

Anianabacter salinae gen. nov., sp. nov. ASV31T, a Facultative Alkaliphilic and Extremely Halotolerant Bacterium Isolated from Brine of a Millennial Continental Saltern
 , , , , , and
, , , , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Strain Isolation and Maintenance
2.2. 16S rRNA Gene-Based Identification and Phylogeny
2.3. Phenotype and Chemotaxonomy
2.4. Genome Sequencing and Analysis
3. Results and Discussion
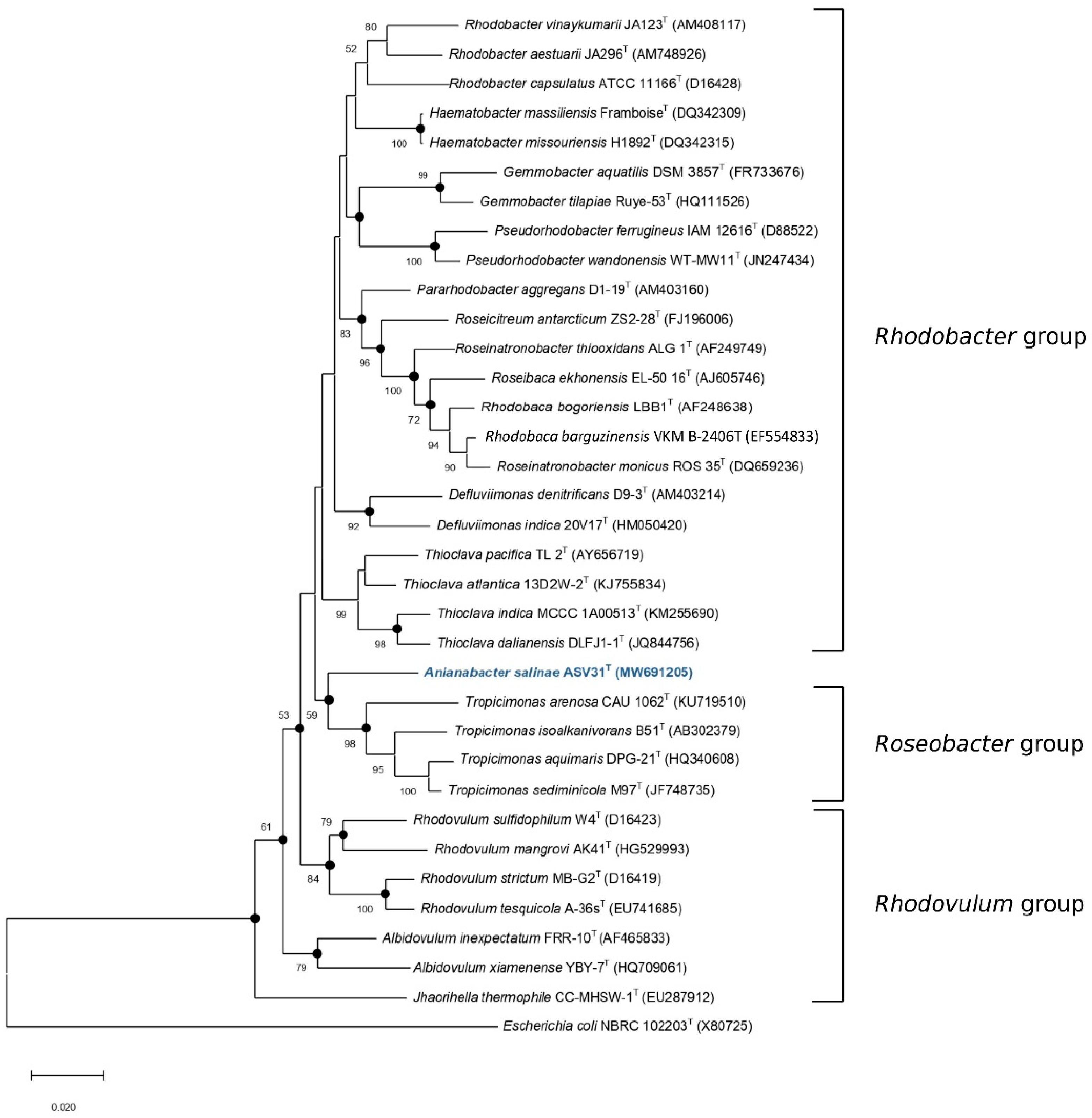
3.1. 16S rRNA Gene-Based Identification and Phylogeny
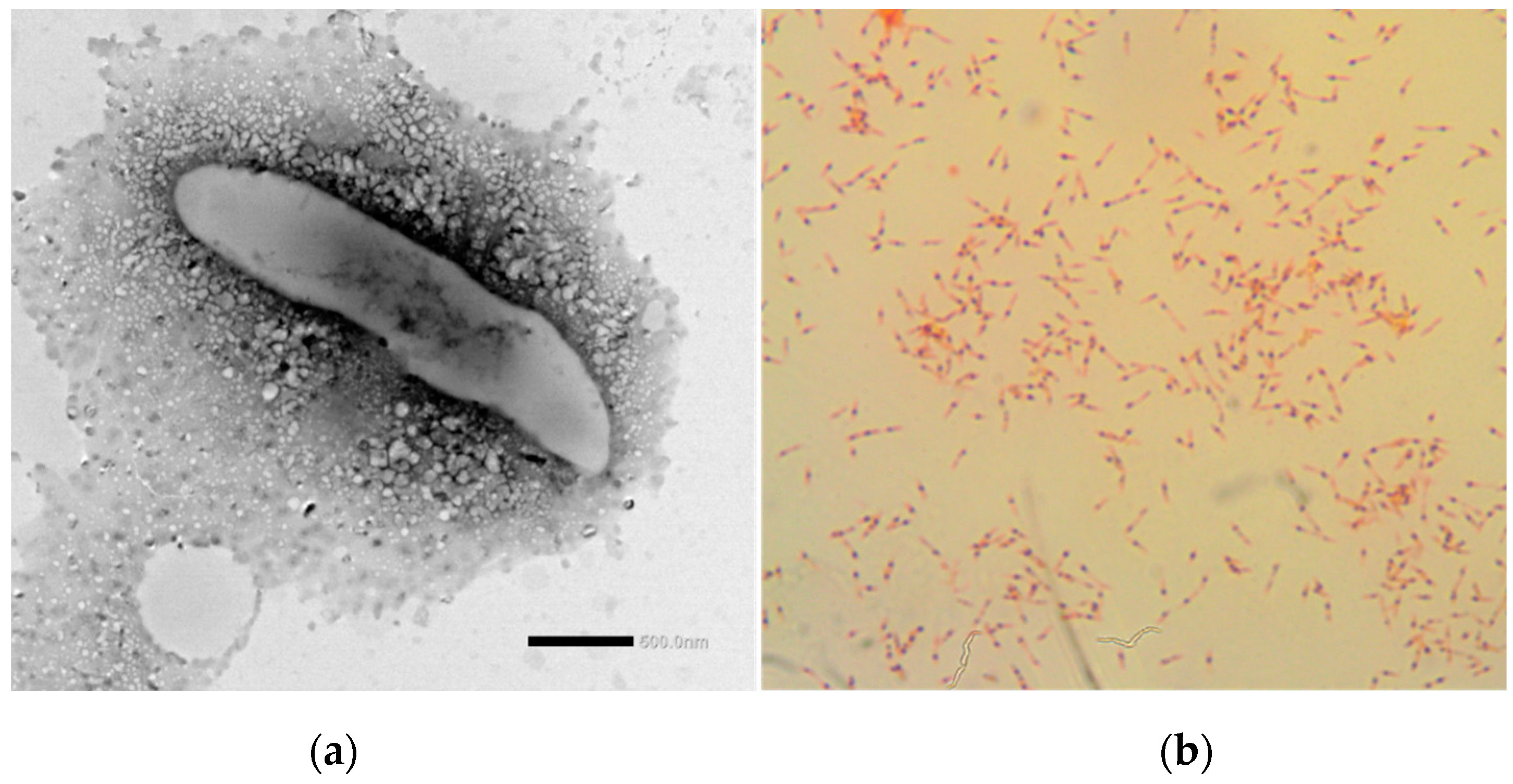
3.2. Phenotypic and Chemotaxonomic Characterisation
3.3. Genome-Based Characterisation and Properties
3.4. Biotechnological Potential of Strain ASV31T for the Production of Secondary Metabolites and Enzymes
4. Conclusions
Description of Anianabacter salinae gen. nov. sp. nov.
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Acknowledgments
Conflicts of Interest
References
- Landa, M.; Montero, A. Salt Valley of Añana. Towarsds Its Full Recovery. Vallée Salée de Añana. Vers Une Récupération Intégrale; Diputación Foral de Álava: Vitoria-Gasteiz, Spain, 2007; ISBN 978-84-7824-680-2. [Google Scholar]
- Garrity, G.M.; Bell, J.A.; Lilburn, T. Family I. Rhodobacteraceae fam. nov. Bergey’s Man. Syst. Bacteriol. 2005, 2, 161. [Google Scholar]
- Hördt, A.; López, M.G.; Meier-Kolthoff, J.P.; Schleuning, M.; Weinhold, L.-M.; Tindall, B.J.; Gronow, S.; Kyrpides, N.C.; Woyke, T.; Göker, M. Analysis of 1,000+ Type-Strain Genomes Substantially Improves Taxonomic Classification of Alphaproteobacteria. Front. Microbiol. 2020, 11, 468. [Google Scholar] [CrossRef] [PubMed]
- Szuróczki, S.; Abbaszade, G.; Buni, D.; Bóka, K.; Schumann, P.; Neumann-Schaal, M.; Vajna, B.; Tóth, E. Fertoeibacter niger gen. nov., sp. nov. a Novel Alkaliphilic Bacterium of the Family Rhodobacteraceae. Int. J. Syst. Evol. Microbiol. 2021, 71, 004762. [Google Scholar] [CrossRef] [PubMed]
- Pujalte, M.J.; Lucena, T.; Ruvira, M.A.; Arahal, D.R.; Macián, M.C. The Family Rhodobacteraceae. In The Prokaryotes; Springer: Berlin/Heidelberg, Germany, 2014; pp. 439–512. [Google Scholar]
- Liang, K.Y.; Orata, F.D.; Boucher, Y.F.; Case, R.J. Roseobacters in a Sea of Poly-and Paraphyly: Whole Genome-Based Taxonomy of the Family Rhodobacteraceae and the Proposal for the Split of the “Roseobacter Clade” into a Novel Family, Roseobacteraceae Fam. Nov. Front. Microbiol. 2021, 12, 1635. [Google Scholar] [CrossRef]
- Barnier, C.; Clerissi, C.; Lami, R.; Intertaglia, L.; Lebaron, P.; Grimaud, R.; Urios, L. Description of Palleronia Rufa sp. nov., a Biofilm-Forming and AHL-Producing Rhodobacteraceae, Reclassification of Hwanghaeicola Aestuarii as Palleronia Aestuarii comb. nov., Maribius Pontilimi as Palleronia Pontilimi comb. nov., Maribius Salinus as Palleronia Salina comb. nov., Maribius Pelagius as Palleronia Pelagia comb. nov. and Emended Description of the Genus Palleronia. Syst. Appl. Microbiol. 2020, 43, 126018. [Google Scholar] [CrossRef]
- Pohlner, M.; Dlugosch, L.; Wemheuer, B.; Mills, H.; Engelen, B.; Reese, B.K. The Majority of Active Rhodobacteraceae in Marine Sediments Belong to Uncultured Genera: A Molecular Approach to Link Their Distribution to Environmental Conditions. Front. Microbiol. 2019, 10, 659. [Google Scholar] [CrossRef]
- Barcelos, M.C.; Vespermann, K.A.; Pelissari, F.M.; Molina, G. Current Status of Biotechnological Production and Applications of Microbial Exopolysaccharides. Crit. Rev. Food Sci. Nutr. 2020, 60, 1475–1495. [Google Scholar] [CrossRef]
- Yin, J.; Chen, J.-C.; Wu, Q.; Chen, G.-Q. Halophiles, Coming Stars for Industrial Biotechnology. Biotechnol. Adv. 2015, 33, 1433–1442. [Google Scholar] [CrossRef]
- Park, S.; Lee, B.; Park, K. Extremophilic Carbohydrate Active Enzymes (CAZymes). J. Nutr. Health Food Eng. 2017, 7, 230–237. [Google Scholar] [CrossRef] [Green Version]
- Weisburg, W.G.; Barns, S.M.; Pelletier, D.A.; Lane, D.J. 16S Ribosomal DNA Amplification for Phylogenetic Study. J. Bacteriol. 1991, 173, 697–703. [Google Scholar] [CrossRef] [Green Version]
- Yoon, S.-H.; Ha, S.-M.; Kwon, S.; Lim, J.; Kim, Y.; Seo, H.; Chun, J. Introducing EzBioCloud: A Taxonomically United Database of 16S RRNA Gene Sequences and Whole-Genome Assemblies. Int. J. Syst. Evol. Microbiol. 2017, 67, 1613. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across Computing Platforms. Mol. Biol. Evol. 2018, 35, 1547. [Google Scholar] [CrossRef] [PubMed]
- Saitou, N.; Nei, M. The Neighbor-Joining Method: A New Method for Reconstructing Phylogenetic Trees. Mol. Biol. Evol. 1987, 4, 406–425. [Google Scholar] [CrossRef] [PubMed]
- Fitch, W.M.; Margoliash, E. Construction of Phylogenetic Trees. Science 1967, 155, 279–284. [Google Scholar] [CrossRef]
- Felsenstein, J. Confidence Limits on Phylogenies: An Approach Using the Bootstrap. Evolution 1985, 39, 783–791. [Google Scholar] [CrossRef]
- Smith, A.C.; Hussey, M.A. Gram Stain Protocols. Am. Soc. Microbiol. 2005, 1, 14. [Google Scholar]
- Zapata, M.; Rodríguez, F.; Garrido, J.L. Separation of Chlorophylls and Carotenoids from Marine Phytoplankton: A New HPLC Method Using a Reversed Phase C8 Column and Pyridine-Containing Mobile Phases. Mar. Ecol. Prog. Ser. 2000, 195, 29–45. [Google Scholar] [CrossRef] [Green Version]
- Seoane, S.; Zapata, M.; Orive, E. Growth Rates and Pigment Patterns of Haptophytes Isolated from Estuarine Waters. J. Sea Res. 2009, 62, 286–294. [Google Scholar] [CrossRef]
- Mesquita, D.P.; Amaral, A.L.; Leal, C.; Oehmen, A.; Reis, M.A.M.; Ferreira, E.C. Polyhydroxyalkanoate Granules Quantification in Mixed Microbial Cultures Using Image Analysis: Sudan Black B versus Nile Blue A Staining. Anal. Chim. Acta 2015, 865, 8–15. [Google Scholar] [CrossRef] [Green Version]
- Santhanam, A.; Sasidharan, S. Microbial Production of Polyhydroxy Alkanotes (PHA) from Alcaligens spp. and Pseudomonas oleovorans Using Different Carbon Sources. Afr. J. Biotechnol. 2010, 9, 3144–3150. [Google Scholar]
- Sasser, M. Identification of Bacteria by Gas Chromatography of Cellular Fatty Acids; MIDI Technical Note 101; MIDI Inc.: Newark, DE, USA, 1990. [Google Scholar]
- MIDI. Sherlock Microbial Identification System Operating Manual; Version 6.1; MIDI Inc.: Newark, DE, USA, 2008. [Google Scholar]
- Bligh, E.G.; Dyer, W.J. A rapid method of total lipid extraction and purification. Can. J. Biochem. Physiol. 1959, 37, 911–917. [Google Scholar] [CrossRef] [PubMed]
- Tindall, B.J.; Sikorski, J.; Smibert, R.M.; Kreig, N.R. Phenotipic characterization and the principlen of comparative systematics. In Methods for General and MOlecular Microbiology, 3rd ed.; Reddy, C.A., Beveridge, T.J., Breznak, J.A., Marluf, G.A., Schmidt, T.M., Snyder, L.R., Eds.; ASM Press: Washington, DC, USA, 2007; pp. 330–393. [Google Scholar]
- Davis, J.J.; Wattam, A.R.; Aziz, R.K.; Brettin, T.; Butler, R.; Butler, R.M.; Chlenski, P.; Conrad, N.; Dickerman, A.; Dietrich, E.M. The PATRIC Bioinformatics Resource Center: Expanding Data and Analysis Capabilities. Nucleic Acids Res. 2020, 48, D606–D612. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Parks, D.H.; Imelfort, M.; Skennerton, C.T.; Hugenholtz, P.; Tyson, G.W. CheckM: Assessing the Quality of Microbial Genomes Recovered from Isolates, Single Cells, and Metagenomes. Genome Res. 2015, 25, 1043–1055. [Google Scholar] [CrossRef] [PubMed]
- Yoon, S.-H.; Ha, S.; Lim, J.; Kwon, S.; Chun, J. A Large-Scale Evaluation of Algorithms to Calculate Average Nucleotide Identity. Antonie Van Leeuwenhoek 2017, 110, 1281–1286. [Google Scholar] [CrossRef] [PubMed]
- Meier-Kolthoff, J.P.; Göker, M. TYGS Is an Automated High-Throughput Platform for State-of-the-Art Genome-Based Taxonomy. Nat. Commun. 2019, 10, 2182. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodriguez-R, L.M.; Konstantinidis, K.T. Bypassing Cultivation to Identify Bacterial Species. Microbe Mag. 2014, 9, 111–118. [Google Scholar] [CrossRef]
- Qin, Q.-L.; Xie, B.-B.; Zhang, X.-Y.; Chen, X.-L.; Zhou, B.-C.; Zhou, J.; Oren, A.; Zhang, Y.-Z. A Proposed Genus Boundary for the Prokaryotes Based on Genomic Insights. J. Bacteriol. 2014, 196, 2210–2215. [Google Scholar] [CrossRef] [Green Version]
- Lefort, V.; Desper, R.; Gascuel, O. FastME 2.0: A Comprehensive, Accurate, and Fast Distance-Based Phylogeny Inference Program. Mol. Biol. Evol. 2015, 32, 2798–2800. [Google Scholar] [CrossRef] [Green Version]
- Aziz, R.K.; Bartels, D.; Best, A.A.; DeJongh, M.; Disz, T.; Edwards, R.A.; Formsma, K.; Gerdes, S.; Glass, E.M.; Kubal, M. The RAST Server: Rapid Annotations Using Subsystems Technology. BMC Genom. 2008, 9, 75. [Google Scholar] [CrossRef] [Green Version]
- Blin, K.; Shaw, S.; Kloosterman, A.M.; Charlop-Powers, Z.; Van Wezel, G.P.; Medema, M.H.; Weber, T. AntiSMASH 6.0: Improving Cluster Detection and Comparison Capabilities. Nucleic Acids Res. 2021, 49, W29–W35. [Google Scholar] [CrossRef]
- Zhang, H.; Yohe, T.; Huang, L.; Entwistle, S.; Wu, P.; Yang, Z.; Busk, P.K.; Xu, Y.; Yin, Y. DbCAN2: A Meta Server for Automated Carbohydrate-Active Enzyme Annotation. Nucleic Acids Res. 2018, 46, W95–W101. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, R.; Lai, Q.; Wang, W.; Li, S.; Shao, Z. Thioclava dalianensis sp. nov., Isolated from Surface Seawater. Int. J. Syst. Evol. Microbiol. 2013, 63, 2981–2985. [Google Scholar] [CrossRef] [PubMed]
- Ramaprasad, E.V.V.; Tushar, L.; Dave, B.; Sasikala, C.; Ramana, C.V. Rhodovulum algae sp. nov., Isolated from an Algal Mat. Int. J. Syst. Evol. Microbiol. 2016, 66, 3367–3371. [Google Scholar] [CrossRef]
- Sorokin, D.Y.; Tourova, T.P.; Spiridonova, E.M.; Rainey, F.A.; Muyzer, G. Thioclava pacifica gen. nov., sp. nov., a Novel Facultatively Autotrophic, Marine, Sulfur-Oxidizing Bacterium from a near-Shore Sulfidic Hydrothermal Area. Int. J. Syst. Evol. Microbiol. 2005, 55, 1069–1075. [Google Scholar] [CrossRef] [Green Version]
- Harwati, T.U.; Kasai, Y.; Kodama, Y.; Susilaningsih, D.; Watanabe, K. Tropicimonas isoalkanivorans gen. nov., sp. nov., a Branched-Alkane-Degrading Bacterium Isolated from Semarang Port in Indonesia. Int. J. Syst. Evol. Microbiol. 2009, 59, 388–391. [Google Scholar] [CrossRef] [Green Version]
- Helsel, L.O.; Hollis, D.; Steigerwalt, A.G.; Morey, R.E.; Jordan, J.; Aye, T.; Radosevic, J.; Jannat-Khah, D.; Thiry, D.; Lonsway, D.R. Identification of “Haematobacter,” a New Genus of Aerobic Gram-Negative Rods Isolated from Clinical Specimens, and Reclassification of Rhodobacter massiliensis as “Haematobacter massiliensis comb. nov.” J. Clin. Microbiol. 2007, 45, 1238–1243. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chun, J.; Oren, A.; Ventosa, A.; Christensen, H.; Arahal, D.R.; da Costa, M.S.; Rooney, A.P.; Yi, H.; Xu, X.-W.; De Meyer, S. Proposed Minimal Standards for the Use of Genome Data for the Taxonomy of Prokaryotes. Int. J. Syst. Evol. Microbiol. 2018, 68, 461–466. [Google Scholar] [CrossRef]
- Wirth, J.S.; Whitman, W.B. Phylogenomic Analyses of a Clade within the Roseobacter Group Suggest Taxonomic Reassignments of Species of the Genera Aestuariivita, Citreicella, Loktanella, Nautella, Pelagibaca, Ruegeria, Thalassobius, Thiobacimonas and Tropicibacter, and the Proposal of Six Novel Genera. Int. J. Syst. Evol. Microbiol. 2018, 68, 2393–2411. [Google Scholar] [CrossRef]
- Suresh, G.; Lodha, T.D.; Indu, B.; Sasikala, C.; Ramana, C.V. Taxogenomics Resolves Conflict in the Genus Rhodobacter: A Two and Half Decades Pending Thought to Reclassify the Genus Rhodobacter. Front. Microbiol. 2019, 10, 2480. [Google Scholar] [CrossRef] [Green Version]
- Orata, F.D.; Meier-Kolthoff, J.P.; Sauvageau, D.; Stein, L.Y. Phylogenomic Analysis of the Gammaproteobacterial Methanotrophs (Order Methylococcales) Calls for the Reclassification of Members at the Genus and Species Levels. Front. Microbiol. 2018, 9, 3162. [Google Scholar] [CrossRef] [Green Version]
- Aliyu, H.; Lebre, P.; Blom, J.; Cowan, D.; De Maayer, P. Phylogenomic Re-Assessment of the Thermophilic Genus Geobacillus. Syst. Appl. Microbiol. 2016, 39, 527–533. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lopes-Santos, L.; Castro, D.B.A.; Ferreira-Tonin, M.; Corrêa, D.B.A.; Weir, B.S.; Park, D.; Ottoboni, L.M.M.; Neto, J.R.; Destéfano, S.A.L. Reassessment of the Taxonomic Position of Burkholderia andropogonis and Description of Robbsia andropogonis gen. nov., comb. nov. Antonie Van Leeuwenhoek 2017, 110, 727–736. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Xue, H.; Sang, S.; Lin, C.; Wang, X. Phylogenetic Analysis of Family Neisseriaceae Based on Genome Sequences and Description of Populibacter corticis gen. nov., sp. nov., a Member of the Family Neisseriaceae, Isolated from Symptomatic Bark of Populus x euramericana Canker. PLoS ONE 2017, 12, e0174506. [Google Scholar] [CrossRef] [PubMed]
- Marsh, W.S.; Heise, B.W.; Krzmarzick, M.J.; Murdoch, R.W.; Fathepure, B.Z. Isolation and Characterization of a Halophilic modicisalibacter sp. Strain Wilcox from Produced Water. Sci. Rep. 2021, 11, 6943. [Google Scholar] [CrossRef] [PubMed]
- Wells, T., Jr.; Ragauskas, A.J. Biotechnological Opportunities with the β-Ketoadipate Pathway. Trends Biotechnol. 2012, 30, 627–637. [Google Scholar] [CrossRef]
- Mahiudddin, M.; Fakhruddin, A.N.M. Degradation of Phenol via Meta Cleavage Pathway by Pseudomonas fluorescens PU1. ISRN Microbiol. 2012, 2012, 1–6. [Google Scholar] [CrossRef] [Green Version]
- Forward, J.A.; Behrendt, M.C.; Wyborn, N.R.; Cross, R.; Kelly, D.J. TRAP Transporters: A New Family of Periplasmic Solute Transport Systems Encoded by the DctPQM Genes of Rhodobacter capsulatus and by Homologs in Diverse Gram-Negative Bacteria. J. Bacteriol. 1997, 179, 5482–5493. [Google Scholar] [CrossRef] [Green Version]
- Rosa, L.T.; Bianconi, M.E.; Thomas, G.H.; Kelly, D.J. Tripartite ATP-Independent Periplasmic (TRAP) Transporters and Tripartite Tricarboxylate Transporters (TTT): From Uptake to Pathogenicity. Front. Cell. Infect. Microbiol. 2018, 8, 33. [Google Scholar] [CrossRef] [Green Version]
- Maeda, S.; Omata, T. Nitrite Transport Activity of the ABC-Type Cyanate Transporter of the Cyanobacterium Synechococcus elongatus. J. Bacteriol. 2009, 191, 3265–3272. [Google Scholar] [CrossRef] [Green Version]
- Bartling, P.; Vollmers, J.; Petersen, J. The First World Swimming Championships of Roseobacters—Phylogenomic Insights into an Exceptional Motility Phenotype. Syst. Appl. Microbiol. 2018, 41, 544–554. [Google Scholar] [CrossRef]
- Czech, L.; Hermann, L.; Stöveken, N.; Richter, A.A.; Höppner, A.; Smits, S.H.J.; Heider, J.; Bremer, E. Role of the Extremolytes Ectoine and Hydroxyectoine as Stress Protectants and Nutrients: Genetics, Phylogenomics, Biochemistry, and Structural Analysis. Genes 2018, 9, 177. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, R.; Wong, H.L.; Kindler, G.S.; MacLeod, F.I.; Benaud, N.; Ferrari, B.C.; Burns, B.P. Discovery of an Abundance of Biosynthetic Gene Clusters in Shark Bay Microbial Mats. Front. Microbiol. 2020, 11, 1950. [Google Scholar] [CrossRef] [PubMed]
- Reddy, G.K.; Leferink, N.G.H.; Umemura, M.; Ahmed, S.T.; Breitling, R.; Scrutton, N.S.; Takano, E. Exploring Novel Bacterial Terpene Synthases. PLoS ONE 2020, 15, e0232220. [Google Scholar] [CrossRef] [PubMed]
- Llamas, I.; Quesada, E.; Martínez-Cánovas, M.J.; Gronquist, M.; Eberhard, A.; Gonzalez, J.E. Quorum Sensing in Halophilic Bacteria: Detection of N-Acyl-Homoserine Lactones in the Exopolysaccharide-Producing Species of Halomonas. Extremophiles 2005, 9, 333–341. [Google Scholar] [CrossRef]
- Harris, L.A.; Saint-Vincent, P.M.B.; Guo, X.; Hudson, G.A.; DiCaprio, A.J.; Zhu, L.; Mitchell, D.A. Reactivity-Based Screening for Citrulline-Containing Natural Products Reveals a Family of Bacterial Peptidyl Arginine Deiminases. ACS Chem. Biol. 2020, 15, 3167–3175. [Google Scholar] [CrossRef] [PubMed]
- Cheng, C.; Hua, Z.-C. Lasso Peptides: Heterologous Production and Potential Medical Application. Front. Bioeng. Biotechnol. 2020, 8, 571165. [Google Scholar] [CrossRef]
- Klaus, T.; Ninck, S.; Albersmeier, A.; Busche, T.; Wibberg, D.; Jiang, J.; Elcheninov, A.G.; Zayulina, K.S.; Kaschani, F.; Bräsen, C.; et al. Activity-Based Protein Profiling for the Identification of Novel Carbohydrate-Active Enzymes Involved in Xylan Degradation in the Hyperthermophilic Euryarchaeon Thermococcus sp. Strain 2319x1E. Front. Microbiol. 2022, 12, 3868. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Characteristics | 1 | 2 | 3 | 4 | 5 |
|---|---|---|---|---|---|
| Cell shape | Rod | Rod | Rod | Rod | Rod |
| Gram staining | - | - | - | - | - |
| Motility | - | + | + | + | - |
| Colony colour | Beige to pink | Beige | Maroon | White | Not pigmented |
| Growth range (optimum): | |||||
| NaCl (% m/v) | 0–23 (3–5) | 1–10 (3–5) | 0–10 (2–5) | 1–6 | NA |
| Temperature (°C) | 18–37 (30) | 8–42 (30) | 4–42 (30) | 10–46 (37) | NA |
| pH | 6.5–9.5 (7–9.5) | 6.0–9.5 (6.0) | 6.0–9.5 (6.0) | 5.5–8.0 | NA |
| Anaerobic growth | - | - | + | - | - |
| Photoorganoheterotrophy | - | - | + | NA | NA |
| Chemolithoautotrophy | - | + | ND | - | NA |
| Photolithoautotrophy | - | - | - | NA | NA |
| Catalase test | + | - | + | - | + |
| Urease test | - | - | + | NA | + |
| Hydrolysis of: | |||||
| Casein | - | - | - | NA | NA |
| Tween 20 | + | - | - | - | NA |
| Tween 80 | - | - | - | - | NA |
| Aesculin | + | + | + | NA | - |
| Assimilation of: | |||||
| Malate | - | - | w | NA | NA |
| Enzymatic activities: | |||||
| Esterase (C 4) | + | w | w | NA | + |
| Esterase lipase (C 8) | + | w | - | NA | + |
| Valine arylamidase | + | - | w | NA | - |
| Acid phosphatase | w | w | + | NA | + |
| Naphthol-AS-BI-phosphohydrolase | w | w | + | NA | w |
| Cystine arylamidase | w | - | - | NA | - |
| α-glucosidase | + | + | - | NA | - |
| β-glucosidase | - | w | - | NA | - |
| Pigments | Spheroidenone, spheroidene and BChl a | Not detected 1,a | Spheroidenone and BChl a 1,a,b | ND | ND |
| Fatty Acid | 1 | 2 | 3 |
|---|---|---|---|
| Saturated: | |||
| C16:0 | 5.33 | 1.71 | 16.22 |
| C17:0 | TR | - | - |
| C18:0 | 1.45 | 3.06 | 3.25 |
| Unsaturated: | |||
| C15:1 ω8c | TR | - | - |
| C20:2 ω6,9c | 1.04 | - | - |
| Branched: | |||
| C18:1 ω7c 11-metilo | 8.13 | - | - |
| C19:0 cyclo ω8c | 38.16 | - | - |
| C19:0 10-metilo | 3.35 | 1.12 | - |
| C16:1 ω7c alcohol | - | - | TR |
| Hydroxy: | |||
| C10:0 3OH | - | 5.14 | 3.24 |
| C12:0 2OH | - | 1.03 | TR |
| C18:0 3OH | - | 4.85 | - |
| Summed Feature 2 1 | 2.11 | - | 1.56 |
| Summed Feature 3 1 | TR | - | 2.97 |
| Summed Feature 7 1 | 1.08 | - | 1.19 |
| Summed Feature 8 1 | 37.34 | 83.08 | 70.12 |
| Strains | Genome Size (Mbp) | DNA G+C Content (%) | ANI (%) | dDDH (%) | AAI (%) | POCP (%) |
|---|---|---|---|---|---|---|
| ASV31T | ASV31T | ASV31T | ASV31T | |||
| Anianabacter salinae ASV31T | 3.6 | 65.7 | - | - | - | - |
| Thioclava pacifica DSM 10166T | 3.7 | 63.9 | 72.8 | 18.9 | 61.3 | 56.0 |
| Rhodovulum algae LMG 29228T | 4.2 | 67.4 | 73.9 | 19.2 | 63.5 | 54.0 |
| Tropicimonas isoalkanivorans B51T | 4.9 | 64.6 | 72.5 | 18.5 | 62.3 | 52.0 |
| Haematobacter massiliensis CCUG 47968T | 4.2 | 64.5 | 72.5 | 19.3 | 60.2 | 50.0 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Azpiazu-Muniozguren, M.; García, M.; Laorden, L.; Martinez-Malaxetxebarria, I.; Seoane, S.; Bikandi, J.; Garaizar, J.; Martínez-Ballesteros, I. Anianabacter salinae gen. nov., sp. nov. ASV31T, a Facultative Alkaliphilic and Extremely Halotolerant Bacterium Isolated from Brine of a Millennial Continental Saltern. Diversity 2022, 14, 1009. https://doi.org/10.3390/d14111009
Azpiazu-Muniozguren M, García M, Laorden L, Martinez-Malaxetxebarria I, Seoane S, Bikandi J, Garaizar J, Martínez-Ballesteros I. Anianabacter salinae gen. nov., sp. nov. ASV31T, a Facultative Alkaliphilic and Extremely Halotolerant Bacterium Isolated from Brine of a Millennial Continental Saltern. Diversity. 2022; 14(11):1009. https://doi.org/10.3390/d14111009
Chicago/Turabian StyleAzpiazu-Muniozguren, Maia, Minerva García, Lorena Laorden, Irati Martinez-Malaxetxebarria, Sergio Seoane, Joseba Bikandi, Javier Garaizar, and Ilargi Martínez-Ballesteros. 2022. "Anianabacter salinae gen. nov., sp. nov. ASV31T, a Facultative Alkaliphilic and Extremely Halotolerant Bacterium Isolated from Brine of a Millennial Continental Saltern" Diversity 14, no. 11: 1009. https://doi.org/10.3390/d14111009