
The Molecular Phylogeny of Land Plants: Progress and Future Prospects

{kind=link}
{kind=link}
Abstract
:1. Introduction
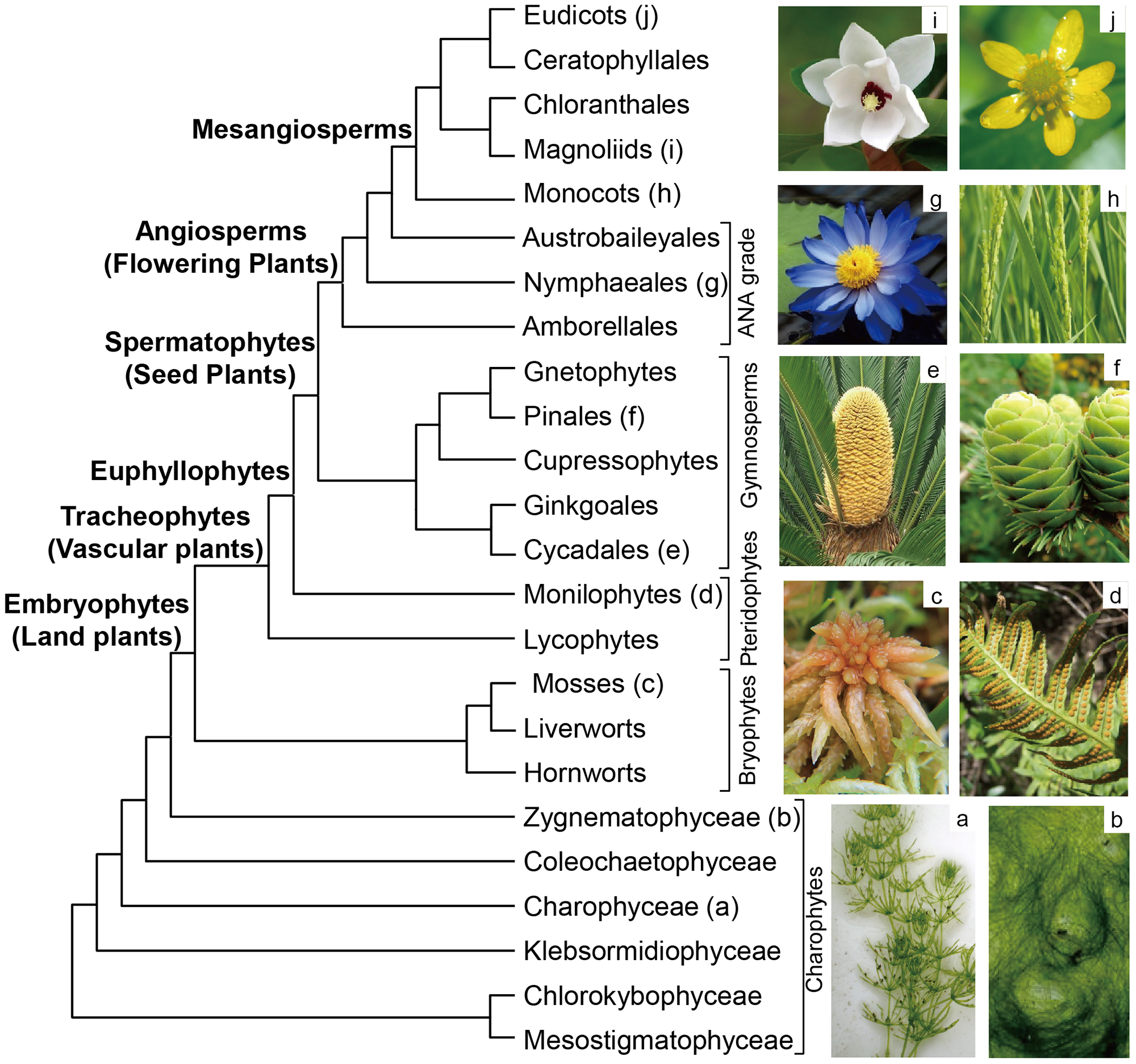
2. Large-Scale Phylogenetic Framework of Land Plants
2.1. Sister Group of Land Plants
2.2. Bryophytes
2.3. Pteridophytes
2.4. Gymnosperms
2.5. Angiosperms
3. The Potential Research Focuses on Reconstructing the TOL of Land Plants in the Long Period Future
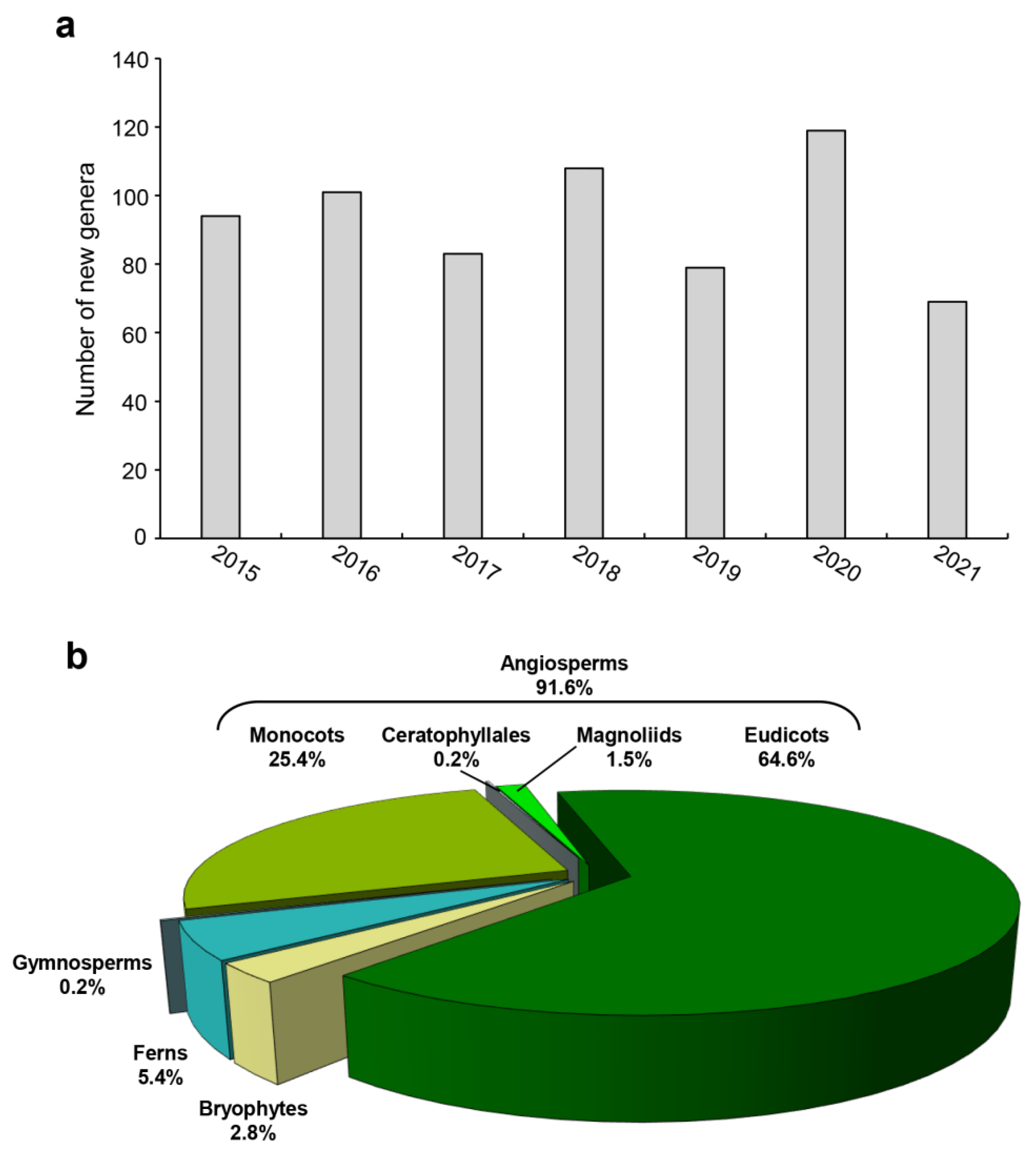
3.1. Reconstructing Genus- and Species-Level Phylogenies for Land Plant Groups
3.2. Updating the Classification Systems by Combining Morphological and Molecular Data
3.3. Integrating Fossil Taxa into Phylogenies Derived from Living Taxa
3.4. Resolve Deep-Level and/or Rapidly Divergent Phylogenetic Relationships using Phylogenomic Data
3.5. Building “Big Trees” Using the Supermatrix Method
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Darwin, C. The Origin of Species by Means of Natural Selection; John Murray: London, UK, 1859. [Google Scholar]
- Davis, C.C.; Matthews, S. Evolution of Land Plants; Oxford University Press: New York, NY, USA, 2019. [Google Scholar] [CrossRef]
- Wickett, N.J.; Mirarab, S.; Nguyen, N.; Warnow, T.; Carpenter, E.; Matasci, N.; Ayyampalayam, S.; Barker, M.S.; Burleigh, J.G.; Gitzendanner, M.A.; et al. Phylotranscriptomic analysis of the origin and early diversification of land plants. Proc. Natl. Acad. Sci. USA 2014, 111, E4859–E4868. [Google Scholar] [CrossRef]
- Su, D.; Yang, L.; Shi, X.; Ma, X.; Zhou, X.; Hedges, S.B.; Zhong, B. Large-scale phylogenomic analyses reveal the monophyly of bryophytes and neoproterozoic origin of land plants. Mol. Biol. Evol. 2021, 38, 3332–3344. [Google Scholar] [CrossRef] [PubMed]
- Donoghue, P.C.J.; Harrison, C.J.; Paps, J.; Schneider, H. The evolutionary emergence of land plants. Curr. Biol. 2021, 31, R1281–R1298. [Google Scholar] [CrossRef] [PubMed]
- Cheng, S.; Xian, W.; Fu, Y.; Marin, B.; Keller, J.; Wu, T.; Sun, W.; Li, X.; Xu, Y.; Zhang, Y.; et al. Genomes of subaerial Zygnematophyceae provide insights into land plant evolution. Cell 2019, 179, 1057–1067.e14. [Google Scholar] [CrossRef]
- Gitzendanner, M.A.; Soltis, P.S.; Wong, G.K.; Ruhfel, B.R.; Soltis, D.E. Plastid phylogenomic analysis of green plants: A billion years of evolutionary history. Am. J. Bot. 2018, 105, 291–301. [Google Scholar] [CrossRef]
- One Thousand Plant Transcriptomes Initiative. One thousand plant transcriptomes and the phylogenomics of green plants. Nature 2019, 574, 679–685. [Google Scholar] [CrossRef] [PubMed]
- Ran, J.H.; Shen, T.T.; Wang, M.M.; Wang, X.Q. Phylogenomic resolves the deep phylogeny of seed plants and indicates partial convergent or homoplastic evolution between Gnetales and angiosperms. Proc. R. Soc. B 2018, 285, 20181012. [Google Scholar] [CrossRef]
- The Pteridophyte Phylogeny Group. A community-derived classification for extant lycophytes and ferns. J. Syst. Evol. 2016, 54, 563–603. [Google Scholar] [CrossRef]
- The Angiosperm Phylogeny Group. An update of the Angiosperm Phylogeny Group classification for the orders and families of flowering plants: APG IV. Bot. J. Linn. Soc. 2016, 181, 1–20. [Google Scholar] [CrossRef] [Green Version]
- Yang, Y.; Ferguson, D.K.; Liu, B.; Mao, K.S.; Gao, L.M.; Zhang, S.Z.; Wan, T.; Rushforth, K.; Zhang, Z.X. Recent advances on phylogenomics of gymnosperms and a new classification. Plant Divers. 2022, 44, 340–350. [Google Scholar] [CrossRef]
- McCourt, R.M.; Delwiche, C.F.; Karol, K.G. Charophyte algae and land plant origins. Trends Ecol. Evol. 2004, 19, 661–666. [Google Scholar] [CrossRef]
- Karol, K.G.; McCourt, R.M.; Cimino, M.T.; Delwiche, C.F. The closest living relatives of land plants. Science 2001, 294, 2351–2353. [Google Scholar] [CrossRef] [PubMed]
- Timme, R.E.; Bachvaroff, T.R.; Delwiche, C.F. Broad phylogenomic sampling and the sister lineage of land plants. PLoS ONE 2012, 7, e29696. [Google Scholar] [CrossRef] [PubMed]
- Wodniok, S.; Brinkmann, H.; Glöckner, G.; Heidel, A.J.; Philippe, H.; Melkonian, M.; Becker, B. Origin of land plants: Do conjugating green algae hold the key? BMC Evol. Biol. 2011, 11, 104. [Google Scholar] [CrossRef]
- Turmel, M.; Otis, C.; Lemieux, C. The chloroplast genome sequence of Chara vulgaris sheds new light into the closest green algal relatives of land plants. Mol. Biol. Evol. 2006, 23, 1324–1338. [Google Scholar] [CrossRef]
- Finet, C.; Timme, R.E.; Delwiche, C.F.; Marlétaz, F. Multigene phylogeny of the green lineage reveals the origin and diversification of land plants. Curr. Biol. 2012, 22, 1456–1457. [Google Scholar] [CrossRef]
- Laurin-Lemay, S.; Brinkmann, H.; Philippe, H. Origin of land plants revisited in the light of sequence contamination and missing data. Curr. Biol. 2012, 22, R593–R594. [Google Scholar] [CrossRef]
- Pires, N.D.; Dolan, L. Morphological evolution in land plants: New designs with old genes. Philos. Trans. R. Soc. B 2012, 367, 508–518. [Google Scholar] [CrossRef]
- Jill Harrison, C. Development and genetics in the evolution of land plant body plans. Philos. Trans. R. Soc. B 2017, 372, 20150490. [Google Scholar] [CrossRef] [Green Version]
- Ishizaki, K. Evolution of land plants: Insights from molecular studies on basal lineages. Biosci. Biotechnol. Biochem. 2017, 81, 73–80. [Google Scholar] [CrossRef] [PubMed]
- Szövényi, P.; Waller, M.; Kirbis, A. Evolution of the plant body plan. Curr. Top. Dev. Biol. 2019, 131, 1–34. [Google Scholar] [CrossRef]
- Wang, Q.H.; Zhang, J.; Liu, Y.; Jia, Y.; Jiao, Y.N.; Xu, B.; Chen, Z.D. Diversity, phylogeny, and adaptation of bryophytes: Insights from genomic and transcriptomic data. J. Exp. Bot. 2022, 73, 4306–4322. [Google Scholar] [CrossRef]
- Qiu, Y.L.; Cho, Y.; Cox, C.; Palmer, J.D. The gain of three mitochondrial introns identifies liverworts as the earliest land plants. Nature 1998, 394, 671–674. [Google Scholar] [CrossRef]
- Qiu, Y.L.; Li, L.B.; Wang, B.; Chen, Z.D.; Knoop, V.; Groth-Malonek, M.; Dombrovska, O.; Lee, J.; Kent, L.; Rest, J.; et al. The deepest divergences in land plants inferred from phylogenomic evidence. Proc. Natl. Acad. Sci. USA 2006, 103, 15511–15516. [Google Scholar] [CrossRef]
- Chang, Y.; Graham, S.W. Bryophyte-specific primers for retrieving plastid genes suitable for phylogenetic inference. Am. J. Bot. 2011, 98, e109–e113. [Google Scholar] [CrossRef]
- Liu, Y.; Cox, C.J.; Wang, W.; Goffinet, B. Mitochondrial phylogenomics of early land plants: Mitigating the effects of saturation, compositional heterogeneity, and codon-usage bias. Syst. Biol. 2014, 63, 862–878. [Google Scholar] [CrossRef]
- Cox, C.J.; Li, B.; Foster, P.G.; Embley, T.M.; Civan, P. Conflicting phylogenies for early land plants are caused by composition biases among synonymous substitutions. Syst. Biol. 2014, 63, 272–279. [Google Scholar] [CrossRef]
- Zhang, J.; Fu, X.X.; Li, R.Q.; Zhao, X.; Liu, Y.; Li, M.H.; Zwaenepoel, A.; Ma, H.; Goffinet, B.; Guan, Y.L.; et al. The hornwort genome and early land plant evolution. Nat. Plants 2020, 6, 107–118. [Google Scholar] [CrossRef]
- De Sousa, F.; Foster, P.G.; Donoghue, P.C.J.; Schneider, H.; Cox, C.J. Nuclear protein phylogenies support the monophyly of the three bryophyte groups (Bryophyta Schimp.). New Phytol. 2019, 222, 565–575. [Google Scholar] [CrossRef]
- Kranz, H.D.; Huss, V.A.R. Molecular evolution of pteridophytes and their relationship to seed plants: Evidence from complete 18S rRNA gene sequences. Plant Syst. Evol. 1996, 202, 1–11. [Google Scholar] [CrossRef]
- Pryer, K.M.; Schneider, H.; Smith, A.R.; Cranfill, R.; Wolf, P.G.; Hunt, J.S.; Sipes, S.D. Horsetails and ferns are a monophyletic group and the closest living relatives to seed plants. Nature 2001, 409, 618–622. [Google Scholar] [CrossRef] [PubMed]
- Rai, H.S.; Graham, S.W. Utility of a large, multigene plastid data set in inferring higher-order relationships in ferns and relatives (monilophytes). Am. J. Bot. 2010, 97, 1444–1456. [Google Scholar] [CrossRef] [PubMed]
- Bowe, L.M.; Coat, G.; DePamphilis, C.W. Phylogeny of seed plants based on all three genomic compartments: Extant gymnosperms are monophyletic and Gnetales’ closest relatives are conifers. Proc. Natl. Acad. Sci. USA 2000, 97, 4092–4097. [Google Scholar] [CrossRef]
- Chaw, S.M.; Parkinson, C.L.; Cheng, Y.C.; Vincent, T.M.; Palmer, J.D. Seed plant phylogeny inferred from all three plant genomes: Monophyly of extant gymnosperms and origin of Gnetales from conifers. Proc. Natl. Acad. Sci. USA 2000, 97, 4086–4091. [Google Scholar] [CrossRef]
- Liu, Y.; Wang, S.; Li, L.; Yang, T.; Dong, S.; Wei, T.; Wu, S.; Liu, Y.; Gong, Y.; Feng, X.; et al. The Cycas genome and the early evolution of seed plants. Nat. Plants 2022, 8, 389–401. [Google Scholar] [CrossRef]
- Wan, T.; Liu, Z.M.; Li, L.F.; Leitch, A.R.; Leitch, I.J.; Lohaus, R.; Liu, Z.J.; Xin, H.P.; Gong, Y.B.; Liu, Y.; et al. A genome for gnetophytes and early evolution of seed plants. Nat. Plants 2018, 4, 82–89. [Google Scholar] [CrossRef]
- Parkinson, C.L.; Adams, K.L.; Palmer, J.D. Multigene analyses identify the three earliest lineages of extant flowering plants. Curr. Biol. 1999, 9, 1485–1491. [Google Scholar] [CrossRef]
- Saarela, J.M.; Rai, H.S.; Doyle, J.A.; Endress, P.K.; Mathews, S.; Marchant, A.D.; Briggs, B.G.; Graham, S.W. Hydatellaceae identified as a new branch near the base of the angiosperm phylogenetic tree. Nature 2007, 446, 312–315. [Google Scholar] [CrossRef]
- Qiu, Y.L.; Dombrovska, O.; Lee, J.; Li, L.; Whitlock, B.A.; Bernasconi-Quadroni, F.; Rest, J.S.; Davis, C.C.; Borsch, T.; Hilu, K.W.; et al. Phylogenetic analyses of basal angiosperms based on nine plastid, mitochondrial, and nuclear genes. Int. J. Plant Sci. 2005, 166, 815–842. [Google Scholar] [CrossRef] [Green Version]
- Amborella Genome Project; Albert, V.A.; Barbazuk, W.B.; Depamphilis, C.W.; Der, J.P.; Leebens-Mack, J.; Ma, H.; Palmer, J.D.; Rounsley, S.; Sankoff, D.; et al. The Amborella genome and the evolution of flowering plants. Science 2013, 342, 1241089. [Google Scholar] [CrossRef]
- Zhang, L.; Chen, F.; Zhang, X.; Li, Z.; Zhao, Y.; Lohaus, R.; Chang, X.; Dong, W.; Ho, S.Y.W.; Liu, X.; et al. The water lily genome and the early evolution of flowering plants. Nature 2020, 577, 79–84. [Google Scholar] [CrossRef] [PubMed]
- Moore, M.J.; Bell, C.D.; Soltis, P.S.; Soltis, D.E. Using plastid genome-scale data to resolve enigmatic relationships among basal angiosperms. Proc. Natl. Acad. Sci. USA 2007, 104, 19363–19368. [Google Scholar] [CrossRef] [PubMed]
- Ma, J.; Sun, P.; Wang, D.; Wang, Z.; Yang, J.; Li, Y.; Mu, W.; Xu, R.; Wu, Y.; Dong, C.; et al. The Chloranthus sessilifolius genome provides insight into early diversification of angiosperms. Nat. Commun. 2021, 12, 6929. [Google Scholar] [CrossRef] [PubMed]
- Guo, X.; Fang, D.; Sahu, S.K.; Yang, S.; Guang, X.; Folk, R.; Smith, S.A.; Chanderbali, A.S.; Chen, S.; Liu, M.; et al. Chloranthus genome provides insights into the early diversification of angiosperms. Nat. Commun. 2021, 12, 6930. [Google Scholar] [CrossRef]
- Chase, M.; Soltis, D.E.; Olmstead, R.G.; Morgan, D.; Les, D.H.; Mishler, B.D.; Duvall, M.R.; Price, R.A.; Hills, H.G.; Qiu, Y.L.; et al. Phylogenetics of seed plants: An analysis of nucleotide sequences from the plastid gene rbcL. Ann. Mo. Bot. Gard. 1993, 80, 528–580. [Google Scholar] [CrossRef]
- Soltis, D.E.; Moore, M.J.; Burleigh, G.; Soltis, P.S. Molecular markers and concepts of plant evolutionary relationships: Progress, promise, and future prospects. Crit. Rev. Plant Sci. 2009, 28, 1–15. [Google Scholar] [CrossRef]
- The Angiosperm Phylogeny Group. An ordinal classification for the families of flowering plants. Ann. Mo. Bot. Gard. 1998, 85, 531–553. [Google Scholar] [CrossRef]
- The Angiosperm Phylogeny Group. An update of the Angiosperm Phylogeny Group classification for the orders and families of flowering plants: APG II. Bot. J. Linn. Soc. 2003, 141, 399–436. [Google Scholar] [CrossRef]
- The Angiosperm Phylogeny Group. An update of the Angiosperm Phylogeny Group classification for the orders and families of flowering plants: APG III. Bot. J. Linn. Soc. 2009, 161, 105–121. [Google Scholar] [CrossRef] [Green Version]
- Lehtonen, S. Towards resolving the complete fern tree of life. PLoS One 2011, 6, e24851. [Google Scholar] [CrossRef]
- Liu, Y.; Johnson, M.G.; Cox, C.J.; Medina, R.; Devos, N.; Vanderpoorten, A.; Hedenäs, L.; Bell, N.E.; Shevock, J.R.; Aguero, B.; et al. Resolution of the ordinal phylogeny of mosses using targeted exons from organellar and nuclear genomes. Nat. Commun. 2019, 10, 1485. [Google Scholar] [CrossRef] [PubMed]
- Hinchliff, C.E.; Smith, S.A.; Allman, J.F.; Burleigh, J.G.; Chaudhary, R.; Coghill, L.M.; Crandall, K.A.; Deng, J.; Drew, B.T.; Gazis, R.; et al. Synthesis of phylogeny and taxonomy into a comprehensive tree of life. Proc. Natl. Acad. Sci. USA 2015, 112, 12764–12769. [Google Scholar] [CrossRef] [PubMed]
- Folk, R.A.; Sun, M.; Soltis, P.S.; Smith, S.A.; Soltis, D.E.; Guralnick, R.P. Challenges of comprehensive taxon sampling in comparative biology: Wrestling with rosids. Am. J. Bot. 2018, 105, 433–445. [Google Scholar] [CrossRef]
- Liu, H.-M.; Jiang, R.-H.; Guo, J.; Hovenkamp, P.; Perrie, L.R.; Shepherd, L.; Hennequin, S.; Schneider, H. Towards a phylogenetic classification of the climbing fern genus Arthropteris. Taxon 2013, 62, 688–700. [Google Scholar] [CrossRef]
- Zhou, X.M.; Zhang, L.; Lu, N.T.; Gao, X.F.; Zhang, L.B. Pteridryaceae: A new fern family of Polypodiineae (Polypodiales) including taxonomic treatments. J. Syst. Evol. 2018, 56, 148–173. [Google Scholar] [CrossRef]
- Su, J.X.; Wang, W.; Zhang, L.B.; Chen, Z.D. Phylogenetic placement of two enigmatic genera, Borthwickia and Stixis, based on molecular and pollen data, and the description of a new family of Brassicales, Borthwickiaceae. Taxon 2012, 61, 601–611. [Google Scholar] [CrossRef]
- Liu, B.; Tan, Y.H.; Liu, S.; Olmstead, R.G.; Min, D.Z.; Chen, Z.D.; Joshee, N.; Vaidya, B.N.; Chung, R.C.K.; Li, B. Phylogenetic relationships of Cyrtandromoea and Wightia revisited: A new tribe in Phrymaceae and a new family in Lamiales. J. Syst. Evol. 2019, 58, 1–17. [Google Scholar] [CrossRef]
- Wang, W.; Liu, Y.; Yu, S.X.; Gao, T.G.; Chen, Z.D. Gymnaconitum, a new genus of Ranunculaceae endemic to the Qinghai–Tibetan Plateau. Taxon 2013, 62, 713–722. [Google Scholar] [CrossRef]
- Kadereit, J.W.; Albach, D.C.; Ehrendorfer, F.; Galbany-Casals, M.; Garcia-Jacas, N.; Gehrke, B.; Kadereit, G.; Kilian, N.; Klein, J.T.; Koch, M.A.; et al. Which changes are needed to render all genera of the German flora monophyletic? Willdenowia 2016, 46, 39–91. [Google Scholar] [CrossRef] [Green Version]
- Wang, W.; Zhang, X.X.; Chen, Z.D.; Lu, A.M. Comments on the APG’s classification of angiosperms. Biodivers. Sci. 2017, 25, 418–426. [Google Scholar] [CrossRef]
- Nandi, O.; Chase, M.W.; Endress, P.K. A combined cladistic analysis of angiosperms using rbcL and non-molecular data sets. Ann. Mo. Bot. Gard. 1998, 85, 137–214. [Google Scholar] [CrossRef]
- Doyle, J.A.; Endress, P.K. Morphological phylogenetic analysis of basal angiosperms: Comparison and combination with molecular data. Int. J. Plant Sci. 2000, 161, S121–S153. [Google Scholar] [CrossRef]
- Soltis, D.E.; Moore, M.J.; Sessa, E.B.; Smith, S.A.; Soltis, P.S. Using and navigating the plant tree of life. Am. J. Bot. 2018, 105, 287–290. [Google Scholar] [CrossRef] [PubMed]
- Glenner, H.; Hansen, A.J.; Sørensen, M.V.; Ronquist, F.; Huelsenbeck, J.P.; Willerslev, E. Bayesian inference of the metazoan phylogeny: A combined molecular and morphological approach. Curr. Biol. 2004, 14, 1644–1649. [Google Scholar] [CrossRef] [PubMed]
- O’Reilly, J.E.; Puttick, M.N.; Pisani, D.; Donoghue, P.C.J. Probabilistic methods surpass parsimony when assessing clade support in phylogenetic analyses of discrete morphological data. Palaeontology 2018, 61, 105–118. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Lu, A.M.; Ren, Y.; Endress, M.E.; Chen, Z.D. Phylogeny and classification of Ranunculales: Evidence from four molecular loci and morphological data. Perspect. Plant Ecol. Evol. Syst. 2009, 11, 81–110. [Google Scholar] [CrossRef]
- Yu, S.X.; Janssens, S.B.; Zhu, X.Y.; Lidén, M.; Gao, T.G.; Wang, W. Phylogeny of Impatiens (Balsaminaceae): Integrating molecular and morphological evidence into a new classification. Cladistics 2016, 32, 179–197. [Google Scholar] [CrossRef] [PubMed]
- Friis, E.M.; Pedersen, K.R.; Crane, P.R. The emergence of core eudicots: New floral evidence from the earliest Late Cretaceous. Proc. R. Soc. B 2016, 283, 20161325. [Google Scholar] [CrossRef]
- Christiansen, P. Phylogeny of the great cats (Felidae: Pantherinae), and the influence of fossil taxa and missing characters. Cladistics 2008, 24, 977–992. [Google Scholar] [CrossRef]
- Parham, J.F.; Donoghue, P.C.; Bell, C.J.; Calway, T.D.; Head, J.J.; Holroyd, P.A.; Inoue, J.G.; Irmis, R.B.; Joyce, W.G.; Ksepka, D.T.; et al. Best practices for justifying fossil calibrations. Syst. Biol. 2012, 61, 346–359. [Google Scholar] [CrossRef]
- Xiang, X.; Xiang, K.; Ortiz, R.D.C.; Jabbour, F.; Wang, W. Integrating palaeontological and molecular data uncovers multiple ancient and recent dispersals in the pantropical Hamamelidaceae. J. Biogeogr. 2019, 46, 2622–2631. [Google Scholar] [CrossRef]
- Pyron, R.A. Divergence time estimation using fossils as terminal taxa and the origins of Lissamphibia. Syst. Biol. 2011, 60, 466–481. [Google Scholar] [CrossRef] [PubMed]
- Palmer, J.D.; Jansen, R.K.; Michaels, H.J.; Chase, M.W.; Manhart, J.R. Chloroplast DNA variation and plant phylogeny. Ann. Mo. Bot. Gard. 1988, 75, 1180–1206. [Google Scholar] [CrossRef]
- Goremykin, V.V.; Nikiforova, S.V.; Biggs, P.J.; Zhong, B.; Delange, P.; Martin, W.; Woetzel, S.; Atherton, R.A.; McLenachan, P.A.; Lockhart, P.J. The evolutionary root of flowering plants. Syst. Biol. 2013, 62, 50–61. [Google Scholar] [CrossRef]
- Drew, B.T.; Ruhfel, B.R.; Smith, S.A.; Moore, M.J.; Briggs, B.G.; Gitzendanner, M.A.; Soltis, P.S.; Soltis, D.E. Another look at the root of the angiosperms reveals a familiar tale. Syst. Biol. 2014, 63, 368–382. [Google Scholar] [CrossRef]
- Xi, Z.; Ruhfel, B.R.; Schaefer, H.; Amorim, A.M.; Sugumaran, M.; Wurdack, K.J.; Endress, P.K.; Matthews, M.L.; Stevens, P.F.; Mathews, S.; et al. Phylogenomics and a posteriori data partitioning resolve the Cretaceous angiosperm radiation Malpighiales. Proc. Natl. Acad. Sci. USA 2012, 109, 17519–17524. [Google Scholar] [CrossRef]
- McKain, M.R.; Johnson, M.G.; Uribe-Convers, S.; Eaton, D.; Yang, Y. Practical considerations for plant phylogenomics. Appl. Plant Sci. 2018, 6, e1038. [Google Scholar] [CrossRef]
- Smith, S.A.; Beaulieu, J.M.; Donoghue, M.J. Mega-phylogeny approach for comparative biology: An alternative to supertree and supermatrix approaches. BMC Evol. Biol. 2009, 9, 37. [Google Scholar] [CrossRef]
- Smith, S.A.; Brown, J.W.; Yang, Y.; Bruenn, R.; Drummond, C.P.; Brockington, S.F.; Walker, J.F.; Last, N.; Douglas, N.A.; Moore, M.J. Disparity, diversity, and duplications in the Caryophyllales. New Phytol. 2018, 217, 836–854. [Google Scholar] [CrossRef] [Green Version]
- Thornhill, A.H.; Baldwin, B.G.; Freyman, W.A.; Nosratinia, S.; Kling, M.M.; Morueta-Holme, N.; Madsen, T.P.; Ackerly, D.D.; Mishler, B.D. Spatial phylogenetics of the native California flora. BMC Biol. 2017, 15, 96. [Google Scholar] [CrossRef]
- Wu, S.D.; Zhang, L.J.; Lin, L.; Yu, S.X.; Chen, Z.D.; Wang, W. Insights into the historical assembly of global dryland floras: The diversification of Zygophyllaceae. BMC Evol. Biol. 2018, 18, 166. [Google Scholar] [CrossRef]
- Eiserhardt, W.L.; Antonelli, A.; Bennett, D.J.; Botigué, L.R.; Burleigh, J.G.; Dodsworth, S.; Enquist, B.J.; Forest, F.; Kim, J.T.; Kozlov, A.M.; et al. A roadmap for global synthesis of the plant tree of life. Am. J. Bot. 2018, 105, 614–622. [Google Scholar] [CrossRef] [Green Version]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, G.-Q.; Lian, L.; Wang, W. The Molecular Phylogeny of Land Plants: Progress and Future Prospects. Diversity 2022, 14, 782. https://doi.org/10.3390/d14100782
Liu G-Q, Lian L, Wang W. The Molecular Phylogeny of Land Plants: Progress and Future Prospects. Diversity. 2022; 14(10):782. https://doi.org/10.3390/d14100782
Chicago/Turabian StyleLiu, Guo-Qing, Lian Lian, and Wei Wang. 2022. "The Molecular Phylogeny of Land Plants: Progress and Future Prospects" Diversity 14, no. 10: 782. https://doi.org/10.3390/d14100782