
Consistent Grime’s CSR Strategy of Daisy Fleabane Erigeron annuus (L.) Pers. despite Its High Morphological Variability—A Case Study from Zagreb and Medvednica Mt., Croatia



Abstract
:1. Introduction
2. Materials and Methods
2.1. Population Sampling
2.2. Measuring Life Traits
2.3. Determining CSR Strategies
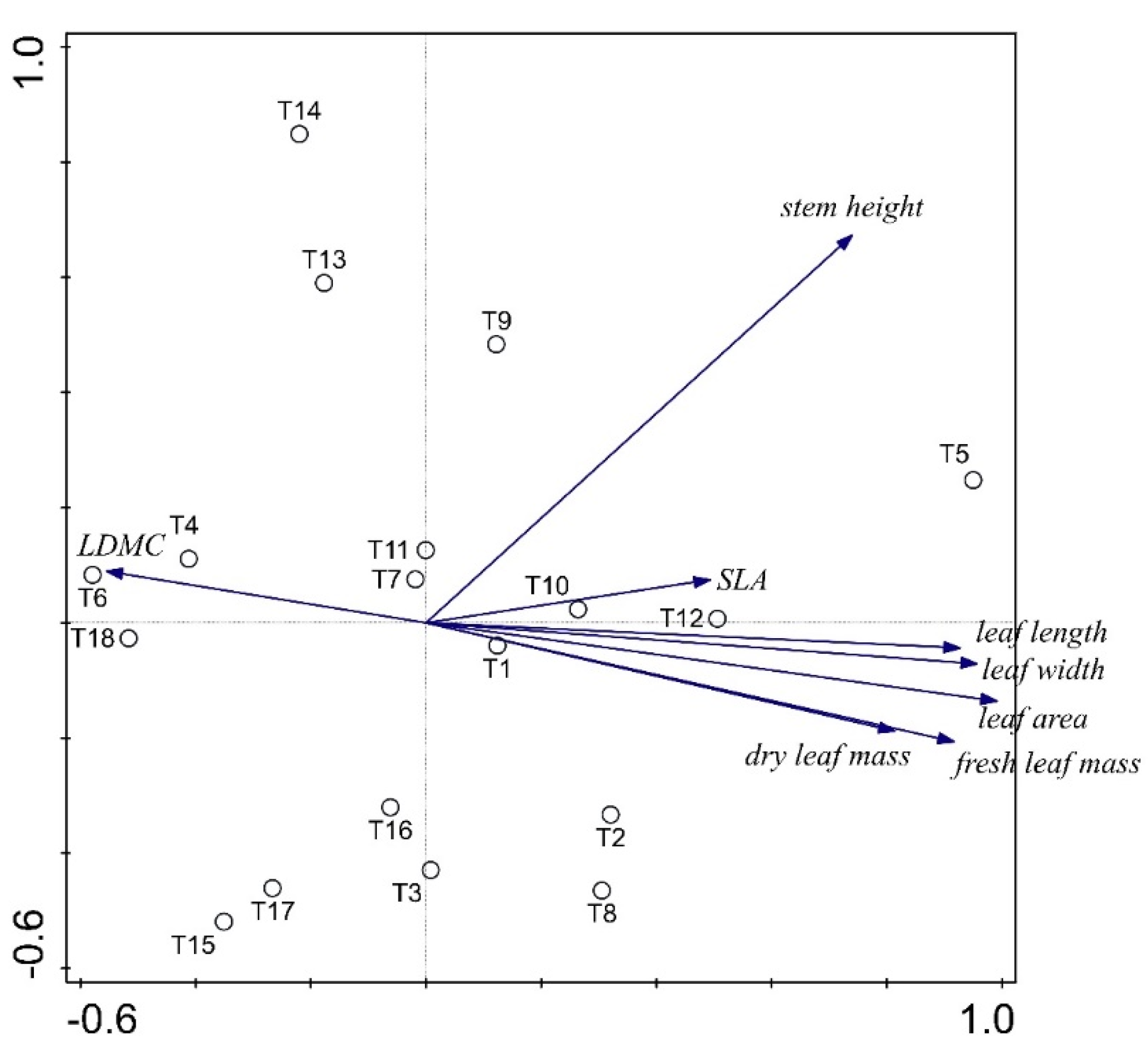
2.4. Statistical Analyses
3. Results
3.1. Measurements of Life Traits
3.2. Determining CSR Strategies
3.3. Comparison of Life Traits Variability between Sampled Populations
3.4. Correlations of Life Traits with Environmental Variables
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Thompson, K.; Hodgson, J.G.; Rich, T.C.G. Native and alien invasive plants: More of the same? Ecography 1995, 18, 390–402. [Google Scholar] [CrossRef]
- Kühn, I.; Klotz, S. Urbanization and homogenization—Comparing the floras of urban and rural areas in Germany. Biol. Conserv. 2006, 127, 292–300. [Google Scholar] [CrossRef]
- Chytrý, M.; Pyšek, P.; Wild, J.; Pino, J.; Maskell, L.C.; Vilà, M. European map of alien plant invasions based on the quantitative assessment across habitats. Divers. Distrib. 2008, 15, 98–107. [Google Scholar] [CrossRef]
- Nikolić, T.; Mitić, B.; Milašinović, B.; Jelaska, S.D. Invasive vascular plants in Croatia: Distirbution pattern, range size and ecology. In Book of Abstracts of the 11th Croatian Biological Congress, Proceedings of 11th Croatian Biological Congress with International Participation, Šibenik, Croatia, 16–21 September 2012; Jelaska, S.D., Klobučar, G.I.V., Šerić Jelaska, L., Leljak Levanić, D., Lukša, Ž., Eds.; Croatian Biological Society: Zagreb, Croatia, 2012. [Google Scholar]
- IUCN. Red List Categories and Criteria. Version 14. Available online: http://www.iucnredlist.org/documents/RedListGuidelines.pdf (accessed on 21 July 2021).
- Hejda, M.; Pyšek, P.; Jarošík, V. Impact of invasive plants on the species richness, diversity and composition of invaded communities. J. Ecol. 2009, 97, 393–403. [Google Scholar] [CrossRef]
- Nikolić, T.; Mitić, B.; Boršić, I. Flora Hrvatske: Invazivne Biljke, 1st ed.; Alfa d.o.o.: Zagreb, Croatia, 2014; pp. 1–296. [Google Scholar]
- Trtikova, M.; Güsewell, S.; Baltisberger, M.; Edwards, P.J. Distribution, growth performance and genetic variation of Erigeron annuus in the Swiss Alps. Biol. Invasions 2010, 13, 413–422. [Google Scholar] [CrossRef]
- Trtikova, M. Effects of competition and mowing on growth and reproduction of the invasive plant Erigeron annuus at two contrasting altitudes. Bot. Helvetica 2009, 119, 1–6. [Google Scholar] [CrossRef]
- Vuković, N.; Pavičević, M.; Jelaska, S.D. Allelopatic effects of five invasive species on germination and seedling growth of Sinapis alba L. and Triticum aestivum L. In Book of Abstracts of the 11th Croatian Biological Congress, Proceedings of 11th Croatian Biological Congress with International Participation, Šibenik, Croatia, 16–21 September 2012; Jelaska, S.D., Klobučar, G.I.V., Šerić Jelaska, L., Leljak Levanić, D., Lukša, Ž., Eds.; Croatian Biological Society: Zagreb, Croatia, 2012. [Google Scholar]
- Flora Croatica Database Allochthonous Plants. Available online: https://hirc.botanic.hr/fcd/InvazivneVrste/ShowResults.aspx?hash=636268493 (accessed on 21 July 2021).
- Veenvliet, J.K.; Veenvliet, J.P.; de Groot, M.; Kutnar, L. A Field Guide to Invasive Alien Species in European Forests, 1st ed.; Institute Symbiosis, so. e. and The Silva Slovenica Publishing Centre, Slovenian Forestry Institute: Ljubljana, Slovenia, 2019; pp. 128–129. [Google Scholar]
- Stratton, D.A. Life-cycle components of selection in Erigeron annuus: I. Phenotypic selection. Evolution 1992, 46, 92–106. [Google Scholar] [CrossRef]
- Trtikova, M.; Edwards, P.J.; Güsewell, S. No adaptation to altitude in the invasive plantErigeron annuusin the Swiss Alps. Ecography 2010, 33, 556–564. [Google Scholar] [CrossRef]
- Halliday, G.; Erigeron, L. Flora Europaea 4, 7th ed.; Tutin, T.G., Heywood, V.H., Burges, N.A., Moore, D.M., Valentine, D.H., Walters, S.M., Webb, D.A., Chater, A.O., DeFilipps, R.A., Richardson, I.B.K., Eds.; Cambridge University Press: New York, NY, USA, 1976; Volume 4, pp. 116–120. [Google Scholar]
- Becker, T.; Dietz, H.; Billeter, R.; Buschmann, H.; Edwards, P.J. Altitudinal distribution of alien plant species in the Swiss Alps. Perspect. Plant Ecol. Evol. Syst. 2005, 7, 173–183. [Google Scholar] [CrossRef]
- Klotz, S.; Kühn, I.; Durka, W. BIOLFLOR- Eine Datenbank zu Biologisch-Ökologischen Merkmalen der Gefäßpflanzen in Deutschland; Schriftenreihe für Vegetationskunde: Bonn, Germany, 2002; pp. 1–5. [Google Scholar]
- Vuković, N. Ecogeography of the Invasive Flora of Croatia. Ph.D. Thesis, University of Zagreb, Faculty of Science, Zagreb, Croatia, 2015. [Google Scholar]
- Bradshaw, A. Evolutionary Significance of Phenotypic Plasticity in Plants. In Advances in Genetics; Caspari, E.W., Thoday, J.M., Eds.; Academic Press: San Diego, CA, USA, 1965; Volume 13, pp. 115–155. [Google Scholar] [CrossRef]
- Schöb, C.; Armas, C.; Guler, M.; Prieto, I.; Pugnaire, F.I. Variability in functional traits mediates plant interactions along stress gradients. J. Ecol. 2013, 101, 753–762. [Google Scholar] [CrossRef]
- Niinemets, Ü.; Valladares, F.; Ceulemans, R. Leaf-level phenotypic variability and plasticity of invasive Rhododendron ponticum and non-invasive Ilex aquifolium co-occurring at two contrasting European sites. Plant Cell Environ. 2003, 26, 941–956. [Google Scholar] [CrossRef] [Green Version]
- Pérez-Harguindeguy, N.; Díaz, S.; Garnier, E.; Lavorel, S.; Poorter, H.; Jaureguiberry, P.; Bret-Harte, M.S.; Cornwell, W.K.; Craine, J.M.; Gurvich, D.E.; et al. New handbook for standardised measurement of plant functional traits worldwide. Aust. J. Bot. 2013, 61, 167–234. [Google Scholar] [CrossRef]
- Grime, J.P. Evidence for the Existence of Three Primary Strategies in Plants and Its Relevance to Ecological and Evolutionary Theory. Am. Nat. 1977, 111, 1169–1194. [Google Scholar] [CrossRef]
- Hodgson, J.G.; Wilson, P.J.; Hunt, R.; Grime, J.P.; Thompson, K. Allocating C-S-R Plant Functional Types: A Soft Approach to a Hard Problem. Oikos 1999, 85, 282. [Google Scholar] [CrossRef]
- Grime, J.P. Plant Strategies and Vegetation Processes; John Wiley & Sons, Ltd.: Hoboken, NJ, USA, 1979; pp. 1–222. [Google Scholar]
- Grime, J.P. Manipulation of plant species and communities. In Ecology and Design in Landscape, 1st ed.; Bradshaw, A.D., Goode, D.A., Thorpe, E., Eds.; Backwell Scientific Publications: Oxford, UK, 1986; pp. 175–194. [Google Scholar]
- Grime, J.P.; Thompson, K.; Hunt, R.; Hodgson, J.G.; Cornelissen, J.H.C.; Rorison, I.H.; Hendry, G.A.F.; Ashenden, T.W.; Askew, A.P.; Band, S.R.; et al. Integrated Screening Validates Primary Axes of Specialisation in Plants. Oikos 1997, 79, 259. [Google Scholar] [CrossRef]
- Westoby, M. A leaf-height-seed (LHS) plant ecology strategy scheme. Plant Soil 1998, 199, 213–227. [Google Scholar] [CrossRef]
- Pierce, S.; Brusa, G.; Vagge, I.; Cerabolini, B.E.L. Allocating CSR plant functional types: The use of leaf economics and size traits to classify woody and herbaceous vascular plants. Funct. Ecol. 2013, 27, 1002–1010. [Google Scholar] [CrossRef] [Green Version]
- Zhao, Y.-T.; Ali, A.; Yan, E.-R. The plant economics spectrum is structured by leaf habits and growth forms across subtropical species. Tree Physiol. 2016, 37, 173–185. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vuković, N.; Miletić, M.; Milović, M.; Jelaska, S.D. Grime’s CSR strategies of the invasive plants in Croatia. Period. Biol. 2014, 116, 323–329. [Google Scholar]
- Hunt, R.; Hodgson, J.; Thompson, K.; Bungener, P.; Dunnett, N.; Askew, A. A new practical tool for deriving a functional signature for herbaceous vegetation. Appl. Veg. Sci. 2004, 7, 163–170. [Google Scholar] [CrossRef]
- Knevel, I.C.; Bekker, R.M.; Kunzmann, D.; Stadler, M.; Thompson, K. The LEDA Traitbase Collecting and Measuring Standards of Life—History Traits of the Northern European Flora, 1st ed.; University of Groningen: Groningen, The Netherlands, 2005; pp. 49–60. [Google Scholar]
- Frey, D. Patterns of Variation within the Erigeron annuus Complex in the United States and Europe. Ph.D. Thesis, Swiss Federal Institute of Technology, Zürich, Switzerland, 2003. [Google Scholar]
- Lambdon, P.W.; Lloret, F.; Hulme, P.E. Do alien plants on Mediterranean islands tend to invade different niches from native species? Biol. Invasions 2007, 10, 703–716. [Google Scholar] [CrossRef]
- Meiners, S.J.; Pickett, S.T.A.; Cadenasso, M.L. Effects of plant invasions on the species richness of abandoned agricultural land. Ecography 2001, 24, 633–644. [Google Scholar] [CrossRef]
- Therriault, T.W.; Kolasa, J. Explicit links among physical stress, habitat heterogeneity and biodiversity. Oikos 2000, 89, 387–391. [Google Scholar] [CrossRef] [Green Version]
- Sennikov, A.N.; Kurtto, A. The taxonomy and invasion status assessment of Erigeron annuus s.l. (Asteraceae) in East Fennoscandia. Memo. Soc. Fauna Flora Fenn. 2019, 95, 40–59. [Google Scholar]
- Radford, I.J. Fluctuating resources, disturbance and plant strategies: Diverse mechanisms underlying plant invasions. J. Arid. Land 2013, 5, 284–297. [Google Scholar] [CrossRef]
- Flora of North America. Available online: http://www.efloras.org/florataxon.aspx?flora_id=1&taxon_id=200023888 (accessed on 21 July 2021).
- Pacanoski, Z. Current situation with invasive Erigeron annuus (l.) Pers. (daisy fleabane) in the Republic of Macedonia. EPPO Bull. 2017, 47, 118–124. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Site No. | Date | Location | Habitat | Altitude (MASL) |
|---|---|---|---|---|
| T1 | 5 June | 15.9702° E, 45.8017° N | Open, ruderal, by the road. Several dozens of individuals on 30-m long strip of soil and gravel. | 118 |
| T2 | 6 June | 15.9714° E, 45.8043° N | Semi-open, SE part of the botanical garden, near fence adjacent to the railway. Approximately 10–15 individuals. | 118 |
| T3 | 7 June | 15.9662° E, 45.8088° N | Shady, ruderal, adjacent to school playground and garbage containers. 5–10 individuals. | 120 |
| T4 | 11 June | 15.9764° E, 45.7716° N | Partial shade, adjacent to urban gardens with vegetables and surrounded by trees. 20–30 individuals. | 116 |
| T5 | 13 June | 15.9797° E, 45.7936° N | Partly open/closed, ruderal, next to bushes. | 111 |
| T6 | 17 June | 16.0354° E, 45.7657° N | Open, within grasslands on, at a time of sampling, dry flood channel of a levee. Present with locally dense stands with numerous individuals. | 107 |
| T7 | 18 June | 15.9637° E, 45.7788° N | Open, partly ruderal partly in secondary succession towards shrubs and trees. Few dozens of individuals scattered across an area of 200–300 m2. | 116 |
| T8 | 19 June | 15. 9829° E, 45.8644° N | Semi-open along the road and stream, ruderal. 20–30 individuals on an area of 100 m2. | 264 |
| T9 | 19 June | 16.0853° E, 45.9641° N | Open on a slope, surrounded by grasslands and other agricultural areas (on the other side of the road is forest). Very dense stand on 1000 m2. | 351 |
| T10 | 19 June | 15.9733° E, 45.9320° N | Semi-open and moist, by the road through forest. 10–20 individuals. | 569 |
| T11 | 19 June | 15.9544° E, 45.8994° N | Semi-open on a slope, by the road through forest. 5–10 individuals. | 908 |
| T12 | 19 June | 15.9420° E, 45.8843° N | Semi-open on a slope, by the road through forest. 10–20 individuals | 617 |
| T13 | 19 June | 15.9445° E, 45.8680° N | Open, by the road through forest. Several hundreds of individuals in dense stand on area adjacent to connection of main road with dirty road. | 443 |
| T14 | 4 September | 15.9427° E, 45.8700° N | Semi-open on a slope by the road. Several individuals. | 496 |
| T15 | 4 September | 15.9408° E, 45.8691° N | Semi-open, overgrown, beneath the remnants of a medieval fortress. Several individuals. | 554 |
| T16 | 4 September | 16.0072° E, 45.8325° N | Open, ruderal/grasslands, several dozen individuals scattered across an area of 200 m2. | 152 |
| T17 | 19 September | 16.0554° E, 45.8843° N | Semi-shaded on a slope, by the road through the settlement. Abandoned grassland. Several dozens of individuals scattered across an area of 500 m2. | 107 |
| T18 | 20 September | 16.0469° E, 45.8007° N | Semi-open, ruderal, adjacent to shopping mall. 10–20 individuals. | 211 |
| Life Trait | Mean | Median | Minimum | Maximum | Lower Quartile | Upper Quartile | Standard Error |
|---|---|---|---|---|---|---|---|
| Stem height (mm) | 1107.82 | 1090 | 600 | 1660 | 960 | 1270 | 10.89 |
| Fresh leaf mass (mg) | 211.08 | 207 | 38 | 552 | 129 | 265 | 4.59 |
| Dry leaf mass (mg) | 43.96 | 41 | 11 | 120 | 29 | 53 | 0.93 |
| Leaf area (mm2) | 1137.83 | 1105.40 | 231.54 | 2883.26 | 717.50 | 1453.25 | 24.04 |
| Leaf length (mm) | 79.48 | 79.74 | 10.28 | 129.25 | 64.73 | 91.66 | 0.89 |
| Leaf width (mm) | 22.79 | 22.23 | 10.30 | 47.61 | 16.19 | 28.15 | 0.36 |
| SLA (mm2 mg−1) | 26.35 | 25.51 | 15.39 | 46.24 | 22.02 | 30.66 | 0.29 |
| LDMC (mg g−1) | 215.07 | 209.30 | 135.22 | 440.00 | 191.67 | 228.81 | 2.04 |
| Site No. | Stem Height (mm) | Dry Matter Content (%) | Dry Leaf Mass (mg) | SLA (mm2 mg−1) | CSR Type |
|---|---|---|---|---|---|
| T1 | 1230 | 21 | 62 | 21 | CR |
| T2 | 1280 | 22 | 61 | 25 | CR |
| T3 | 1080 | 20 | 36 | 33 | CR |
| T4 | 1050 | 28 | 38 | 19 | C/CR |
| T5 | 1660 | 20 | 69 | 31 | CR |
| T6 | 1000 | 24 | 27 | 20 | CR |
| T7 | 1270 | 22 | 50 | 22 | CR |
| T8 | 1190 | 16 | 42 | 37 | CR |
| T9 | 1390 | 19 | 43 | 28 | CR |
| T10 | 1360 | 19 | 46 | 30 | CR |
| T11 | 1390 | 18 | 34 | 33 | CR |
| T12 | 1470 | 22 | 62 | 27 | CR |
| T13 | 1420 | 21 | 35 | 25 | CR |
| T14 | 1520 | 22 | 24 | 34 | CR |
| T15 | 1070 | 21 | 34 | 26 | CR |
| T16 | 1070 | 19 | 45 | 25 | CR |
| T17 | 1070 | 23 | 44 | 21 | CR |
| T18 | 1180 | 32 | 30 | 21 | C/CR |
| Effect | Test Name | Value | F | Degree of Connection | Connectivity Degree Error | p |
|---|---|---|---|---|---|---|
| Intercept | Wilks | 0.000052 | 1,127,896 | 7 | 411 | 0.00 |
| Locality | Wilks | 0.006365 | 26 | 119 | 2689.78064 | 0.00 |
| Life Trait | Altitude | Eastness | Northness | Slope | Mean Temp. in Summer | Mean Prec. in Summer | Mean Temp. in Spring | Mean Prec. in Spring | Total Precipitation | Mean Annual Temperature | Min. Annual Temperature |
|---|---|---|---|---|---|---|---|---|---|---|---|
| Stem height (mm) | 0.2274 | 0.2119 | 0.2120 | 0.2192 | −0.2334 | 0.2662 | 0.2595 | −0.2321 | 0.2671 | −0.2308 | 0.0754 |
| Fresh leaf mass (mg) | −0.0416 | 0.1005 | −0.1498 | −0.0908 | 0.0195 | 0.0007 | 0.0107 | 0.0206 | 0.0038 | 0.0221 | 0.3985 |
| Dry leaf mass (mg) | −0.2139 | 0.2108 | −0.2470 | −0.2201 | 0.2089 | −0.1670 | −0.1379 | 0.2073 | −0.1466 | 0.2117 | 0.4937 * |
| Leaf area (mm2) | 0.0511 | 0.1404 | −0.1757 | 0.0022 | −0.0800 | 0.1034 | 0.1109 | −0.0793 | 0.1079 | −0.0771 | 0.3482 |
| Leaf length (mm) | 0.1891 | 0.1824 | −0.0349 | 0.0430 | −0.1889 | 0.2251 | 0.2622 | −0.1937 | 0.2476 | −0.1859 | 0.2793 |
| Leaf width (mm) | 0.1162 | 0.1641 | −0.2132 | 0.1110 | −0.1503 | 0.1681 | 0.1642 | −0.1482 | 0.1666 | −0.1475 | 0.3022 |
| SLA (mm2 mg) | 0.4741 * | −0.0698 | 0.0948 | 0.4101 | −0.5228 * | 0.4932 * | 0.4608 | −0.5197 * | 0.4708 * | −0.5220 * | −0.1282 |
| LDMC (mg g−1) | −0.3855 | 0.1967 | −0.2053 | −0.3287 | 0.4278 | −0.3944 | −0.3514 | 0.4212 | −0.3614 | 0.4263 | −0.0160 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Levačić, D.; Jelaska, S.D. Consistent Grime’s CSR Strategy of Daisy Fleabane Erigeron annuus (L.) Pers. despite Its High Morphological Variability—A Case Study from Zagreb and Medvednica Mt., Croatia. Diversity 2022, 14, 45. https://doi.org/10.3390/d14010045
Levačić D, Jelaska SD. Consistent Grime’s CSR Strategy of Daisy Fleabane Erigeron annuus (L.) Pers. despite Its High Morphological Variability—A Case Study from Zagreb and Medvednica Mt., Croatia. Diversity. 2022; 14(1):45. https://doi.org/10.3390/d14010045
Chicago/Turabian StyleLevačić, Damjana, and Sven D. Jelaska. 2022. "Consistent Grime’s CSR Strategy of Daisy Fleabane Erigeron annuus (L.) Pers. despite Its High Morphological Variability—A Case Study from Zagreb and Medvednica Mt., Croatia" Diversity 14, no. 1: 45. https://doi.org/10.3390/d14010045