

Risk Screening and Distribution of the Invasive Amphipod Dikerogammarus villosus (Sowinsky, 1894) in the River Adda (Northern Italy)

Abstract
:1. Introduction
2. Materials and Methods
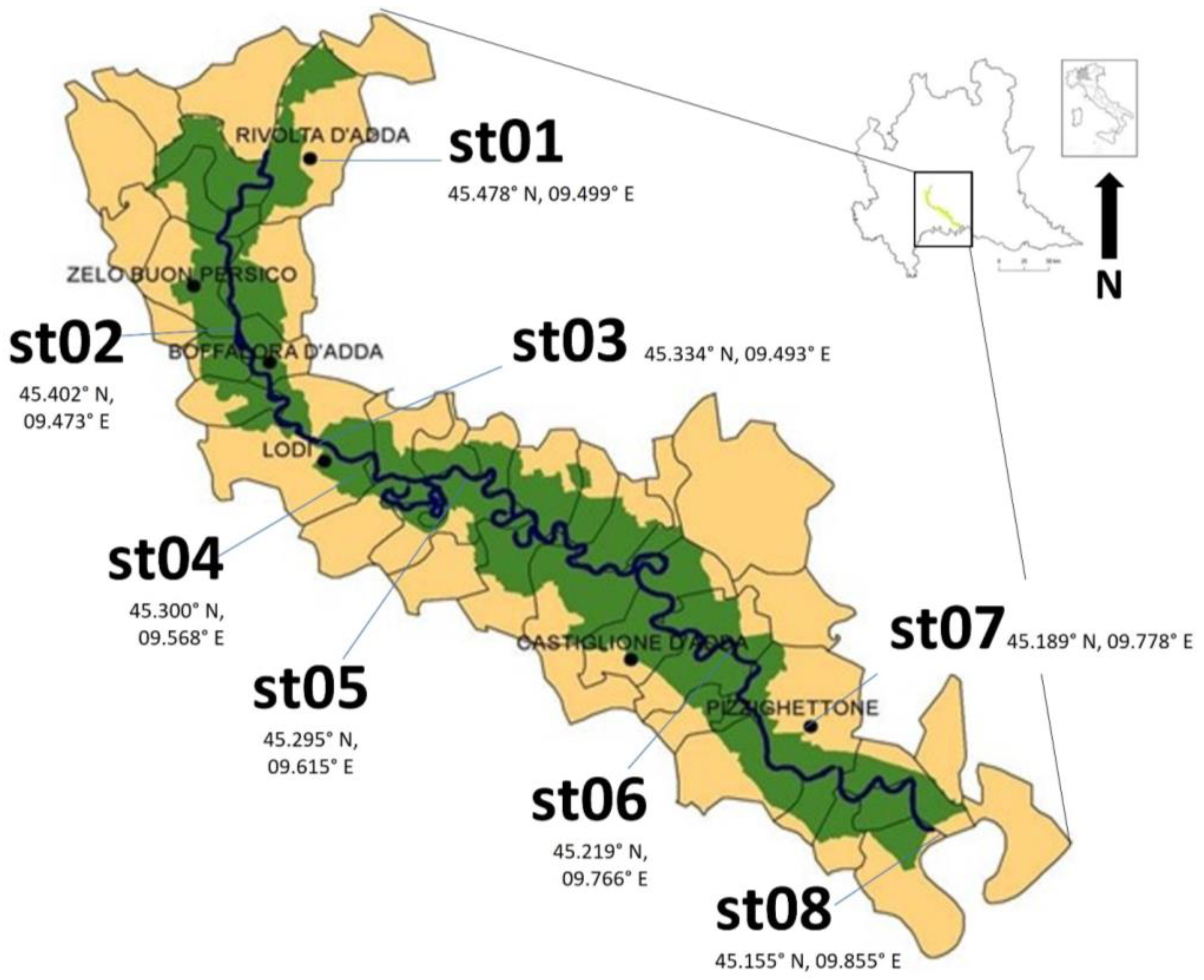
2.1. Risk Assessment Area and Sampling Method
2.2. Risk Screening Tools
2.2.1. Aquatic Species Invasiveness Screening Kit (AS-ISK)
2.2.2. Harmonia+
2.3. Impact Assessment Tool
The Generic Impact Scoring System (GISS)
3. Results
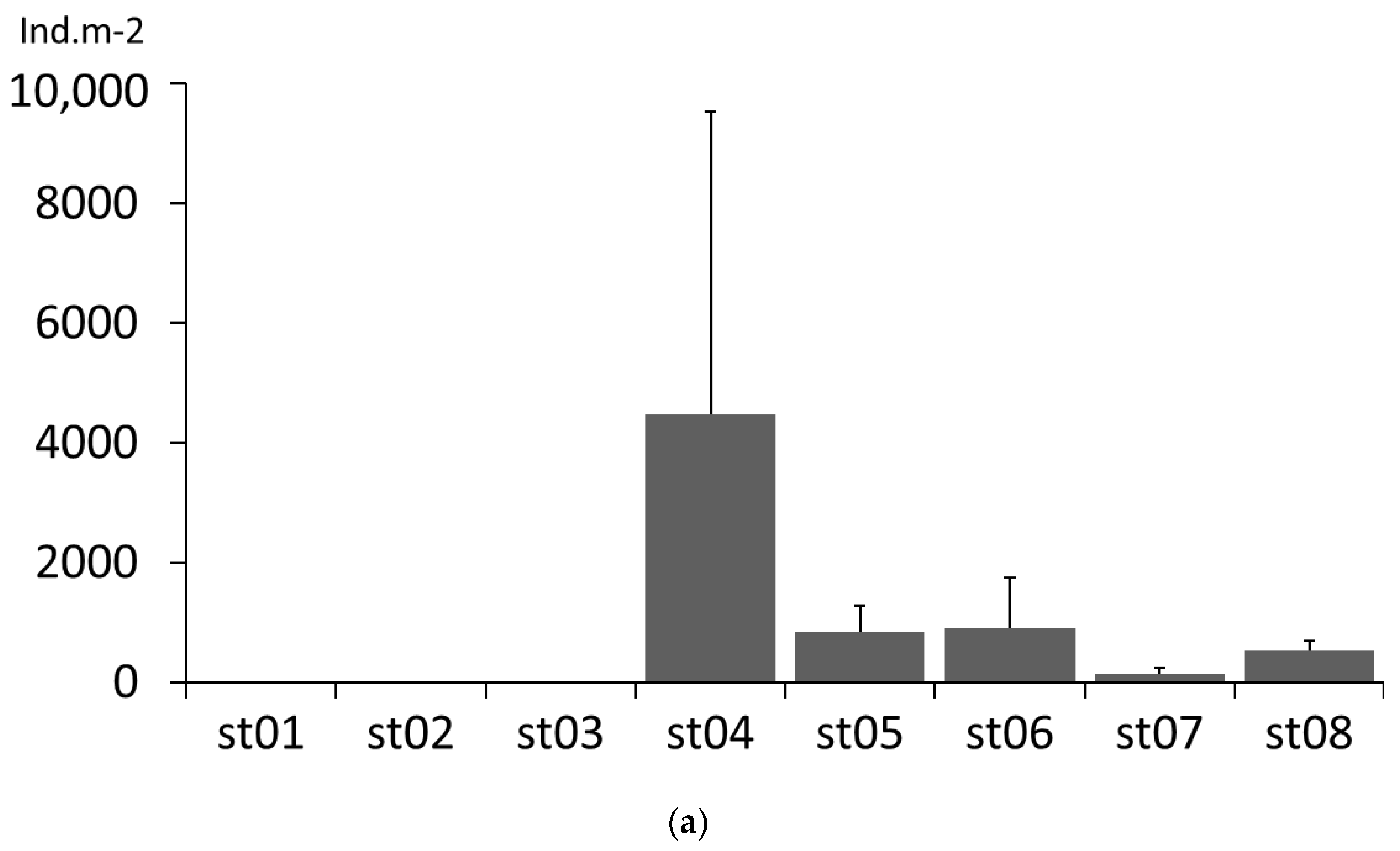
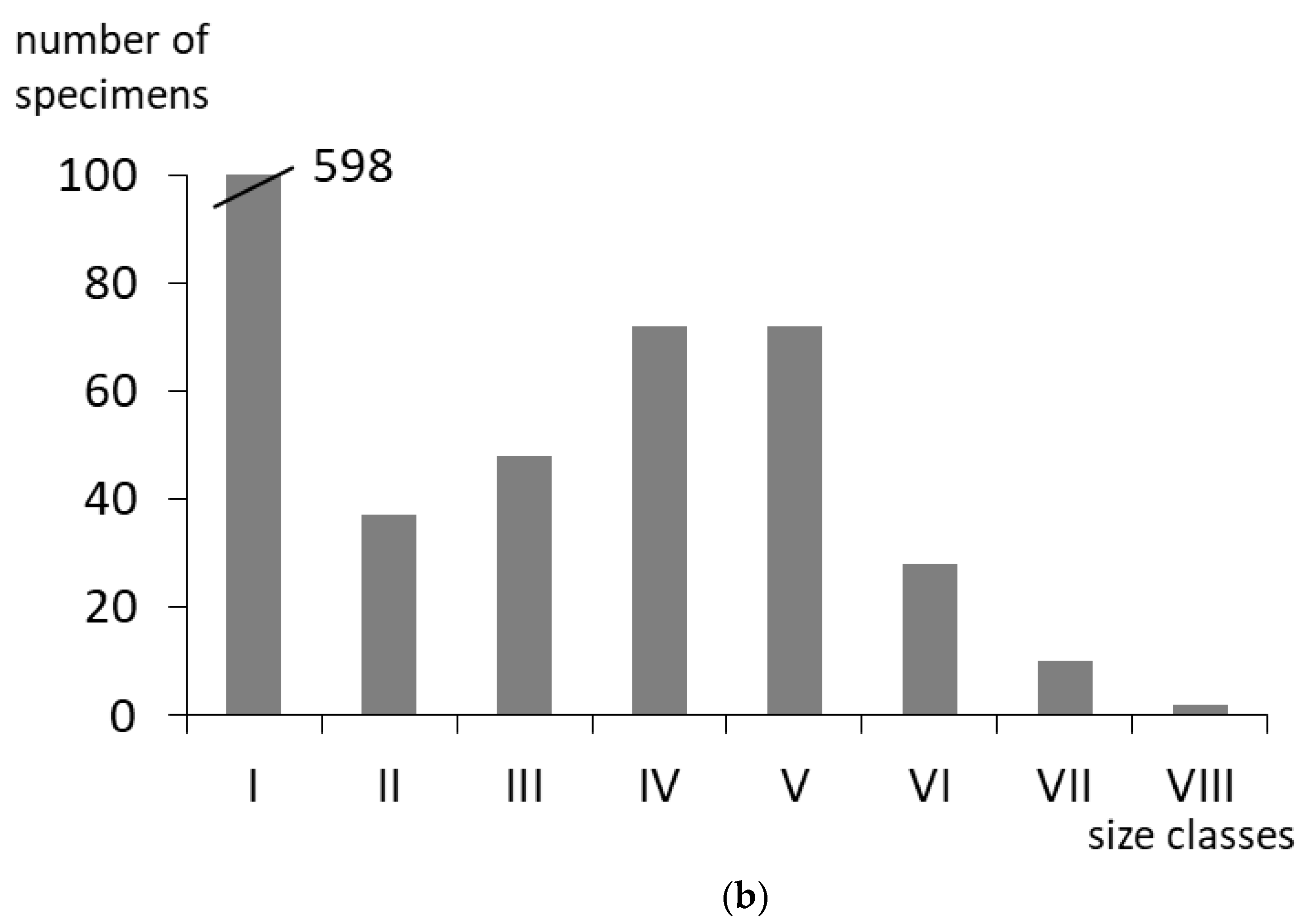
3.1. Density, Distribution and Size Classes of Dikerogammarus villosus
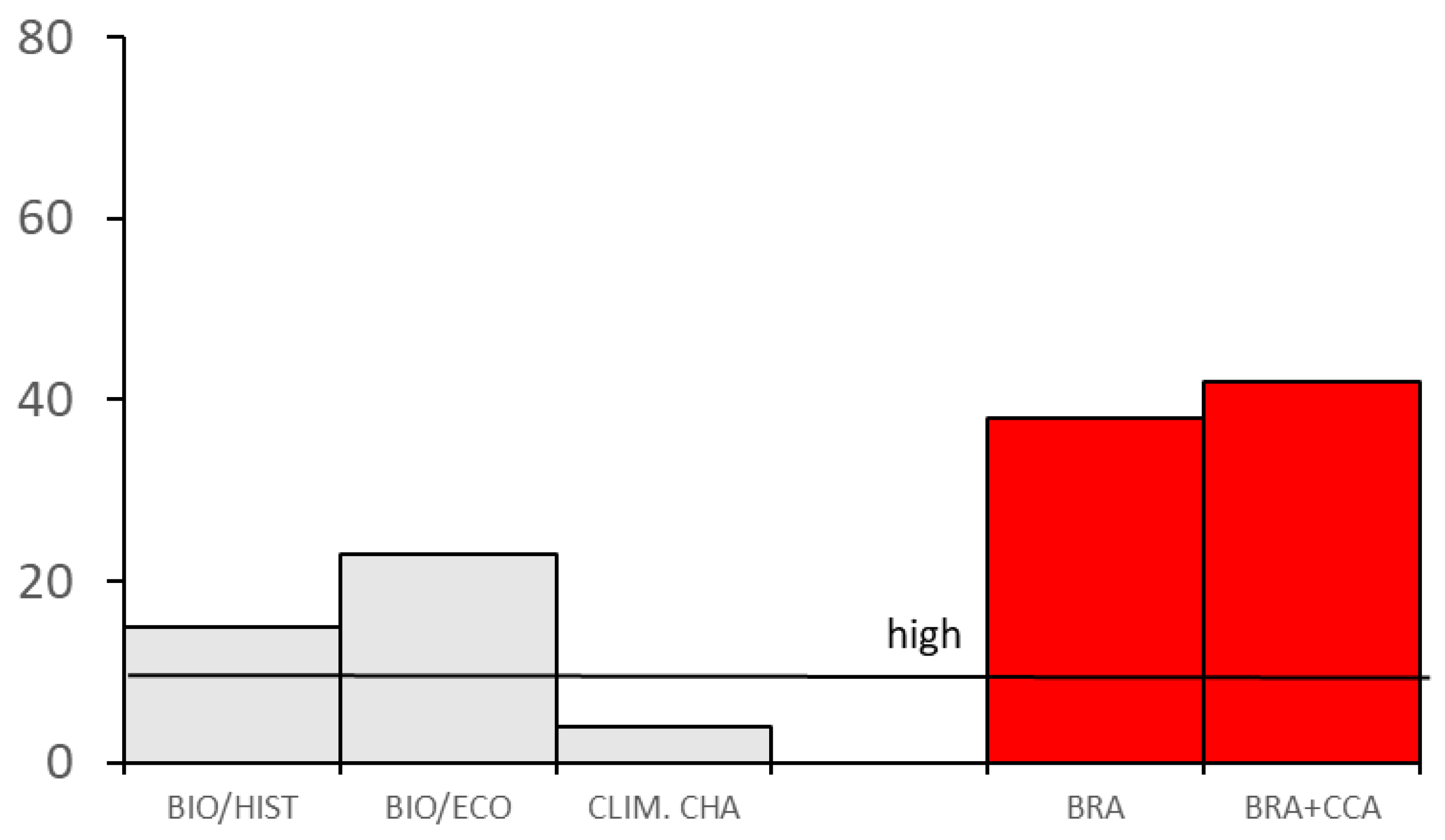
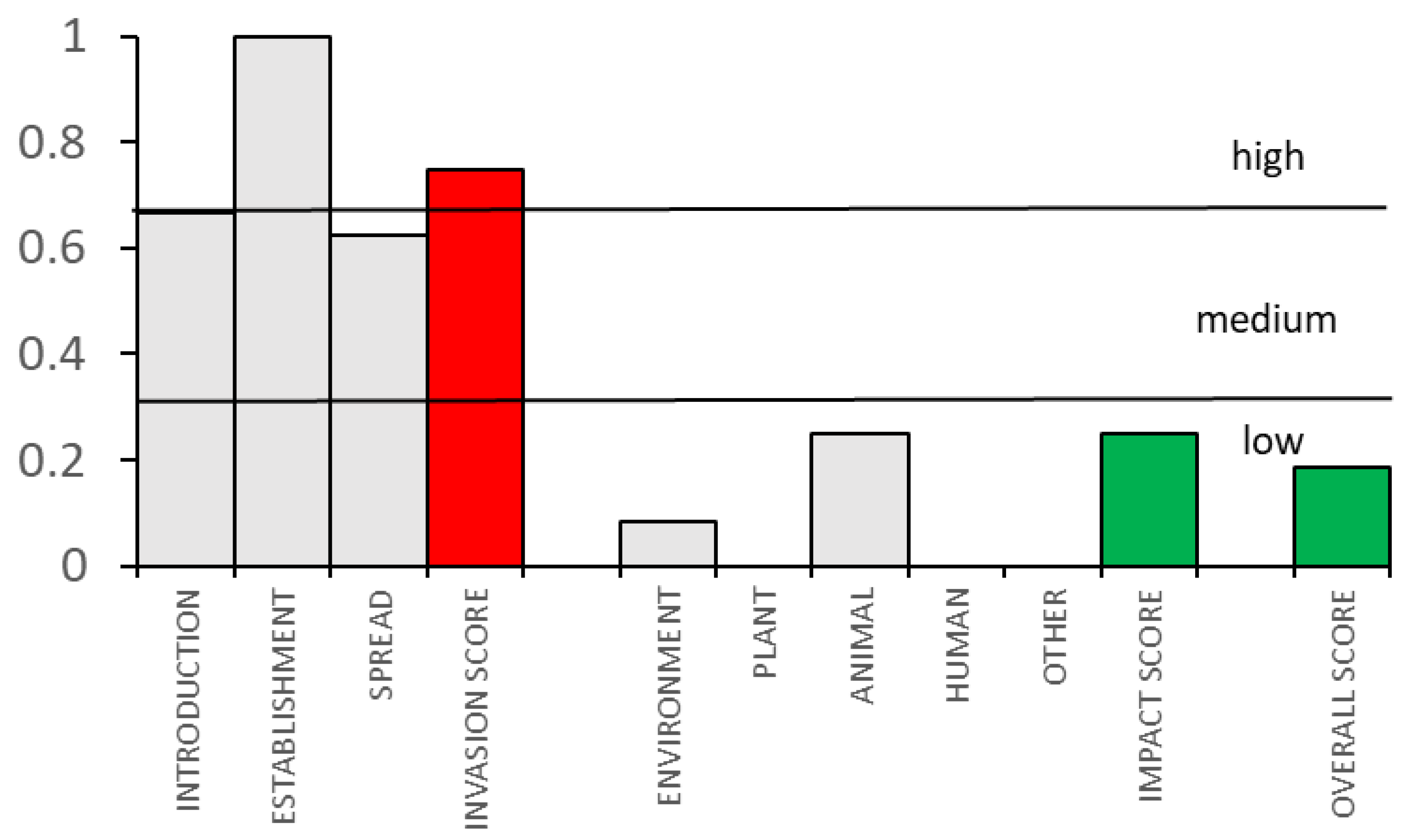
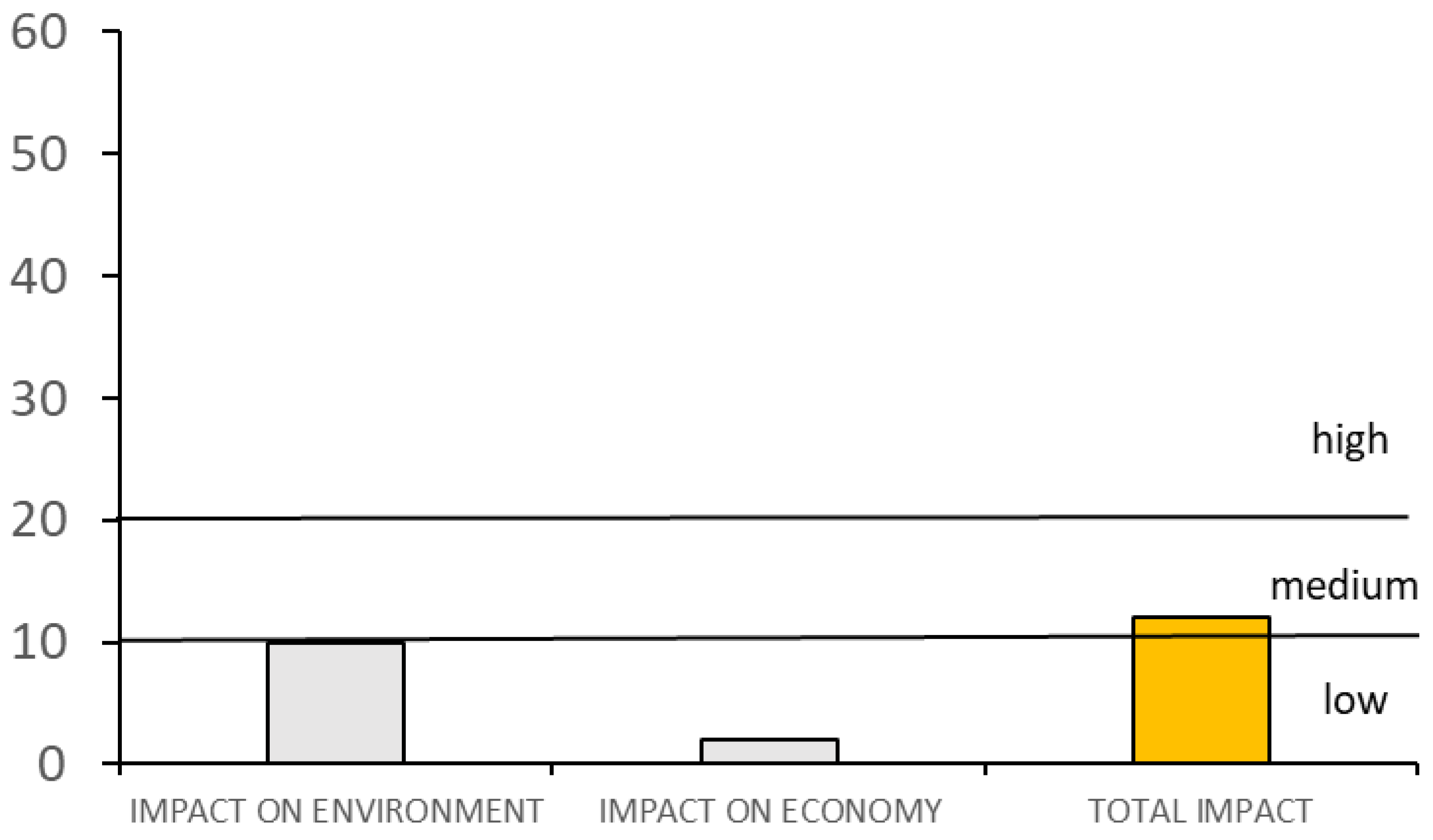
3.2. Definition of the Level of Invasiveness and Impacts of Dikerogammarus villosus
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- IUCN; IUCN Species Survival Commission (SSC); IUCN Species Survival Commission (SSC), Invasive Species Specialist Group. IUCN EICAT Categories and Criteria: The Environmental Impact Classification for Alien Taxa (EICAT), 1st ed.; IUCN: Gland, Switzerland; Cambridge, UK, 2020. [Google Scholar] [CrossRef]
- Leung, B.; Roura Pascual, N.; Bacher, S.; Heikkiläla, J.; Brotons, L.; Burgman, M.A.; Dehnen Schmutz, K.; Essl, F.; Hulme, P.E.; Richardson, D.M.; et al. TEASIng apart alien species risk assessments: A framework for best practices. Ecol. Lett. 2012, 15, 1475–1493. [Google Scholar] [CrossRef] [PubMed]
- Guareschi, S.; Wood, P.J. Biological Invasions of River Ecosystems: A Flow of Implications, Challenges, and Research Opportunities. In Imperiled: The Encyclopedia of Conservation; Dominick, A., Della, S., Goldstein, M.I., Eds.; Elsevier: Amsterdam, The Netherlands, 2022; pp. 485–498. [Google Scholar] [CrossRef]
- Occhipinti-Ambrogi, A.; Galil, B.S. A uniform terminology on bioinvasions: A chimera or an operative tool? Mar. Pollut. Bull. 2004, 49, 688–694. [Google Scholar] [CrossRef] [PubMed]
- Charles, H.; Dukes, J.S. Impacts of invasive species on ecosystem services. In Biological Invasions; Nentwig, W., Ed.; Springer: Berlin/Heidelberg, Germany, 2007; pp. 217–237. [Google Scholar]
- Langor, D.W.; Sweeney, J. Ecological impacts of non-native invertebrates and fungi on terrestrial ecosystems. Biol. Invasions 2009, 11, 1–3. [Google Scholar] [CrossRef] [PubMed]
- Kenis, M.; Auger-Rozenberg, M.A.; Roques, A.; Timms, L.; Péré, C.; Cock, M.J.W. Ecological effects of invasive alien insects. Biol. Invasions 2009, 11, 21–45. [Google Scholar] [CrossRef]
- Kettunen, M.; Genovesi, P.; Gollasch, S.; Pagad, S.; Starfinger, U.; ten Brink, P.; Shine, C. Technical Support to EU Strategy on Invasive Species (IAS)—Assessment of the Impacts of IAS in Europe and the EU, 2009, Final Report. Available online: https://ec.europa.eu/environment/nature/invasivealien/docs/Kettunen2009IASTask%201.pdf (accessed on 29 September 2022).
- Vilà, M.; Basnou, C.; Pysek, P.; Josefsson, M.; Genovesi, P.; Gollasch, S.; Nentwig, W. How well do we understand the impacts of alien species on ecosystem services? A pan-European cross-taxa assessment. Front. Ecol. Environ. 2009, 8, 135–144. [Google Scholar] [CrossRef] [Green Version]
- Haubrock, P.; Turbelin, A.; Cuthbert, R.; Novoa, A.; Taylor, N.; Angulo, E.; Ballesteros-Mejia, L.; Bodey, T.; Capinha, C.; Essl, F.; et al. Economic costs of invasive alien species across Europe. NeoBiota 2021, 67, 153–190. [Google Scholar] [CrossRef]
- Cox, J.G.; Lima, S.L. Naiveté and an aquatic–terrestrial dichotomy in the effects of introduced predators. Trends Ecol. Evol. 2006, 21, 674–680. [Google Scholar] [CrossRef] [PubMed]
- Gallardo, B.; Clavero, M.; Sánchez, M.I.; Vilà, M. Global ecological impacts of invasive species in aquatic ecosystems. Glob. Chang. Biol. 2016, 22, 151–163. [Google Scholar] [CrossRef]
- Rinaldo, A.; Gatto, M.; Rodríguez-Iturbe, I. River Networks as Ecological Corridors: Species, Populations, Pathogens; Cambridge University Press: Cambridge, UK, 2020; p. 438. [Google Scholar]
- Gozlan, R.E.; Britton, J.R.; Cowx, I.; Copp, G.H. Current knowledge on non-native freshwater fish introductions. J. Fish Biol. 2010, 76, 751–786. [Google Scholar] [CrossRef]
- Paganelli, D.; Pandolfi, A.; Sconfietti, R.; Marchini, A.; Vilizzi, L. Potential invasiveness by non-indigenous macrozoobenthos in the secondary hydrographic system of a temperate-climate river catchment. Ecol. Indic. 2018, 88, 274–281. [Google Scholar] [CrossRef]
- Strayer, D.L. Alien species in fresh waters: Ecological effects, interactions with other stressors, and prospects for the future. Freshw. Biol. 2010, 55, 152–174. [Google Scholar] [CrossRef]
- Olden, J.D. Biotic homogenization: A new research agenda for conservation biogeography. J. Biogeogr. 2006, 33, 2027–2039. [Google Scholar] [CrossRef]
- Laverty, C.; Wolfgang, N.; Dick, J.T.A.; Lucy, F.E. Alien aquatics in Europe: Assessing the relative environmental and socioeconomic impacts of invasive aquatic macroinvertebrates and other taxa. Manag. Biol. Invasions 2015, 6, 341–350. [Google Scholar] [CrossRef] [Green Version]
- Diagne, C.; Leroy, B.; Gozlan, R.E.; Vaissière, A.C.; Assailly, C.; Nuninger, L.; Roiz, D.; Jourdain, F.; Jarić, I.; Courchamp, F. InvaCost, a public database of the economic costs of biological invasions worldwide. Sci. Data 2020, 7, 277. [Google Scholar] [CrossRef] [PubMed]
- Boets, P.; Pauwels, I.S.; Lock, K.; Goethals, P.L.M. Using an integrated modelling approach for risk assessment of the ‘killer shrimp’ Dikerogammarus villosus. River Res. Appl. 2014, 30, 403–412. [Google Scholar] [CrossRef]
- Kobak, J.; Jermacz, Ł.; Marcińczyk, J.; Bartoszyńska, E.; Rutkowska, D.; Pawłowska, K. Abiotic factors affecting habitat selection by two invasive gammarids Dikerogammarus villosus and Pontogammarus robustoides. Hydrobiologia 2017, 797, 247–263. [Google Scholar] [CrossRef] [Green Version]
- Minchin, J.D.; Arbačiauskas, K.; Daunys, D.; Ezhova, E.; Grudule, N.; Kotta, J.; Molchanova, N.; Olenin, S.; Višinskienė, G.; Strake, S. Rapid expansion and facilitating factors of the Ponto-Caspian invader Dikerogammarus villosus within the eastern Baltic Sea. Aquat. Invasions 2019, 14, 165–181. [Google Scholar] [CrossRef]
- Rewicz, T.; Wattier, R.; Grabowski, M.; Rigaud, T.; Bącela-Spychalska, K. Out of the Black Sea: Phylogeography of the Invasive Killer Shrimp Dikerogammarus villosus across Europe. PLoS ONE 2015, 10, e0118121. [Google Scholar] [CrossRef]
- Devin, S.; Piscart, C.; Beisel, J.N.; Moreteau, J.C. Ecological traits of the amphipod invader Dikerogammarus villosus on a mesohabitat scale. Arch. Hydrobiol. 2003, 158, 43–56. [Google Scholar] [CrossRef]
- Casellato, S.; La Piana, G.; Latella, L.; Ruffo, S. Dikerogammarus villosus (Sowinsky, 1894) (Crustacea, Amphipoda, Gammaridae) for the first time in Italy. Ital. J. Zool. 2006, 73, 97–104. [Google Scholar] [CrossRef]
- Casellato, S.; Visentin, A.; La Piana, G. The predatory impact of Dikerogammarus villosus on fish. In Biological Invaders in Inland Waters: Profiles, Distribution and Threats; Gherardi, F., Ed.; Springer: Berlin/Heidelberg, Germany, 2007; pp. 495–507. [Google Scholar] [CrossRef]
- Casellato, S.; Masiero, L.; La Piana, G.; Gigliotti, F. The alien amphipod crustacean Dikerogammarus villosus in Lake Garda (N-Italy): The invasion continues. Neobiota 2008, 7, 115–122. [Google Scholar]
- Tricarico, E.; Mazza, G.; Orioli, G.; Rossano, C.; Scapini, F.; Gherardi, F. The killer shrimp, Dikerogammarus villosus (Sowinsky, 1894), is spreading in Italy. Aquat. Invasions 2010, 5, 211–214. [Google Scholar] [CrossRef]
- Jazdzewski, K.; Konopacka, A. Invasive Ponto-Caspian species in waters of the Vistula and Oder basins and the southern Baltic Sea. In Invasive Aquatic Species of Europe: Distribution, Impacts and Management; Leppäkoski, E., Gollasch, S., Dordrecht, O., Eds.; Kluwer Academic Publishers: Amsterdam, The Netherlands, 2002; pp. 384–398. [Google Scholar]
- de Vaate, A.B.; Jazdzewski, K.; Ketelaars, H.A.M.; Gollash, S.; van der Velde, G. Geographical patterns in range extension of Ponto-Caspian macroinvertebrate species in Europe. Can. J. Fish. Aquat. Sci. 2002, 59, 1159–1174. [Google Scholar] [CrossRef] [Green Version]
- Cuthbert, R.N.; Kotronaki, S.G.; Dick, J.T.; Briski, E. Salinity tolerance and geographical origin predict global alien amphipod invasions. Biol. Lett. 2020, 16, 20200354. [Google Scholar] [CrossRef] [PubMed]
- Bacela-Spychalska, K.; Grabowski, M.; Rewicz, T.; Konopacka, A.; Wattier, R. The ‘killer shrimp’ Dikerogammarus villosus (Crustacea, Amphipoda) invading Alpine lakes: Overland transport by recreational boats and scuba-diving gear as potential entry vectors? Aquat. Conserv. Mar. Freshw. Ecosyst. 2013, 23, 606–618. [Google Scholar] [CrossRef]
- Dick, J.; Platvoet, D.; David, K. Predatory impact of the freshwater invader Dikerogammarus villosus (Crustacea: Amphipoda). Can. J. Fish. Aquat. Sci. 2002, 59, 1078–1084. [Google Scholar] [CrossRef] [Green Version]
- Platvoet, D.; Dick, J.; Konijnendijk, N. Feeding on micro-algae in the invasive Ponto-Caspian amphipod Dikerogammarus villosus (Sowinsky, 1894). Aquat. Ecol 2006, 40, 237–245. [Google Scholar] [CrossRef]
- Taylor, N.G.; Dunn, A.M. Size matters: Predation of fish eggs and larvae by native and invasive amphipods. Biol. Invasions 2017, 19, 89–107. [Google Scholar] [CrossRef]
- Warren, D.A.; Bradbeer, S.J.; Dunn, A.M. Superior predatory ability and abundance predicts potential ecological impact towards early-stage anurans by invasive ‘Killer Shrimp’ (Dikerogammarus villosus). Sci. Rep. 2021, 11, 4570. [Google Scholar] [CrossRef]
- Koester, M.; Bayer, B.; Gergs, R. Is Dikerogammarus villosus (Crustacea, Gammaridae) a ‘killer shrimp’ in the River Rhine system? Hydrobiologia 2016, 768, 299–313. [Google Scholar] [CrossRef]
- Maazouzi, C.; Piscart, C.; Pihan, J.C.; Masson, G. Effect of habitat-related resources on fatty acid composition and body weight of the invasive Dikerogammarus villosus in an artificial reservoir. Fundam. Appl. Limnol. 2009, 175, 327. [Google Scholar] [CrossRef]
- Copp, G.H.; Vilizzi, L.; Tidbury, H.; Stebbing, P.D.; Tarkan, A.S.; Miossec, L.; Goulletquer, P. Development of a generic decision-support tool for identifying potentially invasive aquatic taxa: AS-ISK. Manag. Biol. Invasions 2016, 7, 343–350. [Google Scholar] [CrossRef] [Green Version]
- D’hondt, B.; Vanderhoeven, S.; Roelandt, S.; Mayer, F.; Versteirt, V.; Adriaens, T.; Ducheyne, E.; Martin, G.S.; Grégoire, J.; Stiers, I.; et al. Harmonia+ and Pandora+: Risk screening tools for potentially invasive plants, animals and their pathogens. Biol. Invasions 2015, 17, 1869–1883. [Google Scholar] [CrossRef] [Green Version]
- Roy, H.E.; Rabitsch, W.; Scalera, R.; Stewart, A.; Gallardo, B.; Genovesi, P.; Essl, F.; Adriaens, T.; Bacher, S.; Booy, O.; et al. Developing a framework of minimum standards for the risk assessment of alien species. J. Appl. Ecol. 2018, 55, 526–538. [Google Scholar] [CrossRef] [Green Version]
- Srėbalienė, G.; Olenin, S.; Minchin, D.; Narščius, A. A comparison of impact and risk assessment methods based on the IMO Guidelines and EU invasive alien species risk assessment frameworks. PeerJ 2019, 7, e6965. [Google Scholar] [CrossRef] [Green Version]
- Nentwig, W.; Kuhnel, E.; Bacher, S. A generic impact-scoring system applied to alien mammals in Europe. Conserv. Biol. 2010, 24, 302–331. [Google Scholar] [CrossRef]
- Nentwig, W.; Bacher, S.; Pyšek, P.; Vilà, M.; Kumschick, S. The generic impact scoring system (GISS): A standardized tool to quantify the impacts of alien species. Environ. Monit. Assess. 2016, 188, 315. [Google Scholar] [CrossRef] [Green Version]
- Tarkan, A.S.; Emiroğlu, Ö.; Aksu, S.; Başkurt, S.; Aksu, İ.; Vilizzi, L.; Yoğurtçuoğlu, B. Coupling molecular and risk analysis to investigate the origin, distribution and potential impact of non-native species: An application to ruffe Gymnocephalus cernua in Turkey. Eur. Zool. J. 2022, 89, 102–114. [Google Scholar] [CrossRef]
- Voshell, J.R.; Laiton, R.J.; Hiner, S.W. Field techniques for determining the effect of toxic substances on benthic macroinvertebrates in rocky-bottomed streams. In Aquatic Toxicology and Hazard Assessment; Cowgill, U.M., Williams, L.R., Eds.; ASTM: Philadelphia, PA, USA, 1989; Volume 12, pp. 134–155. [Google Scholar]
- Dunn, O.J. Multiple comparisons using rank sums. Technometrics 1964, 6, 241–252. [Google Scholar] [CrossRef]
- Copp, G.H.; Vilizzi, L.; Wei, H.; Li, S.; Piria, M.; Al-Faisal, A.J.; Almeida, D.; Atique, U.; Al-Wazzan, Z.; Bakiu, R.; et al. Speaking their language—Development of a multilingual decision-support tool for communicating invasive species risks to decision makers and stakeholders. Environ. Model. Softw. 2021, 135, 104900. [Google Scholar] [CrossRef]
- Mastrandrea, M.D.; Field, C.B.; Stocker, T.F.; Edenhofer, O.; Ebi, K.L.; Frame, D.J.; Held, H.; Kriegler, E.; Mach, K.J.; Matschoss, P.R.; et al. Guidance Note for Lead Authors of the IPCC Fifth Assessment Report on Consistent Treatment of Uncertainties; Intergovernmental Panel on Climate Change (IPCC): Geneva, Switzerland, 2010. Available online: https://www.ipcc.ch/pdf/supporting-material/uncertainty-guidance-note.pdf (accessed on 29 September 2022).
- Vilizzi, L.; Copp, G.H.; Hill, J.E.; Adamovich, B.; Aislabie, L.; Akin, D.; Al-Faisal, A.J.; Almeida, D.; Azmai, M.N.A.; Bakiu, R.; et al. A global-scale screening of non-native aquatic organisms to identify potentially invasive species under current and future climate conditions. Sci. Total Environ. 2021, 788, 147868. [Google Scholar] [CrossRef] [PubMed]
- Lemmers, P.; Collas, F.P.L.; Gylstra, R.; Crombaghs, B.H.J.M.; van der Velde, G.; Leuven, R.S.E.W. Risks and management of alien freshwater crayfish species in the Rhine-Meuse river district. Manag. Biol. Invasions 2021, 12, 193–220. [Google Scholar] [CrossRef]
- Borza, P.; Huber, T.; Leitner, P.; Remund, N.; Graf, W. How to coexist with the ‘killer shrimp’ Dikerogammarus villosus? Lessons from other invasive Ponto-Caspian peracarids. Aquat. Conserv. Mar. Freshw. Ecosyst. 2018, 28, 1441–1450. [Google Scholar] [CrossRef] [Green Version]
- Grabowski, M.; Bącela, K.; Konopacka, A.; Jażdżewski, K. Salinity-related distribution of alien amphipods in rivers provides refugia for native species. Biol. Invasions 2009, 11, 2107–2117. [Google Scholar] [CrossRef]
- Grabowski, M.; Bacela, K.; Konopacka, A. How to be an invasive gammarid (Amphipoda: Gammaroidea)–comparison of life history traits. Hydrobiologia 2007, 590, 75–84. [Google Scholar] [CrossRef]
- Rewicz, T.; Grabowski, M.; MacNeil, C.; Bącela-Spychalska, K. The profile of a ‘perfect’ invader—The case of killer shrimp, Dikerogammarus villosus. Aquat. Invasions 2014, 9, 267–288. [Google Scholar] [CrossRef]
- Paganelli, D.; Kamburska, L.; Zaupa, S.; Garzoli, L.; Boggero, A. Impacts analysis of macroinvertebrate alien species in the hydrographic system of a subalpine lake on the Italian-Swiss border. Water 2021, 13, 3146. [Google Scholar] [CrossRef]
- Nentwig, W.; Bacher, S.; Kumschick, S.; Pyšek, P.; Vilà, M. More than “100 worst” alien species in Europe. Biol. Invasions 2018, 20, 1611–1621. [Google Scholar] [CrossRef]
- Wood, L.E.; Smith, E.R.C.; Bojko, J.; Stebbing, P. Options for the control of Dikerogammarus villosus (killer shrimp) and other invasive amphipods. Manag. Biol. Invasions 2021, 12, 662–684. [Google Scholar] [CrossRef]
- Balcombe, S.R.; Bunn, S.E.; McKenzie-Smith, F.J.; Davies, P.M. Variability of fish diets between dry and flood periods in an arid zone floodplain river. J. Fish Biol. 2005, 67, 1552–1567. [Google Scholar] [CrossRef] [Green Version]
- Bajer, P.G.; Ghosal, R.; Maselko, M.; Smanski, M.J.; Lechelt, J.D.; Hansen, G.; Kornis, M.S. Biological control of invasive fish and aquatic invertebrates: A brief review with case studies. Manag. Biol. Invasions 2019, 10, 227–254. [Google Scholar] [CrossRef] [Green Version]
- Kinzler, W.; Maier, G. Selective predation by fish: A further reason for the decline of native gammarids in the presence of invasives? J. Limnol 2006, 65, 27–34. [Google Scholar] [CrossRef]
- Kobak, J.; Jermacz, Ł.; Płąchocki, D. Effectiveness of zebra mussels to act as shelters from fish predators differs between native and invasive amphipod prey. Aquat. Ecol. 2014, 48, 397–408. [Google Scholar] [CrossRef] [Green Version]
- Ovcharenko, M.O.; Bacela, K.; Wilkinson, T.; Ironside, J.E.; Rigaud, T.; Wattier, R.A. Cucumispora dikerogammari n. gen. (Fungi: Microsporidia) infecting the invasive amphipod Dikerogammarus villosus: A potential emerging disease in European rivers. Parasitology 2010, 137, 191–204. [Google Scholar] [CrossRef] [Green Version]
- Bacela-Spychalska, K.; Wattier, R.A.; Genton, C.; Rigaud, T. Microsporidian disease of the invasive amphipod Dikerogammarus villosus and the potential for its transfer to local invertebrate fauna. Biol. Invasions 2012, 14, 1831–1842. [Google Scholar] [CrossRef]
- Freeman, M.A.; Turnbull, J.F.; Yeomans, W.E.; Bean, C.W. Prospects for management strategies of invasive crayfish populations with an emphasis on biological control. Aquat. Conserv. 2010, 20, 211–223. [Google Scholar] [CrossRef]
- Sanchez-Bayo, F.P. Insecticides mode of action in relation to their toxicity to non-target organisms. J. Environ. Anal. Toxicol. 2012, 2012, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Stebbing, P.; Irving, S.; Stentiford, G.; Mitchard, N. A Review of Potential Methods to Control and Eradicate the Invasive Gammarid, Dikerogammarus villosus from UK Waters; Cefas: Weymouth, UK, 2012; pp. 4–47.
- De Ventura, L.; Weissert, N.; Tobias, R.; Kopp, K.; Jokela, J. Identifying target factors for interventions to increase boat cleaning in order to prevent spread of invasive species. Manag. Biol. Invasions 2017, 8, 71–84. [Google Scholar] [CrossRef]
- Shannon, C.; Quinn, C.H.; Stebbing, P.D.; Hassall, C.; Dunn, A.M. The practical application of hot water to reduce the introduction and spread of aquatic invasive alien species. Manag. Biol. Invasions 2018, 9, 417–423. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Station | Water Temperature (°C) | O2 (mg L−1) | % of Saturation | Conductivity (µS m−1) | Main Substrates | Water Current |
|---|---|---|---|---|---|---|
| st01 | 19.9 | 9.59 | 105.7 | 241 | Pebbles with periphyton | moderate |
| st02 | 20.1 | 8.61 | 95 | 285 | Pebbles with periphyton | moderate |
| st03 | 20.4 | 10.41 | 115.4 | 328 | Pebbles with periphyton and sand | moderate |
| st04 | 20.9 | 9.67 | 108.3 | 349 | Stones, bryophytes, pebbles and sand | moderate |
| st05 | 21 | 11 | 123.4 | 374 | Small pebbles with periphyton | moderate |
| st06 | 21.5 | 9.82 | 111 | 432 | Small pebbles and sand | low |
| st07 | 23.2 | 7.69 | 89.8 | 359 | Mud and macrophytes | low |
| st08 | 21.9 | 8.83 | 100.6 | 454 | Small pebbles | low |
| Stations | st01 | st02 | st03 | st04 | st05 | st06 | st07 |
|---|---|---|---|---|---|---|---|
| st02 | 0.500 | ||||||
| st03 | 0.500 | 0.500 | |||||
| st04 | 0.001 *** | 0.001 *** | 0.001 *** | ||||
| st05 | 0.008 ** | 0.008 ** | 0.008 ** | 0.238 | |||
| st06 | 0.014 * | 0.014 * | 0.014 * | 0.1868 | 0.429 | ||
| st07 | 0.143 | 0.143 | 0.143 | 0.022 * | 0.096 | 0.129 | |
| st08 | 0.025 * | 0.025 * | 0.025 * | 0.129 | 0.339 | 0.406 | 0.186 |
| p-values | |||||||
| AS-ISK Partition | Score |
|---|---|
| Biogeography/Historical | 17.0 |
| 1. Domestication/Cultivation | 0.0 |
| 2. Climate, distribution and introduction risk | 3.0 |
| 3. Invasive elsewhere | 14.0 |
| Biology/Ecology | 23.0 |
| 4. Undesirable (or persistence) traits | 7.0 |
| 5. Resource exploitation | 7.0 |
| 6. Reproduction | 0.0 |
| 7. Dispersal mechanisms | 4.0 |
| 8. Tolerance attributes | 5.0 |
| Climate change | 4.0 |
| 9. Climate change | 4.0 |
| AS-ISK sector affected | |
| Commercial | 14.0 |
| Environmental | 14.0 |
| Species or population nuisance traits | 22.0 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Paganelli, D.; Cianci, M.A.; Marchini, A. Risk Screening and Distribution of the Invasive Amphipod Dikerogammarus villosus (Sowinsky, 1894) in the River Adda (Northern Italy). Diversity 2022, 14, 838. https://doi.org/10.3390/d14100838
Paganelli D, Cianci MA, Marchini A. Risk Screening and Distribution of the Invasive Amphipod Dikerogammarus villosus (Sowinsky, 1894) in the River Adda (Northern Italy). Diversity. 2022; 14(10):838. https://doi.org/10.3390/d14100838
Chicago/Turabian StylePaganelli, Daniele, Maria Angela Cianci, and Agnese Marchini. 2022. "Risk Screening and Distribution of the Invasive Amphipod Dikerogammarus villosus (Sowinsky, 1894) in the River Adda (Northern Italy)" Diversity 14, no. 10: 838. https://doi.org/10.3390/d14100838