
Waterbodies in the Floodplain of the Drava River Host Species-Rich Macrophyte Communities despite Elodea Invasions

Abstract
:1. Introduction
2. Materials and Methods
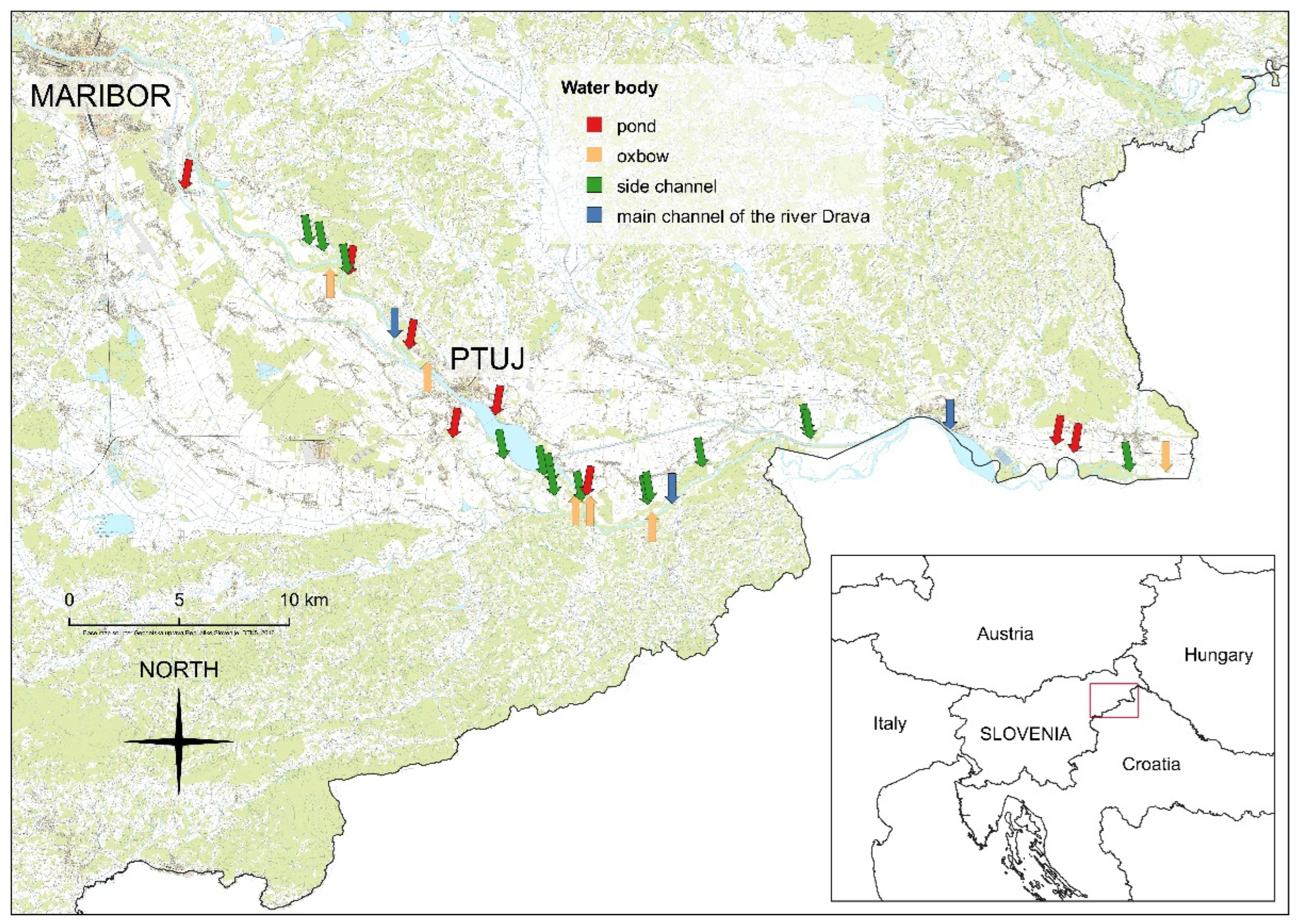
2.1. Study Area
2.2. Macrophyte Data Set
2.3. Environmental Parameters
2.4. Statistical Analyses
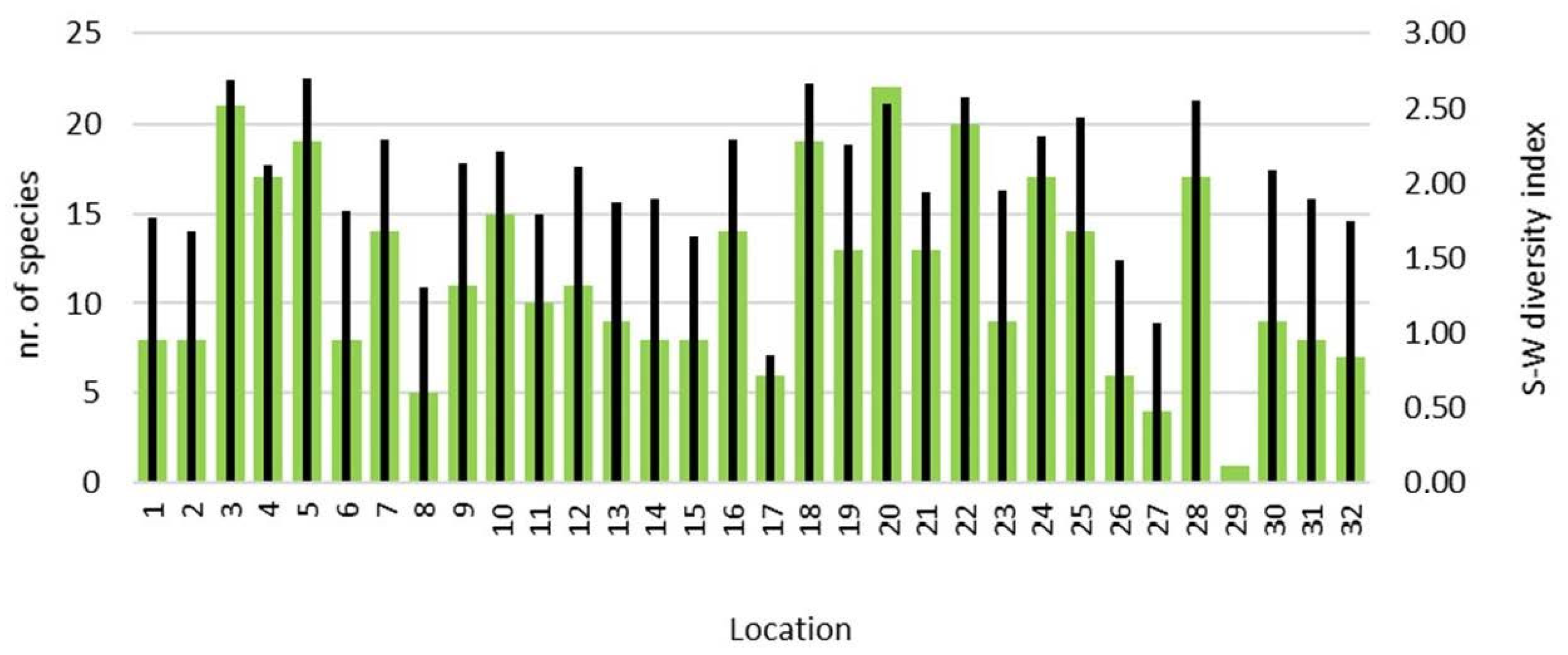
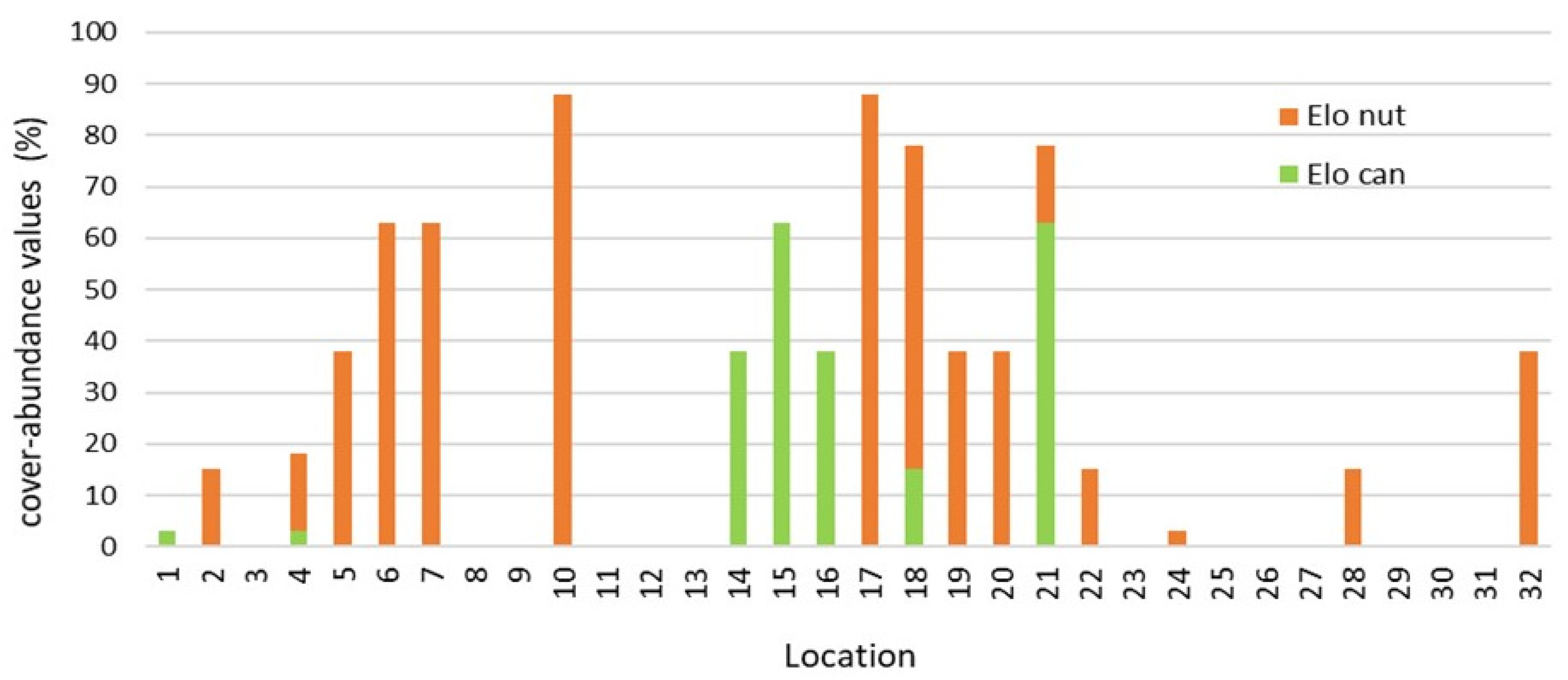
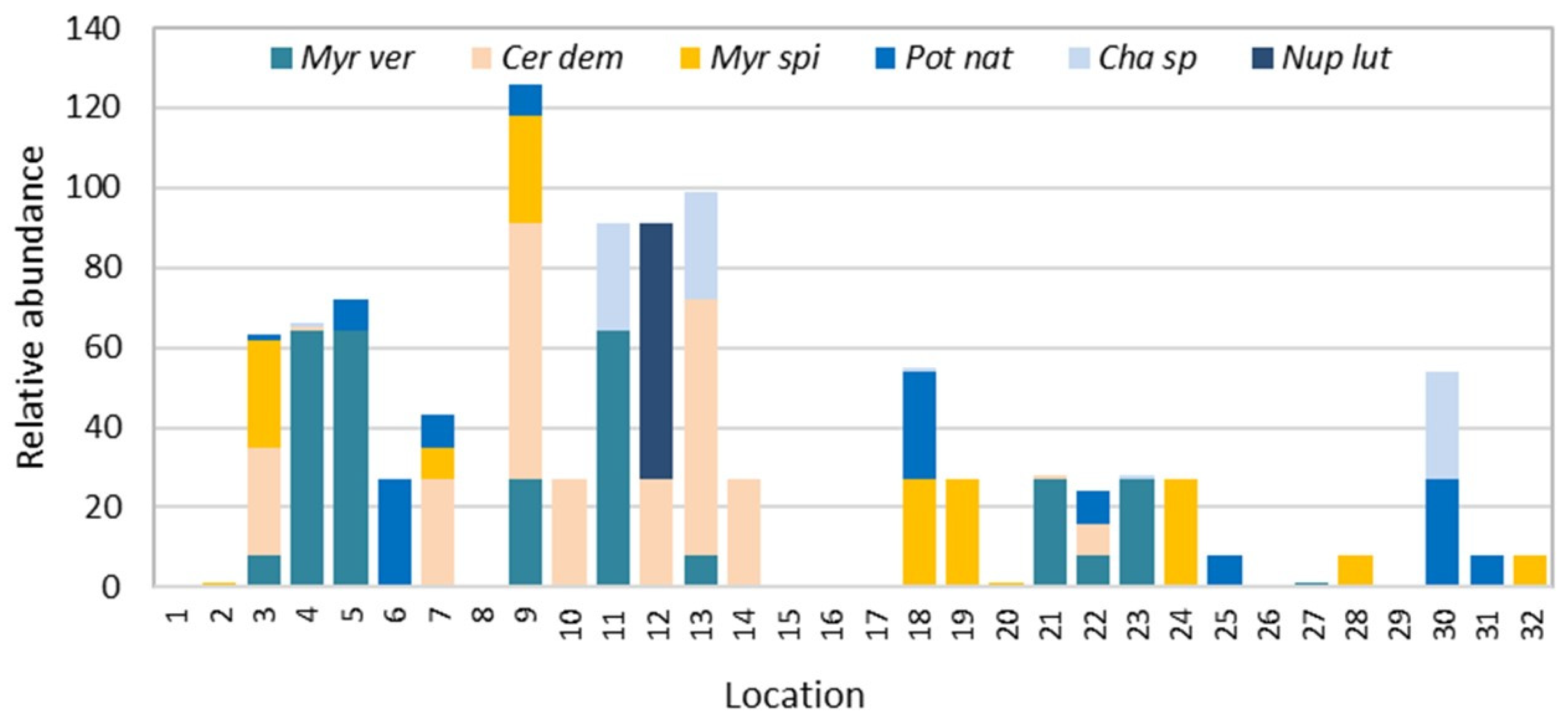
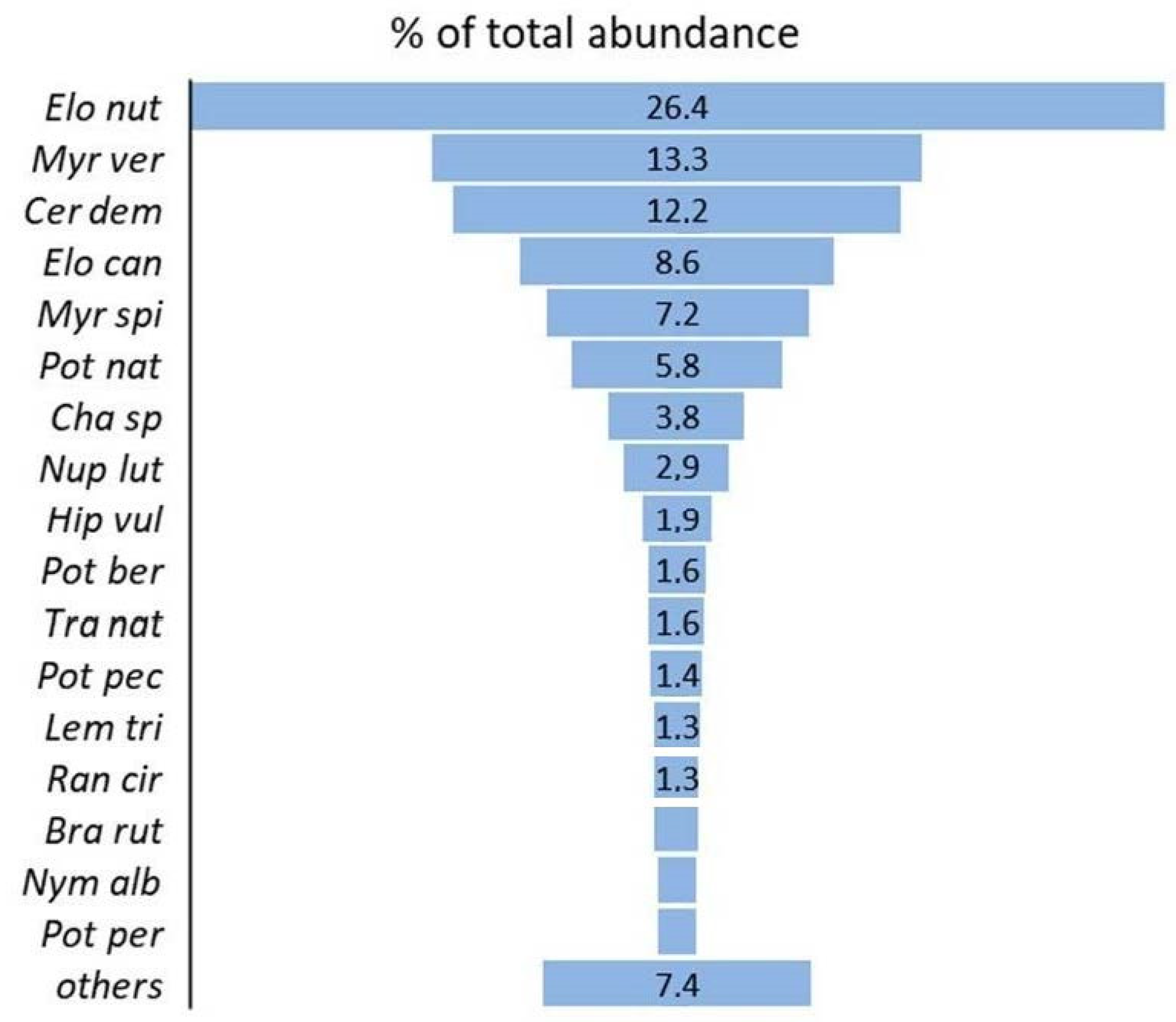
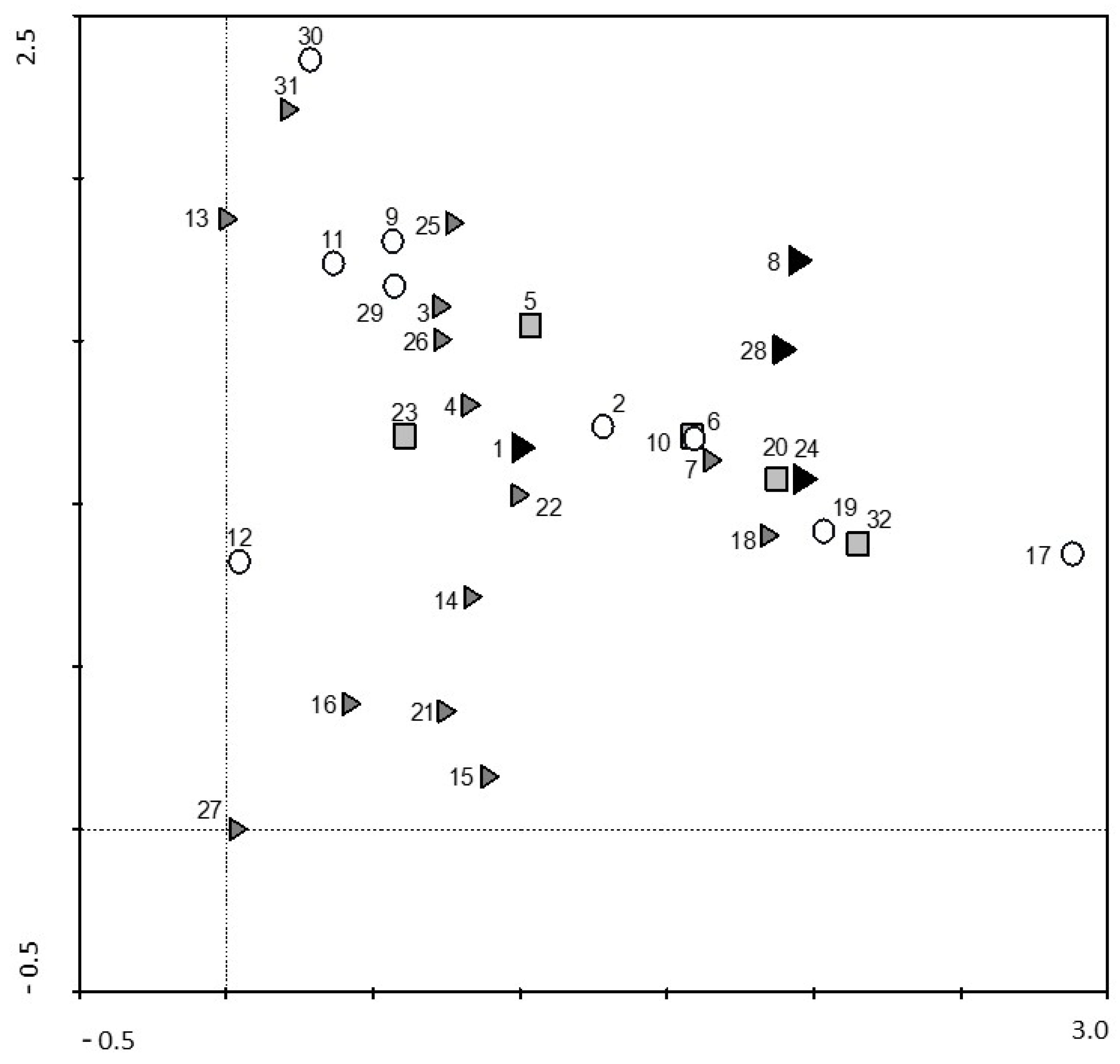
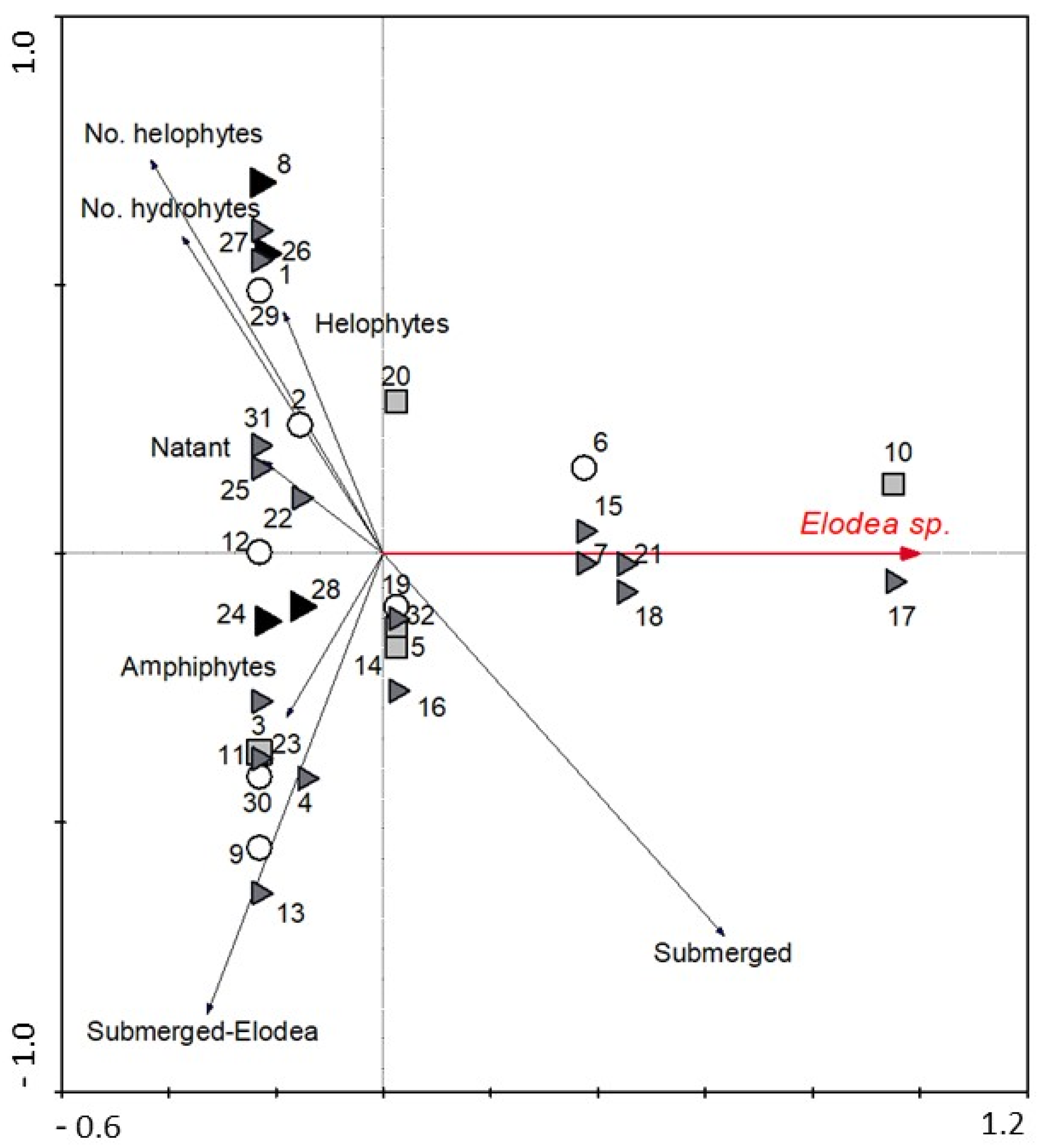
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hynes, H.B.N. The Ecology of Running Waters; University of Toronto Press: Toronto, ON, Canada, 1970. [Google Scholar]
- Zeiringer, B.; Seliger, C.; Greimel, F.; Schmutz, S. River Hydrology, Flow Alteration, and Environmental Flow. In Riverine Ecosystem Management; Springer: Cham, Switzerland, 2018; pp. 67–89. [Google Scholar]
- Tonkin, J.D.; Altermatt, F.; Finn, D.S.; Heino, J.; Olden, J.D.; Pauls, S.U.; Lytle, D.A. The Role of Dispersal in River Network Metacommunities: Patterns, Processes, and Pathways. Freshw. Biol. 2018, 63, 141–163. [Google Scholar] [CrossRef] [Green Version]
- Padial, A.A.; Ceschin, F.; Declerck, S.A.J.; de Meester, L.; Bonecker, C.C.; Lansac-Tôha, F.A.; Rodrigues, L.; Rodrigues, L.C.; Train, S.; Velho, L.F.M.; et al. Dispersal Ability Determines the Role of Environmental, Spatial and Temporal Drivers of Metacommunity Structure. PLoS ONE 2014, 9, e111227. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Molnár, Z. Types and Characteristics of the Oxbow-Lakes in Lower-Tisza-Valley—Classification from Landscape Planning Perspective. Landscape Environ. 2013, 7, 19–25. [Google Scholar]
- Williams, P.; Whitfield, M.; Biggs, J.; Bray, S.; Fox, G.; Nicolet, P.; Sear, D. Comparative Biodiversity of Rivers, Streams, Ditches and Ponds in an Agricultural Landscape in Southern England. Biol. Conserv. 2004, 115, 329–341. [Google Scholar] [CrossRef]
- Shutoh, K.; Yamanouchi, T.; Kato, S.; Yamagishi, H.; Ueno, Y.; Hiramatsu, S.; Nishihiro, J.; Shiga, T. The Aquatic Macrophyte Flora of a Small Pond Revealing High Species Richness in the Aomori Prefecture, Japan. J. Asia Pac. Biodivers. 2019, 12, 448–458. [Google Scholar] [CrossRef]
- Kubiak, A.P.; Krawczyk, R. Diversity of Macrophytes in Riverine Aquatic Habitats: Comparing Active River Channel and Its Cut-Offs. Ann. UMCS Biol. 2014, 69, 49–57. [Google Scholar] [CrossRef] [Green Version]
- Waldon-Rudzionek, B. Is the Flora of Oxbow Lakes Different from That of Fishponds? A Comparison of Two Types of Water Reservoirs in the Noteć River Valley and Bydgoszcz Canal Valley (NW Poland). Ecol. Quest. 2017, 25, 27. [Google Scholar] [CrossRef] [Green Version]
- Panzeca, P.; Troia, A.; Madonia, P. Aquatic Macrophytes Occurrence in Mediterranean Farm Ponds: Preliminary Investigations in North-Western Sicily (Italy). Plants 2021, 10, 1292. [Google Scholar] [CrossRef] [PubMed]
- Obolewski, K. Biodiversity of Macroinvertebrates in Oxbow-Lakes of Early Glacial River Basins in Northern Poland. In Ecosystems Biodiversity; InTech: Rijeka, Croatia, 2011. [Google Scholar]
- Schneiders, A.; Verheyen, R. A Concept of Integrated Water Management Illustrated for Flanders (Belgium). Ecosyst. Health 1998, 4, 256–263. [Google Scholar] [CrossRef]
- Biggs, J.; von Fumetti, S.; Kelly-Quinn, M. The Importance of Small Waterbodies for Biodiversity and Ecosystem Services: Implications for Policy Makers. Hydrobiologia 2017, 793, 3–39. [Google Scholar] [CrossRef]
- Oťaheľová, H.; Valachovič, M.; Hrivnák, R. The Impact of Environmental Factors on the Distribution Pattern of Aquatic Plants along the Danube River Corridor (Slovakia). Limnologica 2007, 37, 290–302. [Google Scholar] [CrossRef]
- Schmidt-Mumm, U.; Janauer, G.A. Macrophyte Assemblages in the Aquatic-Terrestrial Transitional Zone of Oxbow Lakes in the Danube Floodplain (Austria). Folia Geobot. 2016, 51, 251–266. [Google Scholar] [CrossRef]
- Vukov, D.; Ilić, M.; Ćuk, M.; Igić, R.; Janauer, G. The Relationship between Habitat Factors and Aquatic Macrophyte Assemblages in the Danube River in Serbia. Arch. Biol. Sci. 2017, 69, 427–437. [Google Scholar] [CrossRef] [Green Version]
- Gyosheva, B.; Kalchev, R.; Beshkova, M.; Valchev, V. Relationships between Macrophyte Species, Their Life Forms and Environmental Factors in Floodplain Water Bodies from the Bulgarian Danube River Basin. Ecohydrol. Hydrobiol. 2020, 20, 123–133. [Google Scholar] [CrossRef]
- Akasaka, M.; Higuchi, S.; Takamura, N. Landscape- and Local-Scale Actions Are Essential to Conserve Regional Macrophyte Biodiversity. Front. Plant Sci. 2018, 9, 599. [Google Scholar] [CrossRef] [PubMed]
- Bornette, G.; Puijalon, S. Macrophytes: Ecology of Aquatic Plants. In Encyclopedia of Life Sciences (ELS); Wiley: Chichester, UK, 2009. [Google Scholar] [CrossRef]
- Preiner, S.; Dai, Y.; Pucher, M.; Reitsema, R.E.; Schoelynck, J.; Meire, P.; Hein, T. Effects of Macrophytes on Ecosystem Metabolism and Net Nutrient Uptake in a Groundwater Fed Lowland River. Sci. Total Environ. 2020, 721, 137620. [Google Scholar] [CrossRef] [PubMed]
- Glińska-Lewczuk, K.; Burandt, P. Effect of River Straightening on the Hydrochemical Properties of Floodplain Lakes: Observations from the Łyna and Drwęca Rivers, N Poland. Ecol. Eng. 2011, 37, 786–795. [Google Scholar] [CrossRef]
- Ambrožič, Š.; Gaberščik, A.; Vrezec, A.; Germ, M. Hydrophyte Community Structure Affects the Presence and Abundance of the Water Beetle Family Dytiscidae in Water Bodies along the Drava River. Ecol. Eng. 2018, 120, 397–404. [Google Scholar] [CrossRef]
- Brysiewicz, A.; Czerniejewski, P.; Bonisławska, M. Effect of Diverse Abiotic Conditions on the Structure and Biodiversity of Ichthyofauna in Small, Natural Water Bodies Located on Agricultural Lands. Water 2020, 12, 2674. [Google Scholar] [CrossRef]
- Baattrup-Pedersen, A.; Riis, T. Macrophyte Diversity and Composition in Relation to Substratum Characteristics in Regulated and Unregulated Danish Streams. Freshw. Biol. 1999, 42, 375–385. [Google Scholar] [CrossRef]
- Hansen, J.P.; Wikström, S.A.; Axemar, H.; Kautsky, L. Distribution Differences and Active Habitat Choices of Invertebrates between Macrophytes of Different Morphological Complexity. Aquat. Ecol. 2011, 45, 11–22. [Google Scholar] [CrossRef]
- Haslam, S.M. River Plants of Western Europe: The Macrophytic Vegetation of Watercourses of the European Economic Community; Cambridge University Press: Cambridge, UK, 1987; ISBN 0-521-26427-8. [Google Scholar]
- Schneider, S.; Melzer, A. The Trophic Index of Macrophytes (TIM)—A New Tool for Indicating the Trophic State of Running Waters. Int. Rev. Hydrobiol. 2003, 88, 49–67. [Google Scholar] [CrossRef]
- Szpakowska, B.; Świerk, D.; Pajchrowska, M.; Gołdyn, R. Verifying the Usefulness of Macrophytes as an Indicator of the Status of Small Waterbodies. Sci. Total Environ. 2021, 798, 149279. [Google Scholar] [CrossRef]
- Kuhar, U.; Germ, M.; Gaberščik, A.; Urbanič, G. Development of a River Macrophyte Index (RMI) for Assessing River Ecological Status. Limnologica 2011, 41, 235–243. [Google Scholar] [CrossRef]
- van Geest, G.J.; Coops, H.; Roijackers, R.M.M.; Buijse, A.D.; Scheffer, M. Succession of Aquatic Vegetation Driven by Reduced Water-Level Fluctuations in Floodplain Lakes. J. Appl. Ecol. 2005, 42, 251–260. [Google Scholar] [CrossRef]
- Lu, J.; Bunn, S.E.; Burford, M.A. Nutrient Release and Uptake by Littoral Macrophytes during Water Level Fluctuations. Sci. Total Environ. 2018, 622–623, 29–40. [Google Scholar] [CrossRef] [PubMed]
- Bogut, I.; Vidaković, J.; Palijan, G.; Čerba, D. Benthic Macroinvertebrates Associated with Four Species of Macrophytes. Biologia 2007, 62, 600–606. [Google Scholar] [CrossRef]
- Pyšek, P.; Richardson, D.M. Invasive Species, Environmental Change and Management, and Health. Annu. Rev. Environ. Resour. 2010, 35, 25–55. [Google Scholar] [CrossRef] [Green Version]
- Janauer, G.; Exler, N.; Anačkov, G.; Barta, V.; Berczik, Á.; Boža, P.; Dinka, M.; Georgiev, V.; Germ, M.; Holcar, M.; et al. Distribution of the Macrophyte Communities in the Danube Reflects River Serial Discontinuity. Water 2021, 13, 918. [Google Scholar] [CrossRef]
- Ricciardi, A.; Hoopes, M.F.; Marchetti, M.P.; Lockwood, J.L. Progress toward Understanding the Ecological Impacts of Nonnative Species. Ecol. Monogr. 2013, 83, 263–282. [Google Scholar] [CrossRef]
- Katsanevakis, S.; Wallentinus, I.; Zenetos, A.; Leppäkoski, E.; Çinar, M.E.; Oztürk, B.; Grabowski, M.; Golani, D.; Cardoso, A.C. Impacts of Invasive Alien Marine Species on Ecosystem Services and Biodiversity: A Pan-European Review. Aquat. Invasions 2014, 9, 391–423. [Google Scholar] [CrossRef]
- Vimercati, G.; Probert, A.F.; Volery, L.; Bernardo-Madrid, R.; Bertolino, S.; Céspedes, V.; Essl, F.; Evans, T.; Gallardo, B.; Gallien, L.; et al. The EICAT+ Framework Enables Classification of Positive Impacts of Alien Taxa on Native Biodiversity. PLoS Biol. 2022, 20, e3001729. [Google Scholar] [CrossRef] [PubMed]
- Kohler, A.; Janauer, G.A. Zur Methodik Der Untersuchung von Aquatischen Makrophyten in Flie_gewassern. In Handbuch Angewandte Limnologie; Steinberg, C.H., Bernhardt, H., Klapper, H., Eds.; Ecomed: Landsberg am Lech, Germany, 1995; pp. 3–22. [Google Scholar]
- Euro+Med PlantBase—The Information Resource for Euro-Mediterranean Plant Diversity. Available online: http://ww2.bgbm.org/EuroPlusMed/ (accessed on 1 September 2022).
- Schaumburg, J.; Schranz, C.; Foerster, J.; Gutowski, A.; Hofmann, G.; Meilinger, P.; Schneider, S.; Schmedtje, U. Ecological Classification of Macrophytes and Phytobenthos for Rivers in Germany According to the Water Framework Directive. Limnologica 2004, 34, 283–301. [Google Scholar] [CrossRef] [Green Version]
- Braun-Blanquet, J. Pflanzensoziologie, Grundzüge Der Vegetationskunde, 3rd ed.; Springer: Berlin, Germany, 1964. [Google Scholar]
- Germ, M.; Janež, V.; Gaberščik, A.; Zelnik, I. Diversity of Macrophytes and Environmental Assessment of the Ljubljanica River (Slovenia). Diversity 2021, 13, 278. [Google Scholar] [CrossRef]
- Hammer, Ø.; Harper, D.A.T.; Ryan, P.D. PAST: Paleontological Statistics Software Package for Education and Data Analysis. Palaeontol. Electron. 2001, 4, 1–9. [Google Scholar]
- ter Braak, C.J.F.; Verdonschot, P.F.M. Canonical Correspondence Analysis and Related Multivariate Methods in Aquatic Ecology. Aquat. Sci. 1995, 57, 255–289. [Google Scholar] [CrossRef]
- ter Braak, C.J.F.; Smilauer, P. CANOCO Reference Manual and CanoDraw for Windows User’s Guide: Software for Canonical Community Ordination (Version 4.5); Canoco: Ithaca, NY, USA, 2002. [Google Scholar]
- Huang, Y.; Chen, X.-S.; Li, F.; Hou, Z.-Y.; Li, X.; Zeng, J.; Deng, Z.-M.; Zou, Y.-A.; Xie, Y.-H. Community Trait Responses of Three Dominant Macrophytes to Variations in Flooding During 2011–2019 in a Yangtze River-Connected Floodplain Wetland (Dongting Lake, China). Front. Plant Sci. 2021, 12, 604677. [Google Scholar] [CrossRef] [PubMed]
- Zelnik, I.; Kuhar, U.; Holcar, M.; Germ, M.; Gaberščik, A. Distribution of Vascular Plant Communities in Slovenian Watercourses. Water 2021, 13, 1071. [Google Scholar] [CrossRef]
- Pereira, K.M.; Hefler, S.M.; Trentin, G.; Rolon, A.S. Influences of Landscape and Climatic Factors on Aquatic Macrophyte Richness and Composition in Ponds. Flora 2021, 279, 151811. [Google Scholar] [CrossRef]
- Zehnsdorf, A.; Hussner, A.; Eismann, F.; Rönicke, H.; Melzer, A. Management Options of Invasive Elodea Nuttallii and Elodea Canadensis. Limnologica 2015, 51, 110–117. [Google Scholar] [CrossRef]
- Barrat-Segretain, M.-H. Invasive Species in the Rhône River Floodplain (France): Replacement of Elodea Canadensis Michaux by E. Nuttallii St. John in Two Former River Channels. Arch. Hydrobiol. 2001, 152, 237–251. [Google Scholar] [CrossRef]
- Barrat-Segretain, M.; Elger, A. Experiments on Growth Interactions between Two Invasive Macrophyte Species. J. Veg. Sci. 2004, 15, 109–114. [Google Scholar] [CrossRef]
- Szabó, S.; Peeters, E.T.H.M.; Borics, G.; Veres, S.; Nagy, P.T.; Lukács, B.A. The Ecophysiological Response of Two Invasive Submerged Plants to Light and Nitrogen. Front. Plant Sci. 2020, 10, 01747. [Google Scholar] [CrossRef] [Green Version]
- Szabó, S.; Peeters, E.T.H.M.; Várbíró, G.; Borics, G.; Lukács, B.A. Phenotypic Plasticity as a Clue for Invasion Success of the Submerged Aquatic Plant Elodea Nuttallii. Plant Biol. 2019, 21, 54–63. [Google Scholar] [CrossRef] [Green Version]
- Mazej Grudnik, Z.; Germ, M. Spatial Pattern of Native Species Myriophyllum Spicatum and Invasive Alien Species Elodea Nuttallii after Introduction of the Latter One into the Drava River (Slovenia). Biologia 2013, 68, 202–209. [Google Scholar] [CrossRef] [Green Version]
- Barrat-Segretain, M.-H.; Elger, A.; Sagnes, P.; Puijalon, S. Comparison of Three Life-History Traits of Invasive Elodea Canadensis Michx. and Elodea Nuttallii (Planch.) H. St. John. Aquat. Bot. 2002, 74, 299–313. [Google Scholar] [CrossRef]
- Ellenberg, H.; Weber, H.; Düll, R.; Wirth, V.; Werner, W.; Pauliβen, D. Zeigerwertevon Pflanzen in Mitteleuropa; Verlag Erich Goltze: Göttingen, Germany, 1992. [Google Scholar]
- Grudnik, Z.M.; Jelenko, I.; Germ, M. Influence of Abiotic Factors on Invasive Behaviour of Alien Species Elodea Nuttallii in the Drava River (Slovenia). Ann. Limnol. Int. J. Limnol. 2014, 50, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Szabo, S.; Scheffer, M.; Roijackers, R.; Waluto, B.; Braun, M.; Nagy, P.T.; Borics, G.; Zambrano, L. Strong Growth Limitation of a Floating Plant (Lemna Gibba) by the Submerged Macrophyte (Elodea Nuttallii) under Laboratory Conditions. Freshw. Biol. 2010, 55, 681–690. [Google Scholar] [CrossRef]
- van Donk, E.; Gulati, R.D.; Iedema, A.; Meulemans, J.T. Macrophyte-Related Shifts in the Nitrogen and Phosphorus Contents of the Different Trophic Levels in a Biomanipulated Shallow Lake. In Nutrient Dynamics and Retention in Land/Water Ecotones of Lowland, Temperate Lakes and Rivers; Springer: Dordrecht, The Netherland, 1993; pp. 19–26. [Google Scholar] [CrossRef] [Green Version]
- Netten, J.J.C.A.; Nes, E.H.; Scheffer, M.; Roijackers, R.M.M. Effect of Temperature and Nutrients on the Competition between Free-Floating Salvinia Natans and Submerged Elodea Nuttallii in Mesocosms. Fundament. Appl. Limnol. 2010, 177, 125–132. [Google Scholar] [CrossRef]
- Klančnik, K.; Iskra, I.; Gradinjan, D.; Gaberščik, A. The Quality and Quantity of Light in the Water Column Are Altered by the Optical Properties of Natant Plant Species. Hydrobiologia 2018, 812, 203–212. [Google Scholar] [CrossRef]
- Kuhar, U.; Germ, M.; Gaberščik, A. Habitat Characteristics of an Alien Species Elodea Canadensis in Slovenian Watercourses. Hydrobiologia 2010, 656, 205–212. [Google Scholar] [CrossRef]
- Hoff, H.K.; Thum, R.A. Hybridization and Invasiveness in Eurasian Watermilfoil (Myriophyllum Spicatum): Is Prioritizing Hybrids in Management Justified? Invasive Plant Sci. Manag. 2022, 15, 3–8. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Location | Width of Riparian Zone | Land-Use behind the Riparian Zone | Type |
|---|---|---|---|
| 1 | 1–5 m | arable land, grassland, houses | river |
| 2 | 1–5 m | arable land, grassland, houses | pond |
| 3 | 1–5 m | arable land, grassland, houses | channel |
| 4 | 1–5 m | arable land, grassland, houses | channel |
| 5 | 1–5 m | arable land, grassland, houses | oxbow |
| 6 | <1 m | mainly arable land or urban area | pond |
| 7 | 1–5 m | mainly arable land or urban area | channel |
| 8 | <1 m | arable land, grassland, houses | river |
| 9 | <1 m | mainly arable land or urban area | pond |
| 10 | 1–5 m | mainly arable land or urban area | oxbow |
| 11 | <1 m | mainly arable land or urban area | pond |
| 12 | 5–30 m | grassland, forest and/or wetland, some arable land | pond |
| 13 | 5–30 m | grassland, forest and/or wetland, some arable land | channel |
| 14 | 1–5 m | mainly arable land or urban area | channel |
| 15 | <1 m | arable land, grassland, houses | channel |
| 16 | 1–5 m | arable land, grassland, houses | channel |
| 17 | <1 m | arable land, grassland, houses | oxbow |
| 18 | <1 m | mainly arable land or urban area | channel |
| 19 | 5–30 m | arable land, grassland, houses | pond |
| 20 | <1 m | mainly arable land or urban area | oxbow |
| 21 | 1–5 m | arable land, grassland, houses | channel |
| 22 | 1–5 m | arable land, grassland, houses | channel |
| 23 | <1 m | arable land, grassland, houses | oxbow |
| 24 | 1–5 m | arable land, grassland, houses | river |
| 25 | <1 m | arable land, grassland, houses | channel |
| 26 | <1 m | mainly arable land or urban area | channel |
| 27 | <1 m | mainly arable land or urban area | channel |
| 28 | 5–30 m | arable land, grassland, houses | river |
| 29 | 1–5 m | arable land, grassland, houses | pond |
| 30 | 1–5 m | arable land, grassland, houses | pond |
| 31 | 1–5 m | grassland, forest and/or wetland, some arable land | channel |
| 32 | 1–5 m | mainly arable land or urban area | oxbow |
| Variable | E. canadensis | E. nuttallii | E. canadensis and nuttallii |
|---|---|---|---|
| Number of native taxa | n.s. | n.s. | n.s. |
| Total abundance of plants | n.s. | 0.2679 | 0.2669 |
| Shannon–Wiener diversity index | n.s. | n.s. | n.s. |
| Concentration of O2 [mg/L] | n.s. | 0.2679 | n.s. |
| Temperature of the water [°C] | n.s. | 0.2614 * | n.s. |
| Cover of floating-leaved macrophytes | n.s. | −0.2782 | n.s. |
| Land-use in the catchment | n.s. | 0.2988 | 0.2805 |
| Abundance of Nymphaea alba | n.s. | −0.2617 | −0.3079 |
| Abundance of Spirodela polyrhiza | n.s. | −0.2886 | n.s. |
| Oxbows | Ponds | Side-Channels | River | |
|---|---|---|---|---|
| E. canadensis | 0 | 0 | 15.7 | 0.75 |
| E. nuttallii | 14.5 | 48 | 12.2 | 4.5 |
| Nr. of native taxa | 8 | 10,5 | 11.4 | 9.5 |
| S–W diversity index | 1.7 | 1.9 | 2 | 1.9 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zelnik, I.; Germ, M.; Kuhar, U.; Gaberščik, A. Waterbodies in the Floodplain of the Drava River Host Species-Rich Macrophyte Communities despite Elodea Invasions. Diversity 2022, 14, 870. https://doi.org/10.3390/d14100870
Zelnik I, Germ M, Kuhar U, Gaberščik A. Waterbodies in the Floodplain of the Drava River Host Species-Rich Macrophyte Communities despite Elodea Invasions. Diversity. 2022; 14(10):870. https://doi.org/10.3390/d14100870
Chicago/Turabian StyleZelnik, Igor, Mateja Germ, Urška Kuhar, and Alenka Gaberščik. 2022. "Waterbodies in the Floodplain of the Drava River Host Species-Rich Macrophyte Communities despite Elodea Invasions" Diversity 14, no. 10: 870. https://doi.org/10.3390/d14100870