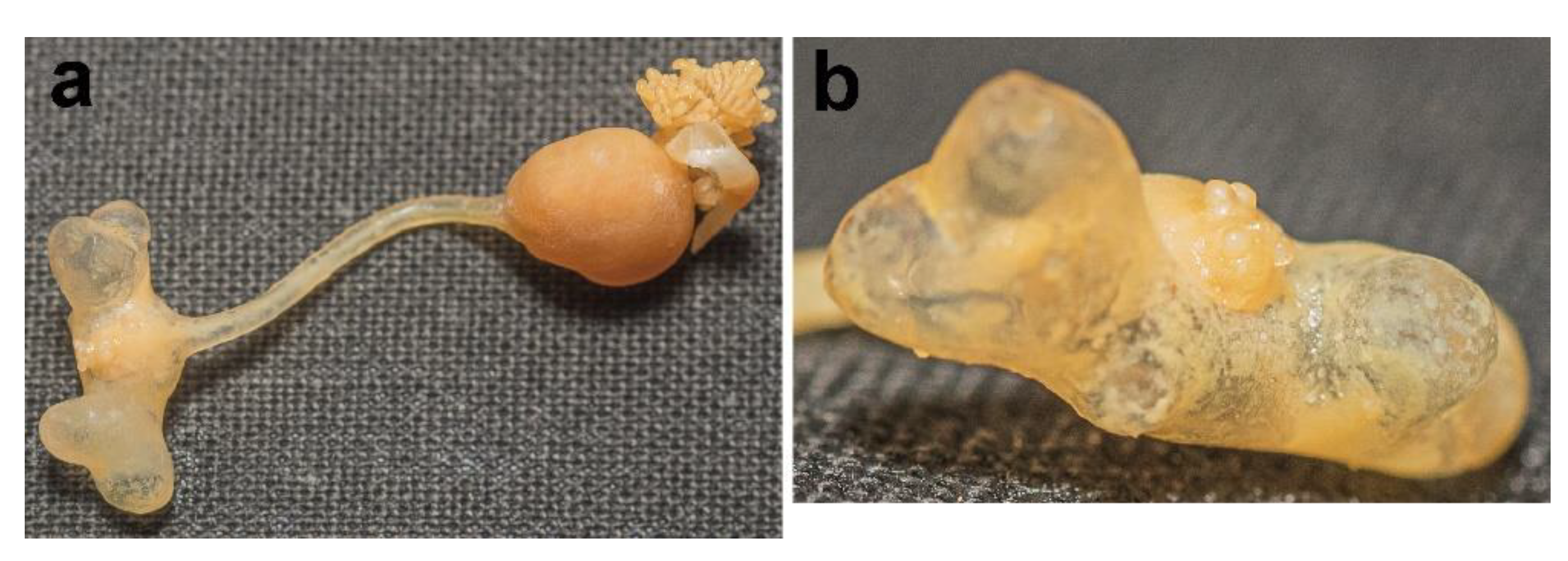
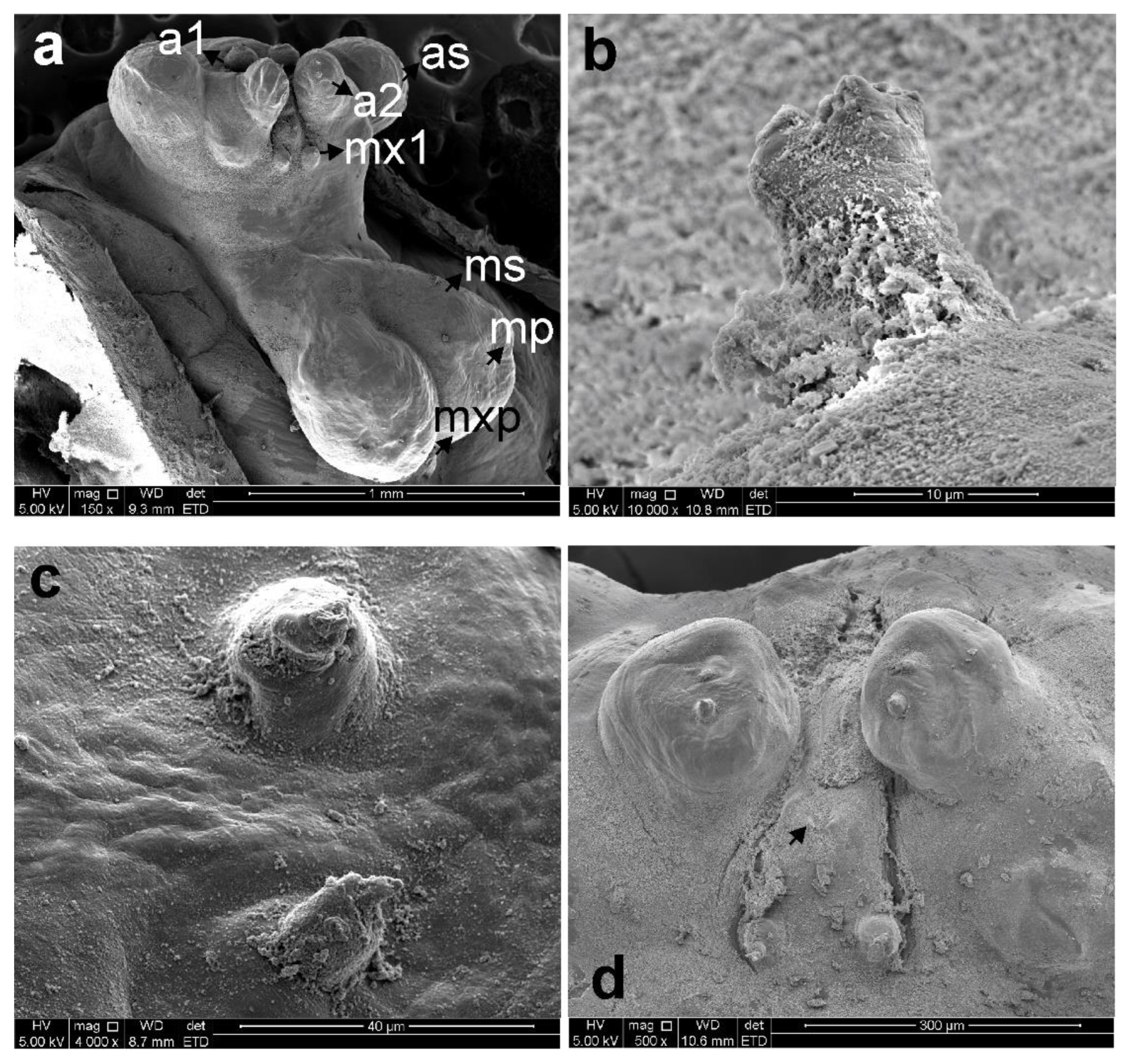
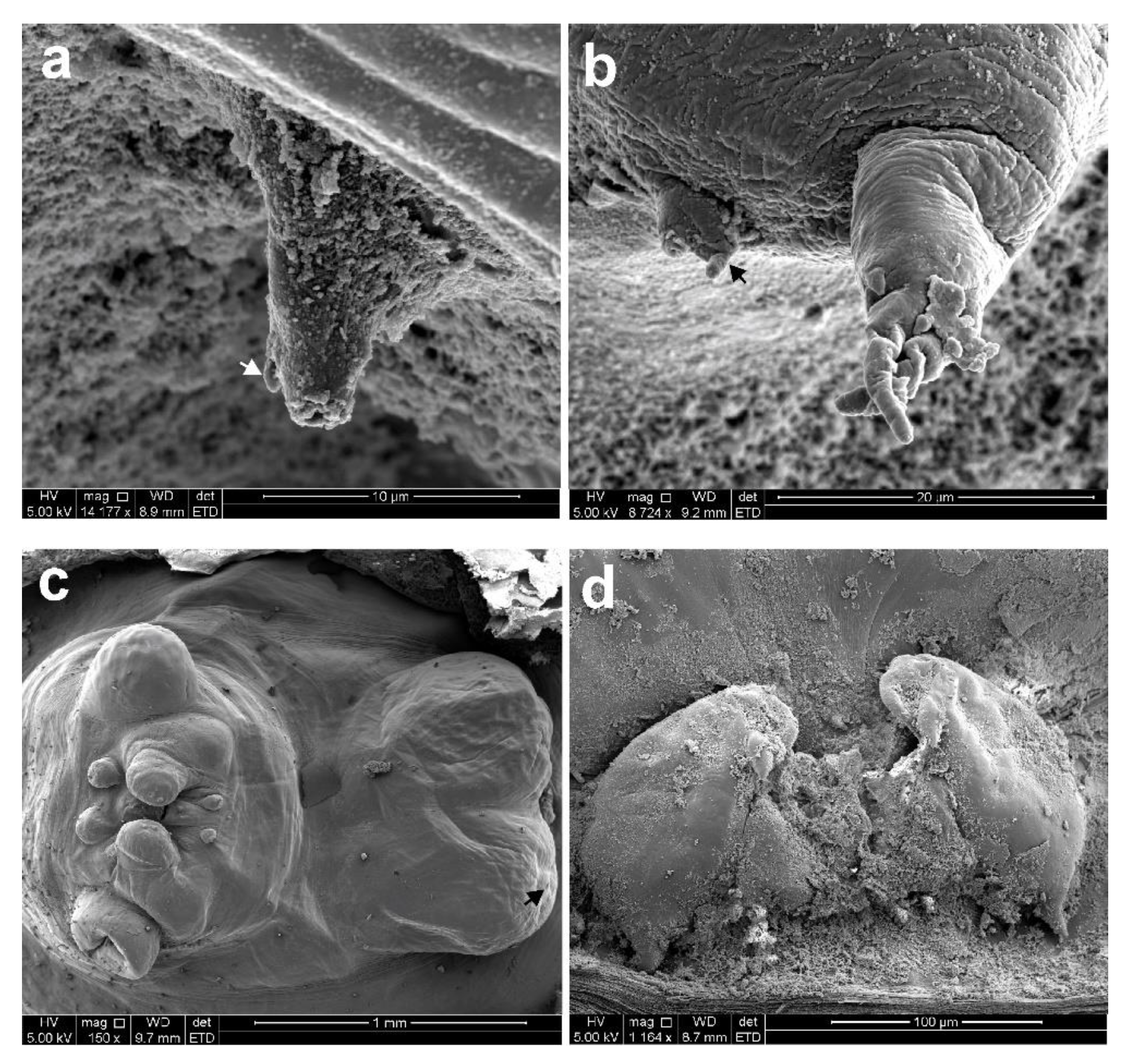
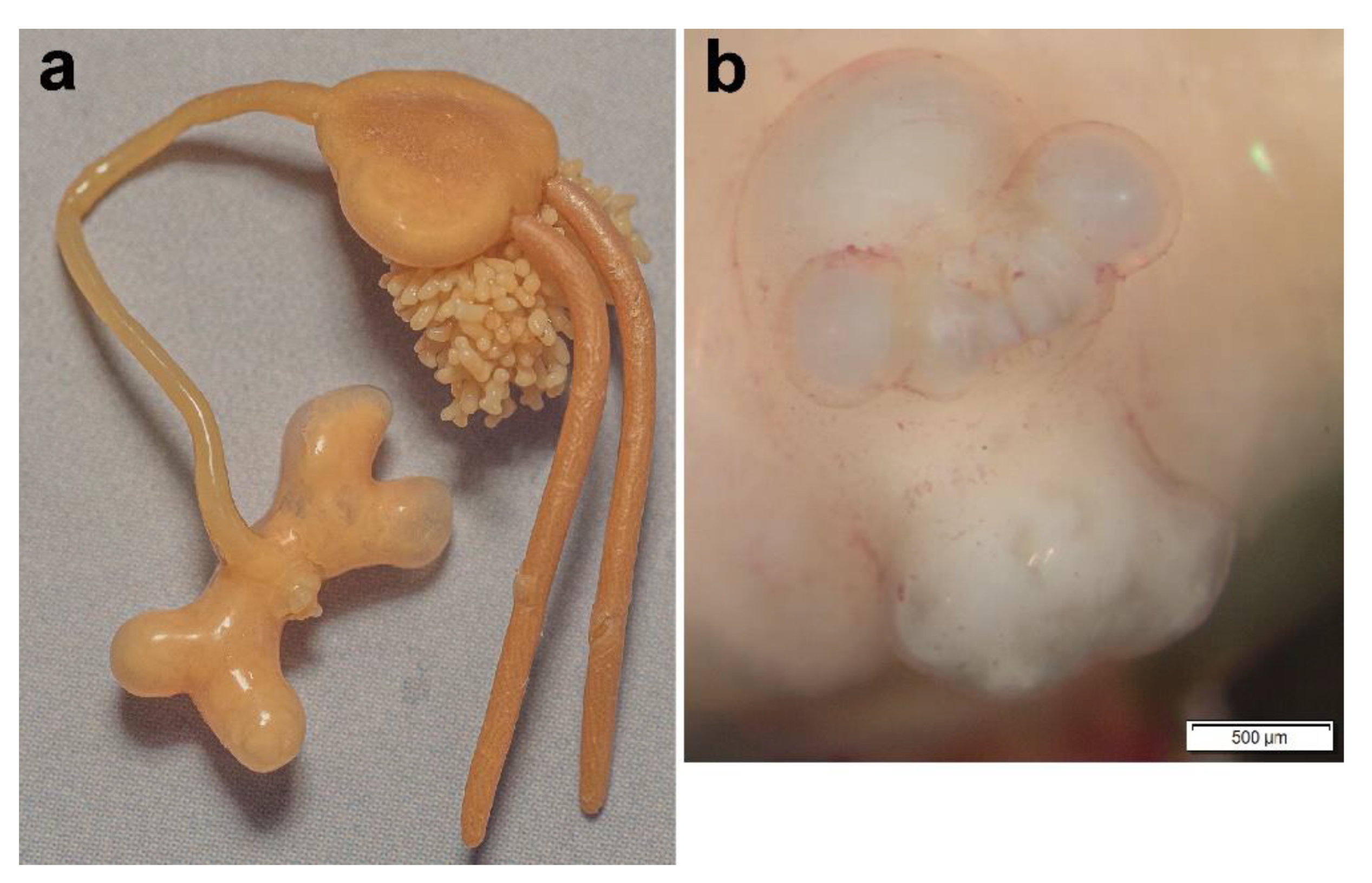
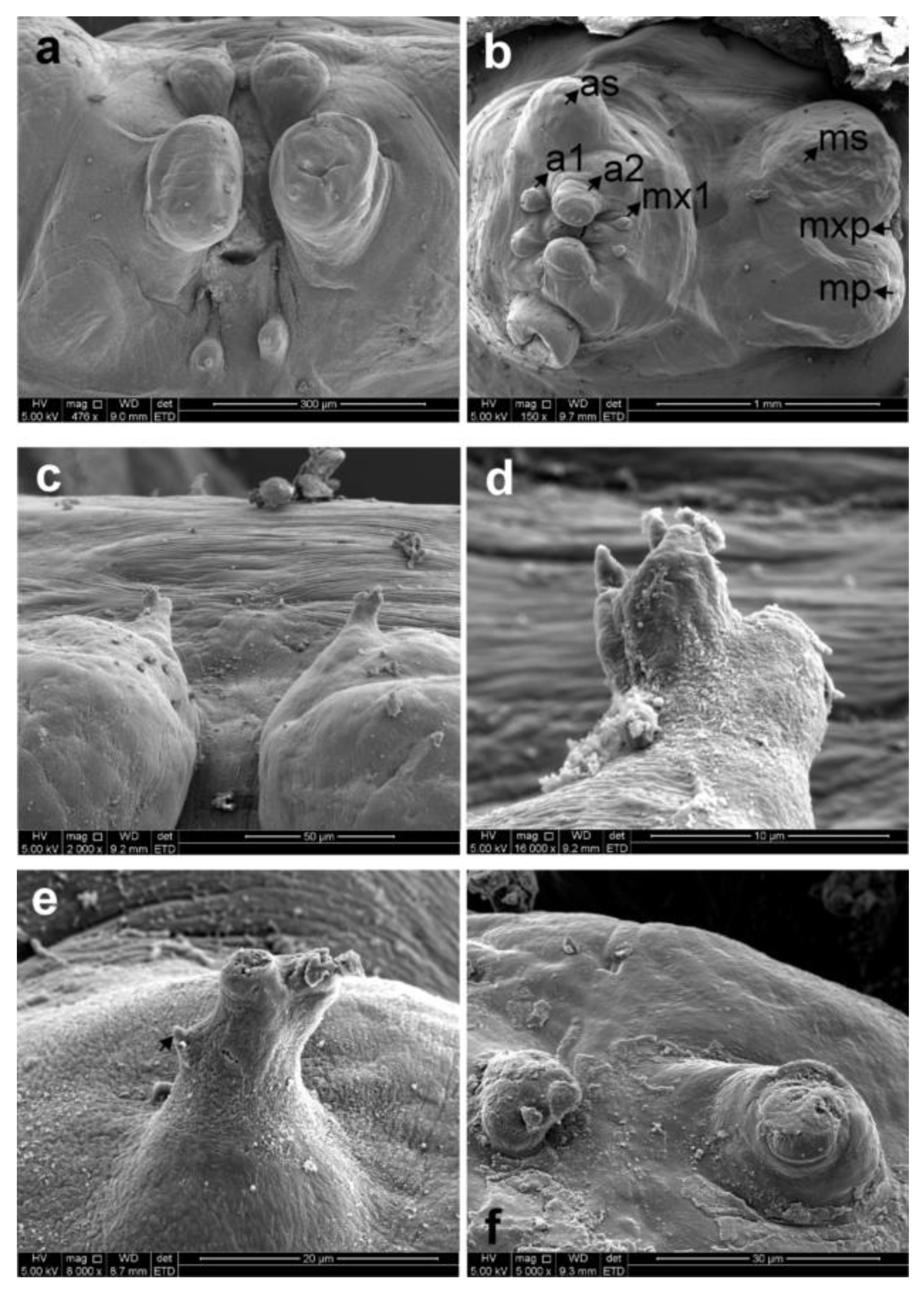
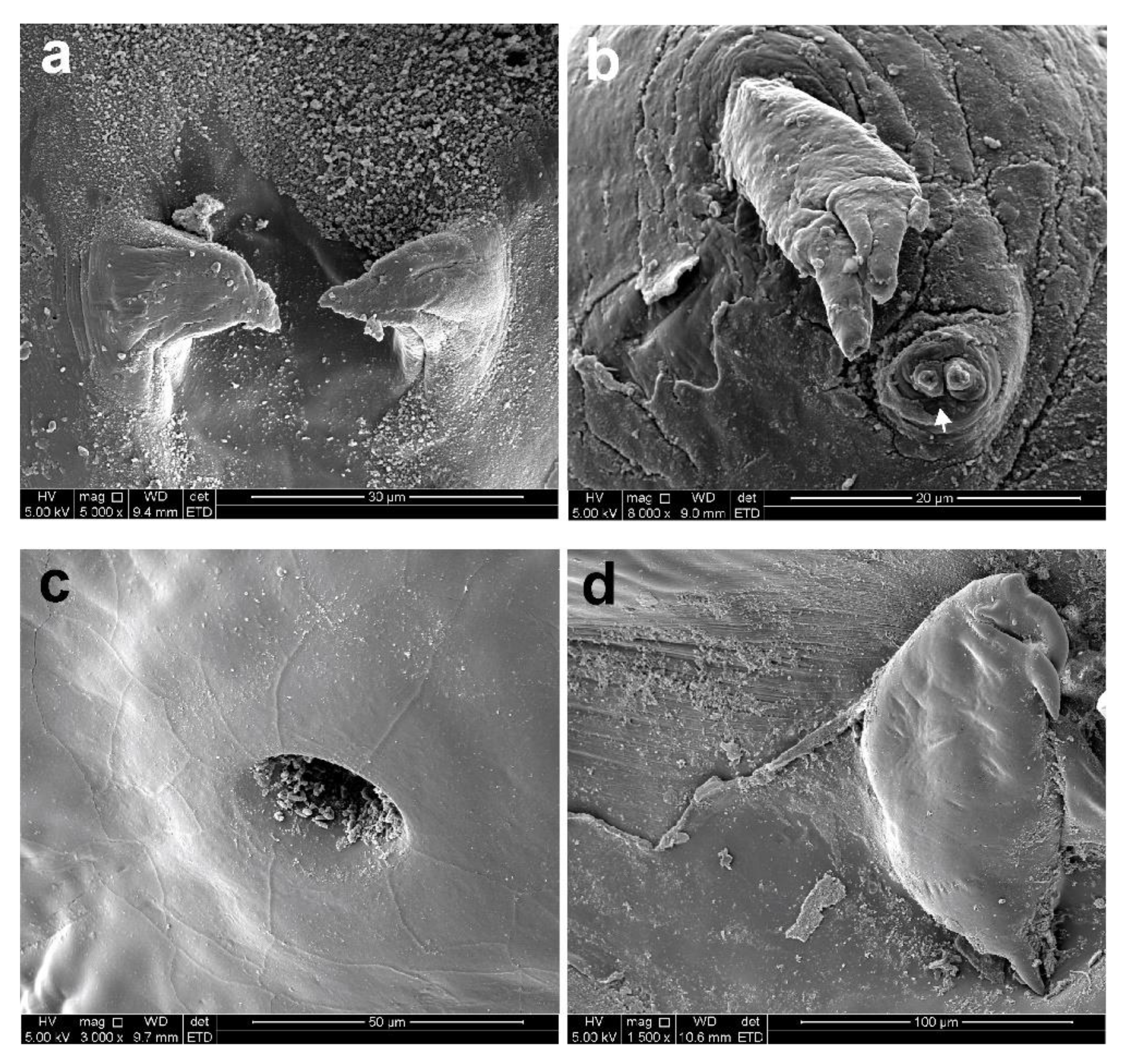
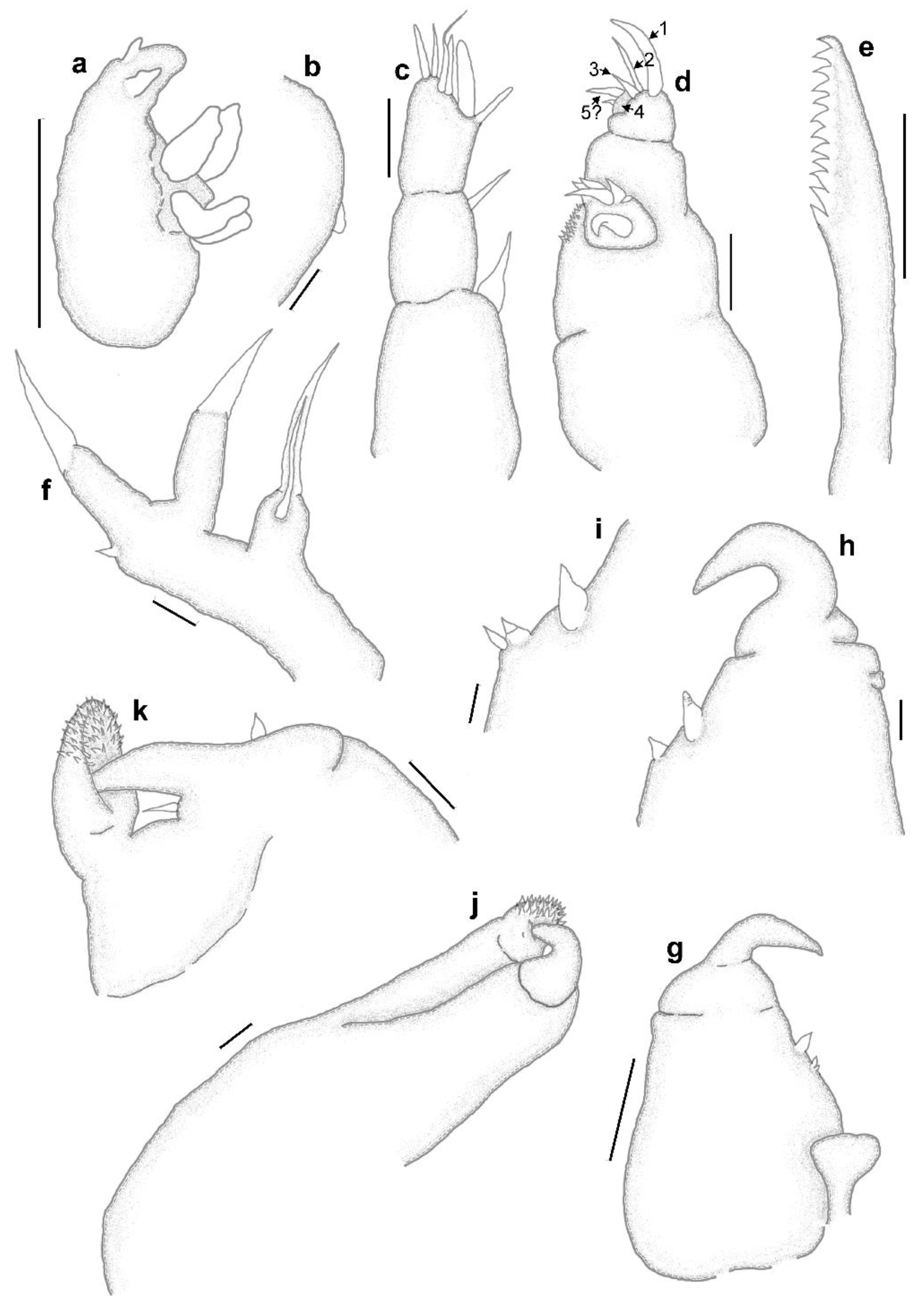
3.1. Sphyrion laevigatum (Quoy and Gaimard, 1824)
3.2. Sphyrion lumpi (Krøyer, 1845)
3.3. Sphyrion quadricornis Gaevskaya and Kovaleva, 1984
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Walter, T.C.; Boxshall, G.A. World of Copepods Database. Sphyrion Cuvier, 1830. 2021. Available online: https://www.marinespecies.org/copepoda/aphia.php?p=taxdetails&id=135656 (accessed on 2 June 2022).
- Dojiri, M.; Deets, G.B. Norkus cladocephalus, new genus, new species (Siphonostomatoida: Sphyriidae), a copepod parasitic on an elasmobranch from southern California waters, with a phylogenetic analysis of the Sphyriidae. J. Crustacean Biol. 1988, 8, 679–687. [Google Scholar] [CrossRef]
- Ho, J.-S. Does Sphyrion lumpi (Kroyer) (Copepoda, Sphyriidae) occur in the Sea of Japan? With discussion on the origin and dispersal of Sphyrion Cuvier, 1830. Rep. Sado Mar. Biol. Stn. Niigata Univ. 1992, 22, 37–48. [Google Scholar]
- Moran, J.; Piasecki, W. External morphology of the male and female of Sphyrion lumpi (Krøyer,1845) (Copepoda; Siphonostomatoida; Sphyriidae). Hydrobiologia 1994, 292, 171–178. [Google Scholar] [CrossRef]
- Kazachenko, V.N. Nahodki predstavitelei roda Sphyrion (Crustacea: Copepoda) u yugo-zapadnyh beregov Afriki. Findings of representatives of the genus Sphyrion (Crustacea: Copepoda) off the south-western coasts of Africa. Nauchnye Tr. Dalrybvtuza 2017, 40, 13–16. [Google Scholar]
- Kensley, B.; Grindley, J.R. South African parasitic Copepoda. Ann. South Afr. Mus. 1973, 62, 69–130. [Google Scholar]
- Gaevskaya, A.V.; Kovaleva, A.A. Rachki roda Sphyrion (Copepoda, Sphyridae) u ryb Atlantiki. Crustacea of the genus Sphyrion (Copepoda, Sphyriidae) in Atlantic fishes. Gidrobiol. Zhurnal Saratov 1984, 20, 41–45. [Google Scholar]
- Boxshall, G.A.; Halsey, S. An Introduction to Copepod Diversity; The Ray Society: London, UK, 2004; p. 966. [Google Scholar]
- Kabata, Z. Parasitic Copepoda of British Fishes; The Ray Society: London, UK, 1979; p. 468. [Google Scholar]
- Barnard, K.H. South African Parasitic Copepoda. Ann. South Afr. Mus. 1955, 41, 223–312. [Google Scholar]
- Ho, J.-S.; Kim, I. Lophoura (Copepoda: Sphyriidae) parasitic on the rattails (Pisces: Macrouridae) in the Pacific, with note on Sphyrion lumpi from the Sea of Japan. Publ. Seto Mar. Biol. Lab. 1989, 34, 37–54. [Google Scholar] [CrossRef] [Green Version]
- Wilson, C.B. North American parasitic copepods belonging to the new family Sphyriidae. Proc. United States Natl. Mus. 1919, 55, 549–604. [Google Scholar] [CrossRef]
- Squires, H.J. Reproduction in Sphyrion lumpi, a copepod parasitic on redfish (Sebastes spp.). J. Fish. Res. Board Can. 1966, 23, 521–526. [Google Scholar] [CrossRef]
- Payne, A.I.L. Observations on some conspicuous parasites of the southern African kingklip Genypterus capensis. S. Afr. J. Mar. Sci. 1986, 4, 163–168. [Google Scholar] [CrossRef] [Green Version]
- Dippenaar, S.M. Reported siphonostomatoid copepods parasitic on marine fishes of southern Africa. Crustaceana 2004, 77, 1281–1328. [Google Scholar] [CrossRef]
- Humes, A.G.; Gooding, R.U. A method for studying the external anatomy of copepods. Crustaceana 1964, 6, 238–240. [Google Scholar] [CrossRef] [Green Version]
- Fricke, R.; Eschmeyer, W.N.; Fong, J.D. Eschmeyer’s Catalog of Fishes: Genera/Species by Family/Subfamily. 2022. Available online: http://researcharchive.calacademy.org/research/ichthyology/catalog/SpeciesByFamily.asp (accessed on 8 March 2022).
- Huys, R.; Boxshall, G.A. Copepod Evolution; The Ray Society: London, UK, 1991; p. 468. [Google Scholar]
- Dippenaar, S.M. Description of four new species and a revision of the genus Tripaphylus Richiardi in Anonymous, 1878 (Copepoda: Siphonostomatoida: Sphyriidae). Syst. Parasitol. 2018, 95, 173–200. [Google Scholar] [CrossRef] [PubMed]
- Russell, B.C.; Golani, D.; Tikochinski, Y. Saurida lessepsianus a new species of lizardfish (Pisces: Synodontidae) from the Red Sea and Mediterranean Sea, with a key to Saurida species in the Red Sea. Zootaxa 2015, 3956, 559–568. [Google Scholar] [CrossRef]
- Boxshall, G.A.; Barton, D.P.; Kirke, A.; Zhu, X.; Johnson, G. Two new parasitic copepods of the family Sphyriidae (Copepoda: Siphonostomatoida) from Australian elasmobranchs. Syst. Parasitol. 2022, 99, 659–669. [Google Scholar] [CrossRef] [PubMed]











Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dippenaar, S.M.; Sebone, M.M. Morphology of Three Sphyrion (Copepoda: Siphonostomatoida: Sphyriidae) Species Infecting Teleost Fishes off South Africa with the First Description of Males of Two Species. Diversity 2022, 14, 929. https://doi.org/10.3390/d14110929
Dippenaar SM, Sebone MM. Morphology of Three Sphyrion (Copepoda: Siphonostomatoida: Sphyriidae) Species Infecting Teleost Fishes off South Africa with the First Description of Males of Two Species. Diversity. 2022; 14(11):929. https://doi.org/10.3390/d14110929
Chicago/Turabian StyleDippenaar, Susan M., and Makwena M. Sebone. 2022. "Morphology of Three Sphyrion (Copepoda: Siphonostomatoida: Sphyriidae) Species Infecting Teleost Fishes off South Africa with the First Description of Males of Two Species" Diversity 14, no. 11: 929. https://doi.org/10.3390/d14110929