
Effect of Different Altitudes on Morpho-Physiological Attributes Associated with Mango Quality

Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material and Growth Conditions
2.2. Sample Preparation and Data Collection
2.3. Identification and Characterization of Volatile Compounds
2.4. Data Analysis
3. Results
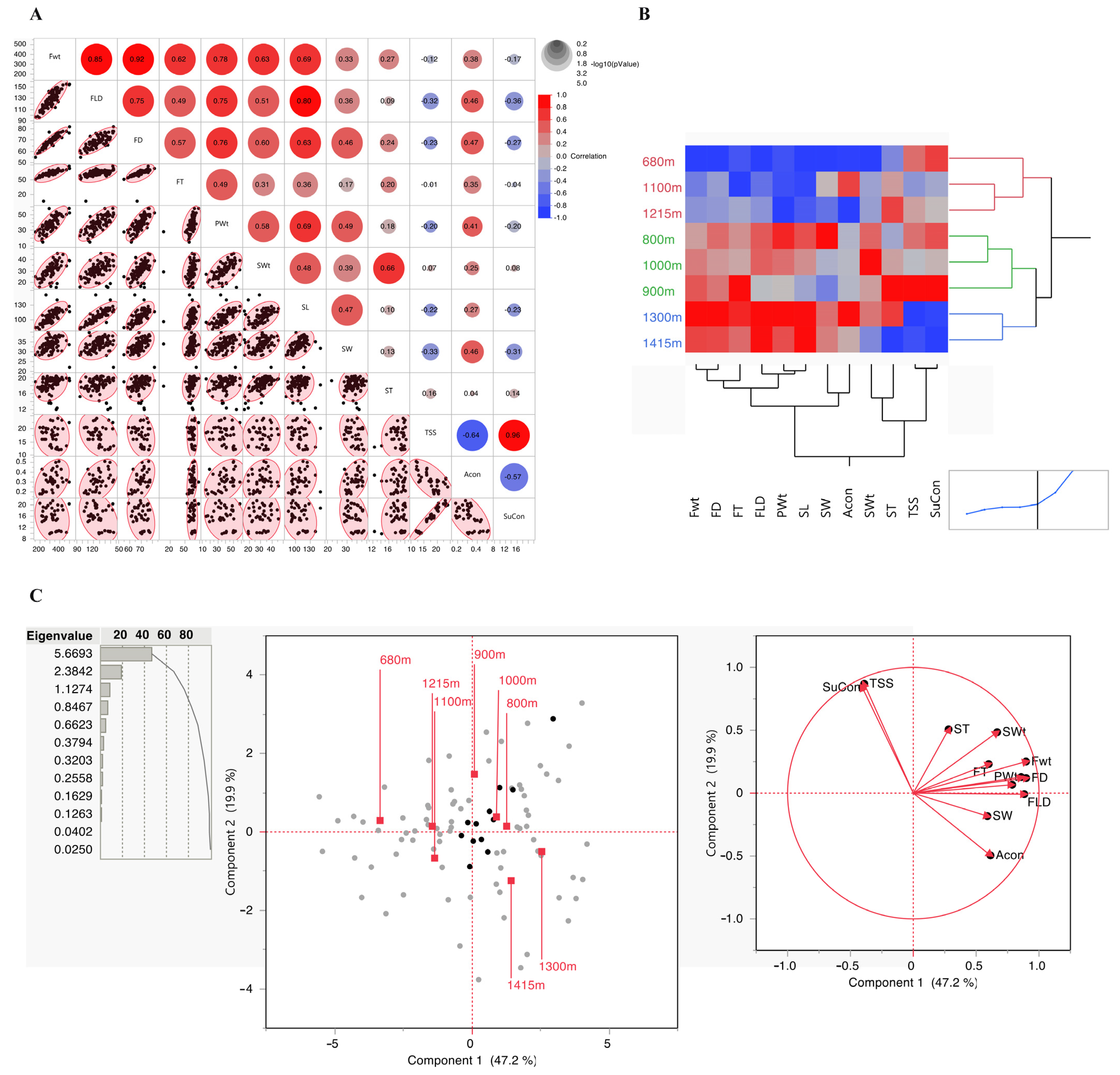
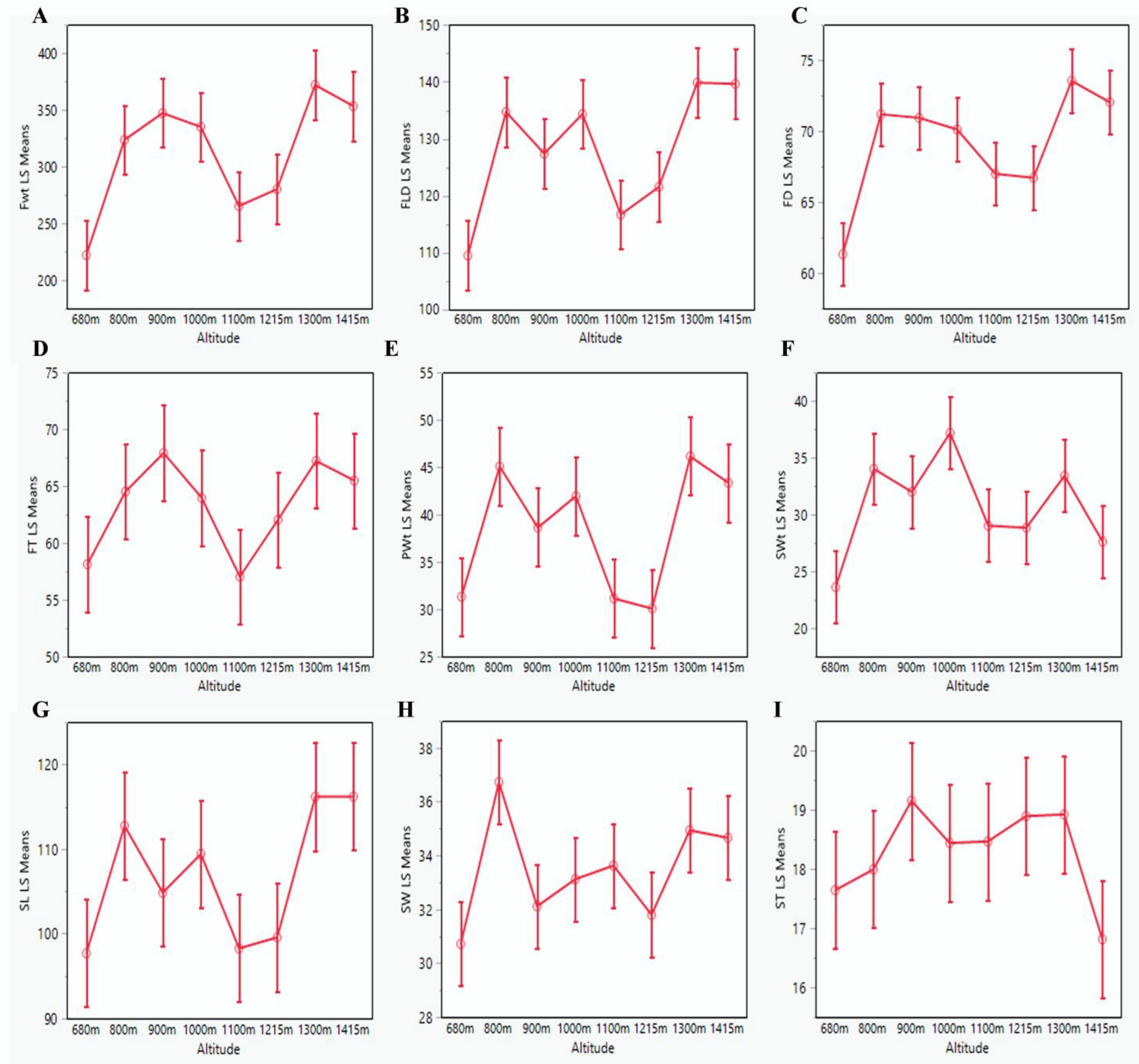
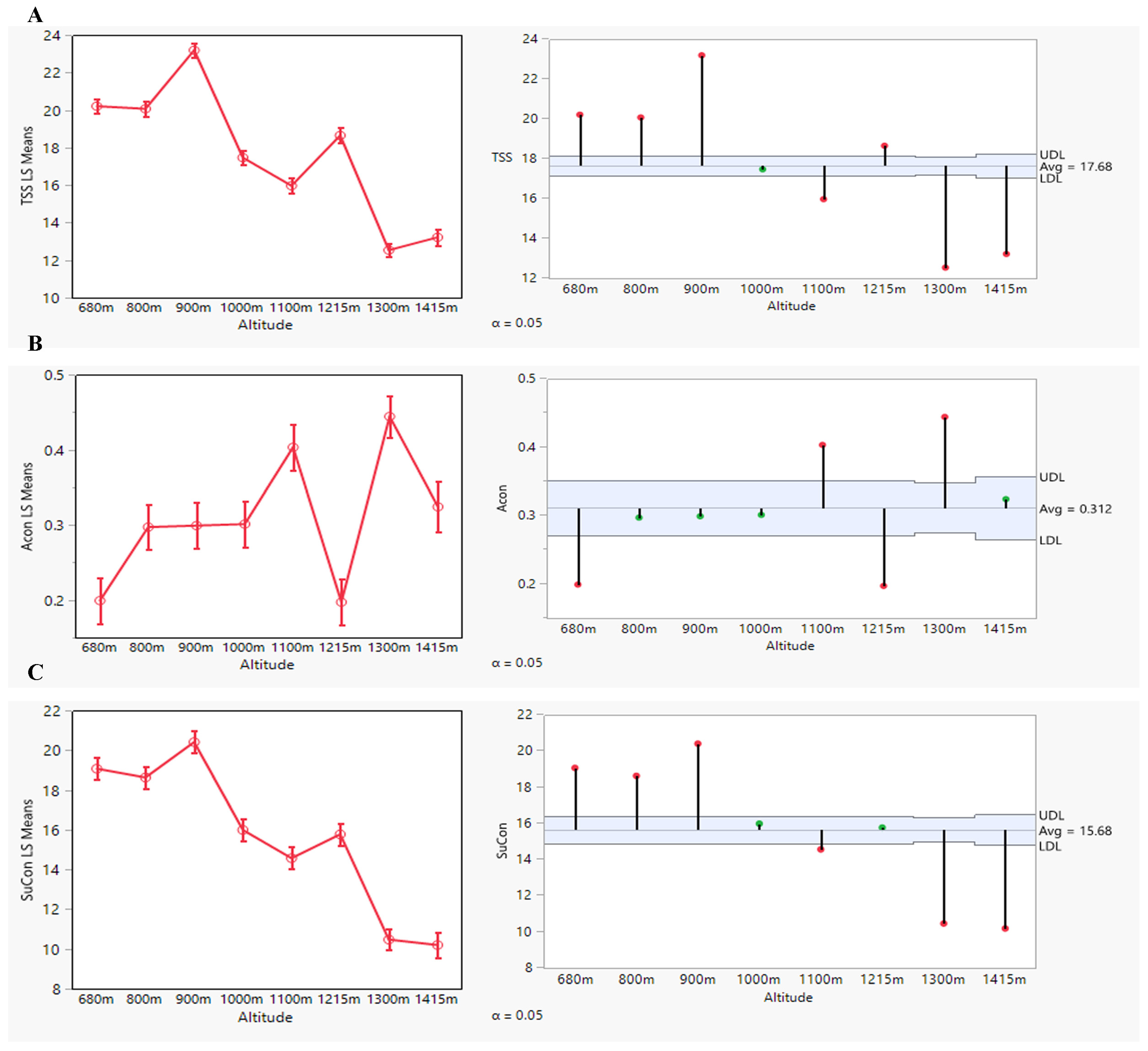
3.1. Phenotypic Characterization
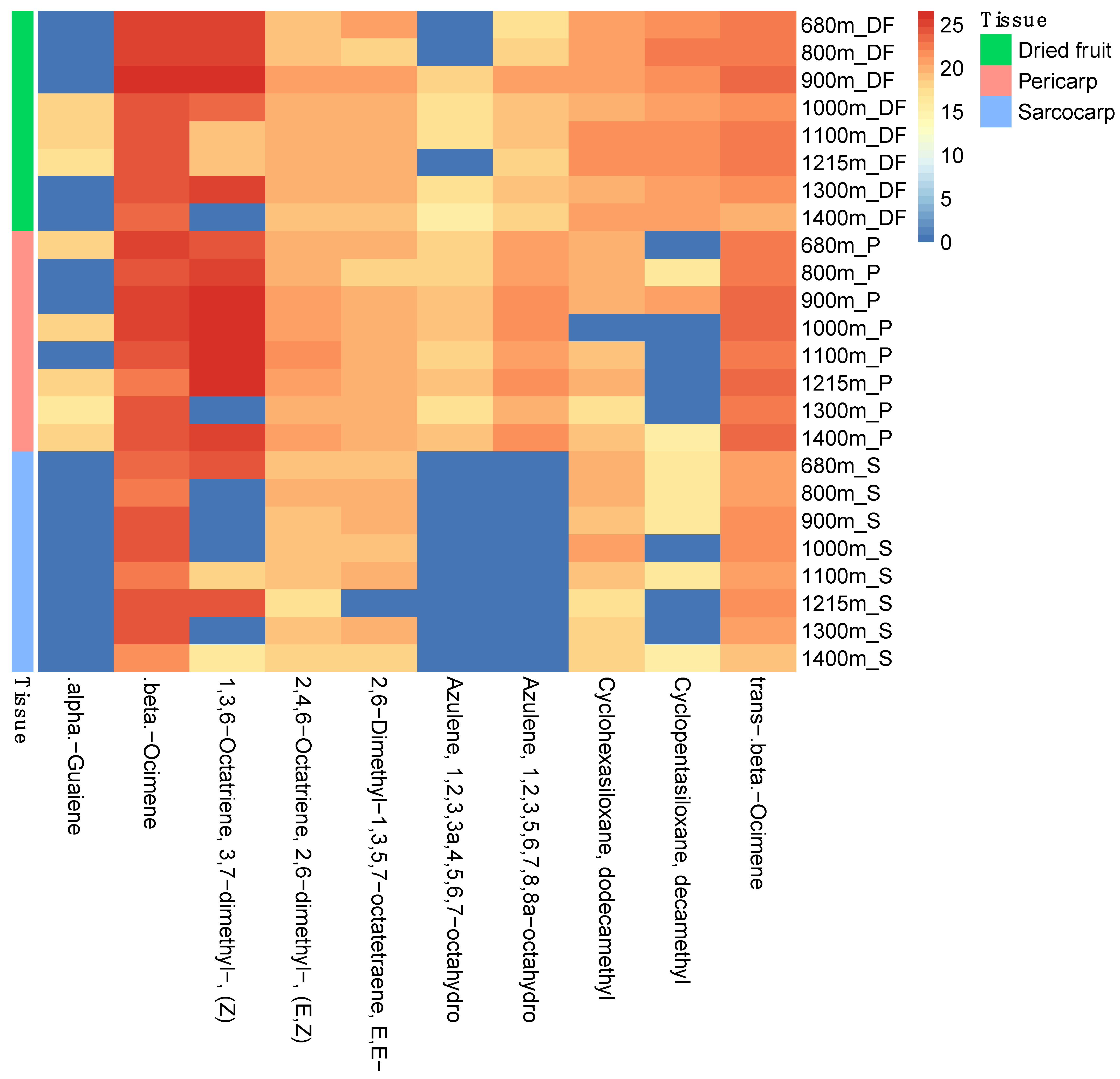
3.2. Characterization of Volatile Compounds
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zhang, C.; Xie, D.; Bai, T.; Luo, X.; Zhang, F.; Ni, Z.; Chen, Y. Diversity of a large collection of natural populations of mango (Mangifera indica Linn.) revealed by agro-morphological and quality traits. Diversity 2020, 12, 27. [Google Scholar] [CrossRef] [Green Version]
- Thakor, N. Indian mango–production and export scenario. Adv. Agric. Res. Technol. J. 2019, 3, 80–88. [Google Scholar]
- Salami, A.; Dalibi, J. An Evaluation of the Effects of Local Climates on the Horticultural Improvement Techniques of Mangifera Indica (L.) in a Dry-Land Ecosystem of Nigeria. J. Innov. Res. Adv. Stud. (IJIRAS) 2019, 6, 18–23. [Google Scholar]
- Orwa, C.; Mutua, A.; Kindt, R.; Jamnadass, R.; Simons, A. Agroforestree Database: A Tree Reference and Selection Guide; Version 4; World Agroforestry Centre: Nairobi, Kenya, 2009. [Google Scholar]
- Zhang, D.; Chong, W.; LI, X.-L. Yield gap and production constraints of mango (Mangifera indica) cropping systems in Tianyang County, China. J. Integr. Agric. 2019, 18, 1726–1736. [Google Scholar] [CrossRef]
- Gao, A.; Chen, Y.; Luo, R.; Huang, J.; Zhao, Z.; Wang, W.; Wang, Y.; Dang, Z. Development status of Chinese mango industry in 2018. Adv. Agric. Hortic. Entomol. 2019, 1, 21–60. [Google Scholar]
- Bhagwan, A. Mango cultivation practices in the tropics: Good agricultural practices to maximize sustainable yields Sisir Mitra, International Society for Horticultural Science, India. In Achieving Sustainable Cultivation of Mangoes; Burleigh Dodds Science Publishing: Cambridgeshire, UK, 2018; pp. 169–184. [Google Scholar]
- Ramírez, F.; Davenport, T.L.; Fischer, G.; Pinzón, J.C.A.; Ulrichs, C. Mango trees have no distinct phenology: The case of mangoes in the tropics. Sci. Hortic. 2014, 168, 258–266. [Google Scholar] [CrossRef]
- Kobra, K.; Hossain, M.; Talukder, M.; Bhuyan, M. Performance of twelve mango cultivars grown in different agroecological zones of Bangladesh. Bangladesh J. Agric. Res. 2012, 37, 691–710. [Google Scholar] [CrossRef] [Green Version]
- Bally, I.S. Mangifera indica (mango). In Species Profiles for Pacific Island Agroforestry; Permanent Agriculture Resources (PAR): Holualoa, HI, USA, 2006; pp. 1–25. [Google Scholar]
- Subedi, S. Climate change effects of Nepalese fruit production. Adv. Plants Agric. Res. 2019, 9, 141–145. [Google Scholar]
- Das, S.; Datta, M.; Ray, P.; Singh, S.; Jena, R.; Das, B.; Ray, S. Mango (Mangifera indica) Cultivation in North-Eastern Region of India. Adv. Agric. Res. Technol. J. 2019, 3, 54–66. [Google Scholar]
- Wu, H.; Wang, S.; Ma, X.; Ma, W.; Zhan, R.; Yao, Q. Effect of bagging on fruit quality in mango. In Proceedings of the IX International Mango Symposium 992, Sanya, China, 8–12 April 2010; pp. 587–592. [Google Scholar]
- Davenport, T. Reproductive physiology. In The Mango: Botany Production and Uses; CAB International: Wallingford, UK, 2009; pp. 97–169. [Google Scholar]
- Tharanathan, R.; Yashoda, H.; Prabha, T. Mango (Mangifera indica L.), “The king of fruits”—An overview. Food Rev. Int. 2006, 22, 95–123. [Google Scholar] [CrossRef]
- Pott, D.M.; Vallarino, J.G.; Osorio, S. Metabolite changes during postharvest storage: Effects on fruit quality traits. Metabolites 2020, 10, 187. [Google Scholar] [CrossRef] [PubMed]
- Vasco, C.; Avila, J.; Ruales, J.; Svanberg, U.; Kamal-Eldin, A. Physical and chemical characteristics of golden-yellow and purple-red varieties of tamarillo fruit (Solanum betaceum Cav.). Int. J. Food Sci. Nutr. 2009, 60 (Suppl. S7), 278–288. [Google Scholar] [CrossRef] [PubMed]
- Lebaka, V.R.; Wee, Y.-J.; Ye, W.; Korivi, M. Nutritional composition and bioactive compounds in three different parts of mango fruit. Int. J. Environ. Res. Public Health 2021, 18, 741. [Google Scholar] [CrossRef] [PubMed]
- Bhatt, R.; Hossain, A. Concept and consequence of evapotranspiration for sustainable crop production in the era of climate change. In Advanced Evapotranspiration Methods and Applications; IntechOpen: London, UK, 2019; Volume 1, pp. 1–13. [Google Scholar]
- Kamala Jayanthi, P.D.; Kempraj, V.; Aurade, R.M.; Venkataramanappa, R.K.; Nandagopal, B.; Verghese, A.; Bruce, T.J. Specific volatile compounds from mango elicit oviposition in gravid Bactrocera dorsalis females. J. Chem. Ecol. 2014, 40, 259–266. [Google Scholar] [CrossRef] [PubMed]
- Sakho, M.; Chassagne, D.; Crouzet, J. African mango glycosidically bound volatile compounds. J. Agric. Food Chem. 1997, 45, 883–888. [Google Scholar] [CrossRef]
- Pino, J.A.; Mesa, J.; Muñoz, Y.; Martí, M.P.; Marbot, R. Volatile components from mango (Mangifera indica L.) cultivars. J. Agric. Food Chem. 2005, 53, 2213–2223. [Google Scholar] [CrossRef]
- Rojas, C.M.; Senthil-Kumar, M.; Tzin, V.; Mysore, K.S. Regulation of primary plant metabolism during plant-pathogen interactions and its contribution to plant defense. Front. Plant Sci. 2014, 5, 17. [Google Scholar] [CrossRef] [Green Version]
- Sarkar, A.; Hore, M.; Chakraborty, K.; Roy, S. Phytochemical screening and determination of antioxidant activity of Lakhanbhog and Langra mango of Malda district, West Bengal. Pharma Innov. J. 2018, 18, 511–515. [Google Scholar]
- Agneswari, S.; Amutha, S.; Ariharan, V. Comparative evaluation of antioxidant and antimicrobial activity of Dendrophthoe falcata leaves collected from two different host plants. Int. J. Bot. Stud. 2021, 6, 13–18. [Google Scholar]
- Dar, M.S.; Oak, P.; Chidley, H.; Deshpande, A.; Giri, A.; Gupta, V. Nutrient and flavor content of mango (Mangifera indica L.) cultivars: An appurtenance to the list of staple foods. In Nutritional Composition of Fruit Cultivars; Elsevier: Amsterdam, The Netherlands, 2016; pp. 445–467. [Google Scholar]
- Rajwana, I.A.; Khan, I.A.; Malik, A.U.; Saleem, B.A.; Khan, A.S.; Ziaf, K.; Anwar, R.; Amin, M. Morphological and biochemical markers for varietal characterization and quality assessment of potential indigenous mango (Mangifera indica) germplasm. Int. J. Agric. Biol. 2011, 13, 151–158. [Google Scholar]
- Begum, H.; Reddy, M.T.; Malathi, S.; Reddy, B.P.; Narshimulu, G.; Nagaraju, J.; Siddiq, E.A. Morphological and microsatellite analysis of intravarietal heterogeneity in ‘Beneshan’mango (Mangifera indica L.). Int. J. Agric. Food Res. 2013, 1, 1–18. [Google Scholar]
- Bhamini, K.; Jaiswal, U.; Nayyer, M.F.A.A. Studies on the physio-morphology, flowering and fruit characteristics of mango genotypes in subtropical climate of Eastern India. J. Pharmacogn. Phytochem. 2018, 7, 3783–3789. [Google Scholar]
- Bhamini, K.; Kumar, A.; Jaiswal, U.; Ahmad, F.; Rani, R. Morphological characterization of mango (Mangifera indica L.) germplasm using DUS testing. Int. J. Curr. Microbiol. App. Sci 2018, 7, 2944–2959. [Google Scholar] [CrossRef]
- Gill, M.S.; Gill, P. Characterization of sucking type mango genotypes under sub-tropics of Punjab. Indian J. Hortic. 2014, 71, 6–11. [Google Scholar]
- Mohammed, Z.M.A. Morphological and Genetical Diversity of Mango (Mangifera indica L.) Cultivars in Shendi Area. Extensive J. Appl. Sci. 2015, 3, 219–224. [Google Scholar]
- Makroo, H.; Prabhakar, P.K.; Rastogi, N.; Srivastava, B. Characterization of mango puree based on total soluble solids and acid content: Effect on physico-chemical, rheological, thermal and ohmic heating behavior. LWT 2019, 103, 316–324. [Google Scholar] [CrossRef]
- Hocking, R.R. Methods and Applications of Linear Models: Regression and the Analysis of Variance; John Wiley & Sons: Hoboken, NJ, USA, 2013. [Google Scholar]
- FAO. Major Tropical Fruits—Preliminary Market Results 2019; FAO: Rome, Italy, 2020. [Google Scholar]
- Kandegama, W.; Rathnayake, R.M.P.J.; Baig, M.B.; Behnassi, M. Impacts of Climate Change on Horticultural Crop Production in Sri Lanka and the Potential of Climate-Smart Agriculture in Enhancing Food Security and Resilience. In Food Security and Climate-Smart Food Systems; Springer: New York, NY, USA, 2022; pp. 67–97. [Google Scholar]
- Bhattacharyya, T.; Haldankar, P.; Haldavanekar, P.; Burondkar, M.; Salvi, B.; Chakurkar, E. Impact of Climate Change on Horticulture in Konkan, Maharashtra: Activities and Strategies. Indian J. Fertil. 2021, 17, 258–273. [Google Scholar]
- Saqib, M.; Anjum, M.A.; Ali, M.; Ahmad, R.; Sohail, M.; Zakir, I.; Ahmad, S.; Hussain, S. Horticultural Crops as Affected by Climate Change. In Building Climate Resilience in Agriculture; Springer: New York, NY, USA, 2022; pp. 95–109. [Google Scholar]
- Shafqat, W.; Naqvi, S.A.; Maqbool, R.; Haider, M.S.; Jaskani, M.J.; Khan, I.A. Climate change and citrus. In Citrus-Research, Development and Biotechnology; Khan, M.S., Khan, I.A., Eds.; IntechOpen: London, UK, 2021; p. 147. [Google Scholar]
- Halder, S.; Hasan, M.A. Climate change and mango production. Chem. Sci. Rev. Lett. 2020, 9, cs122050121. [Google Scholar] [CrossRef]
- Zainuri, Z.; Sjah, T.; Nurrachman, N.; Ayu, C. Mango off-season technology (MOST): Innovative, applicable, adaptive to climate change, and brings many positive impacts. AIP Conf. Proc. 2019, 2199, 040008. [Google Scholar]
- Wubshet, T.; Wang, Z.; Yang, J.; Chen, H.; Schaefer, D.; Goldberg, S.; Mortimer, P.; Lu, P.; Xu, J. Effect of elevation on photosynthesis of young mango (Mangifera indica L.) trees. Photosynthetica 2021, 59, 508–516. [Google Scholar] [CrossRef]
- Mo, J.; Zhao, G.; Li, Q.; Solangi, G.S.; Tang, L.; Guo, T.; Huang, S.; Hsiang, T. Identification and characterization of Colletotrichum species associated with mango anthracnose in Guangxi, China. Plant Dis. 2018, 102, 1283–1289. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Q.; Bu, J.; Shu, J.; Yu, Z.; Tang, L.; Huang, S.; Guo, T.; Mo, J.; Luo, S.; Solangi, G.S. Colletotrichum species associated with mango in southern China. Sci. Rep. 2019, 9, 18891. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Urban, L.; Le Roux, X.; Sinoquet, H.; Jaffuel, S.; Jannoyer, M. A biochemical model of photosynthesis for mango leaves: Evidence for the effect of fruit on photosynthetic capacity of nearby leaves. Tree Physiol. 2003, 23, 289–300. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bindi, M.; Fibbi, L.; Miglietta, F. Free Air CO2 Enrichment (FACE) of grapevine (Vitis vinifera L.): II. Growth and quality of grape and wine in response to elevated CO2 concentrations. Eur. J. Agron. 2001, 14, 145–155. [Google Scholar] [CrossRef]
- Beckles, D.M. Factors affecting the postharvest soluble solids and sugar content of tomato (Solanum lycopersicum L.) fruit. Postharvest Biol. Technol. 2012, 63, 129–140. [Google Scholar] [CrossRef]
- Welles, G.; Buitelaar, K. Factors affecting soluble solids content of muskmelon (Cucumis melo L.). Neth. J. Agric. Sci. 1988, 36, 239–246. [Google Scholar] [CrossRef]
- Nerd, A.; Gutman, F.; Mizrahi, Y. Ripening and postharvest behaviour of fruits of two Hylocereus species (Cactaceae). Postharvest Biol. Technol. 1999, 17, 39–45. [Google Scholar] [CrossRef]
- Shafique, M.; Ibrahim, M.; Helali, M.; Biswas, S. Studies on the physiological and biochemical composition of different mango cultivars at various maturity levels. Bangladesh J. Sci. Ind. Res. 2006, 41, 101–108. [Google Scholar] [CrossRef]
- Mattheis, J.P.; Fellman, J.K. Preharvest factors influencing flavor of fresh fruit and vegetables. Postharvest Biol. Technol. 1999, 15, 227–232. [Google Scholar] [CrossRef]
- Bertin, N.; Génard, M. Tomato quality as influenced by preharvest factors. Sci. Hortic. 2018, 233, 264–276. [Google Scholar] [CrossRef]
- Zhang, Y.; Hu, W.; Peng, X.; Sun, B.; Wang, X.; Tang, H. Characterization of anthocyanin and proanthocyanidin biosynthesis in two strawberry genotypes during fruit development in response to different light qualities. J. Photochem. Photobiol. B Biol. 2018, 186, 225–231. [Google Scholar] [CrossRef] [PubMed]
- Keller, M. Managing grapevines to optimise fruit development in a challenging environment: A climate change primer for viticulturists. Aust. J. Grape Wine Res. 2010, 16, 56–69. [Google Scholar] [CrossRef]
- Tyagi, S.; Sahay, S.; Imran, M.; Rashmi, K.; Mahesh, S.S. Pre-harvest factors influencing the postharvest quality of fruits: A review. Curr. J. Appl. Sci. Technol 2017, 23, 1–12. [Google Scholar] [CrossRef]
- Garrido, G.; González, D.; Romay, C.; Núñez-Sellés, A.J.; Delgado, R. Scavenger effect of a mango (Mangifera indica L.) food supplement’s active ingredient on free radicals produced by human polymorphonuclear cells and hypoxanthine–xanthine oxidase chemiluminescence systems. Food Chem. 2008, 107, 1008–1014. [Google Scholar] [CrossRef]
- Nunez Selles, A.J.; Aguero Aguero, J.; Paz, L.N. GC-MS analysis of mango stem bark extracts (Mangifera indica L.), Haden variety. Possible contribution of volatile compounds to its health effects. Open Chem. 2021, 19, 27–38. [Google Scholar] [CrossRef]
- Mattivi, F. Key enzymes behind black pepper aroma in wines. J. Exp. Bot. 2016, 67, 555–557. [Google Scholar] [CrossRef] [Green Version]
- Feng, Y.; Guo, X.; Luo, J. GC-MS analysis of volatile oil of Herba Pogostemonis collected from Leizhou county. J. Chin. Med. Mater. 1999, 22, 241–243. [Google Scholar]
- Zakaria, S.R.; Saim, N.; Osman, R.; Abdul Haiyee, Z.; Juahir, H. Combination of sensory, chromatographic, and chemometrics analysis of volatile organic compounds for the discrimination of authentic and unauthentic Harumanis mangoes. Molecules 2018, 23, 2365. [Google Scholar] [CrossRef] [Green Version]
- Zakaria, S.R.; Tajuddin, R.; Osman, R.; Saim, N.; Saaid, M. Optimization of headspace solid phase microextraction (HS-SPME) for the extraction of volatile organic compounds (VOCs) in Mangoes (Harumanis cv.) using 2 stages multivariate analysis. Pertanika J. Sci. Technol. 2017, 25, 167–174. [Google Scholar]
- An, M.-R.; Keum, Y.-S.; Lee, S.-K. Analysis of Volatile Flavor Compounds in Jeju Apple Mango by Using Different Extraction Methods. J. Korean Soc. Food Sci. Nutr. 2015, 44, 775–783. [Google Scholar] [CrossRef] [Green Version]
- Janzantti, N.S.; Macoris, M.S.; Garruti, D.S.; Monteiro, M. Influence of the cultivation system in the aroma of the volatile compounds and total antioxidant activity of passion fruit. LWT-Food Sci. Technol. 2012, 46, 511–518. [Google Scholar] [CrossRef]
- Lapsongphol, S.; Mahayothee, B.; Phupaichitkun, S.; Leis, H.; Haewsungcharoen, M.; Janjai, S.; Mueller, J. Effect of drying temperature on changes in volatile compounds of longan (Dimocarpus longan Lour.) fruit. In Proceedings of the Book of abstracts of the conference on International Agricultural Research for Development, Tropentag, Witzenhausen, Germany, 9 October 2007. [Google Scholar]
- Jaleel, W.; Li, Q.; Shi, Q.; Qi, G.; Latif, M.; Ali, S.; Yasin, N.; Lyu, L.; He, Y. Using GCMS to find out the volatile components in the aroma of three different commercial fruits in China. JAPS J. Anim. Plant Sci. 2021, 31, 166–174. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sources | Fwt | FLD | FD | FT | Pwt | Swt | SL | SW | ST | TSS | Acon | SuCon |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Altitudes | 29,033 | 1380.2 | 169.24 | 173.79 | 492.05 | 200.71 | 677.06 | 41.647 | 6.66 | 67.165 | 0.039 | 71.810 |
| Error | 2567 | 104.15 | 13.704 | 48.521 | 47.346 | 27.761 | 112.17 | 6.7952 | 2.72 | 0.1854 | 0.001 | 0.3819 |
| Signifi. | ** | ** | ** | ** | ** | ** | ** | ** | * | ** | ** | ** |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, C.; Yi, H.; Gao, X.; Bai, T.; Ni, Z.; Chen, Y.; Wang, M.; Zhang, Y.; Pan, J.; Yu, W.; et al. Effect of Different Altitudes on Morpho-Physiological Attributes Associated with Mango Quality. Diversity 2022, 14, 876. https://doi.org/10.3390/d14100876
Zhang C, Yi H, Gao X, Bai T, Ni Z, Chen Y, Wang M, Zhang Y, Pan J, Yu W, et al. Effect of Different Altitudes on Morpho-Physiological Attributes Associated with Mango Quality. Diversity. 2022; 14(10):876. https://doi.org/10.3390/d14100876
Chicago/Turabian StyleZhang, Cuixian, Huaifeng Yi, Xianyu Gao, Tianqi Bai, Zhangguang Ni, Yufu Chen, Meicun Wang, Yong Zhang, Jihong Pan, Weilin Yu, and et al. 2022. "Effect of Different Altitudes on Morpho-Physiological Attributes Associated with Mango Quality" Diversity 14, no. 10: 876. https://doi.org/10.3390/d14100876