
Responses and Indicators of Composition, Diversity, and Productivity of Plant Communities at Different Levels of Disturbance in a Wetland Ecosystem

Abstract
:1. Introduction
2. Materials and Methods
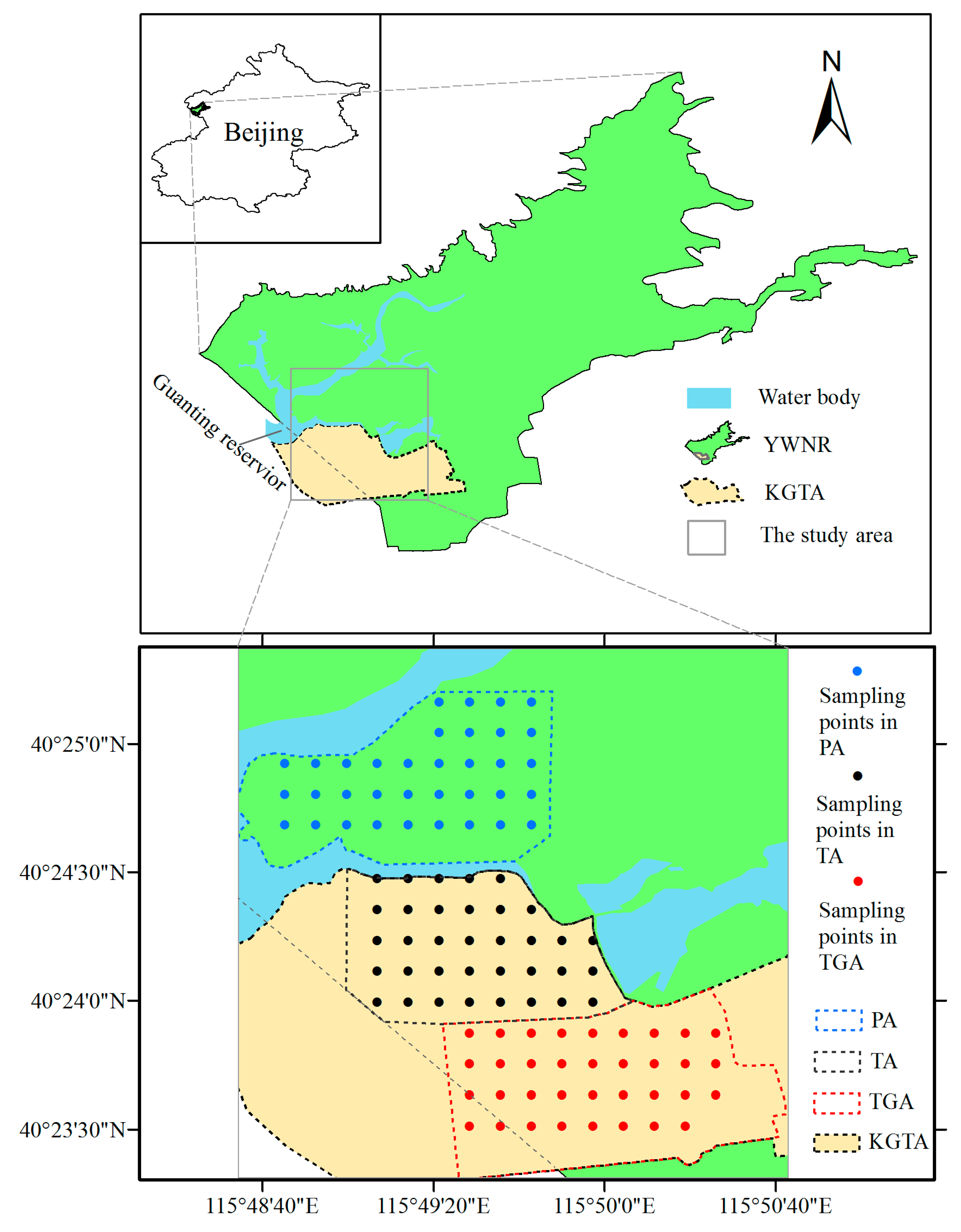
2.1. Study Area
2.2. Experimental Design and Sampling
2.3. Data Analysis
3. Results
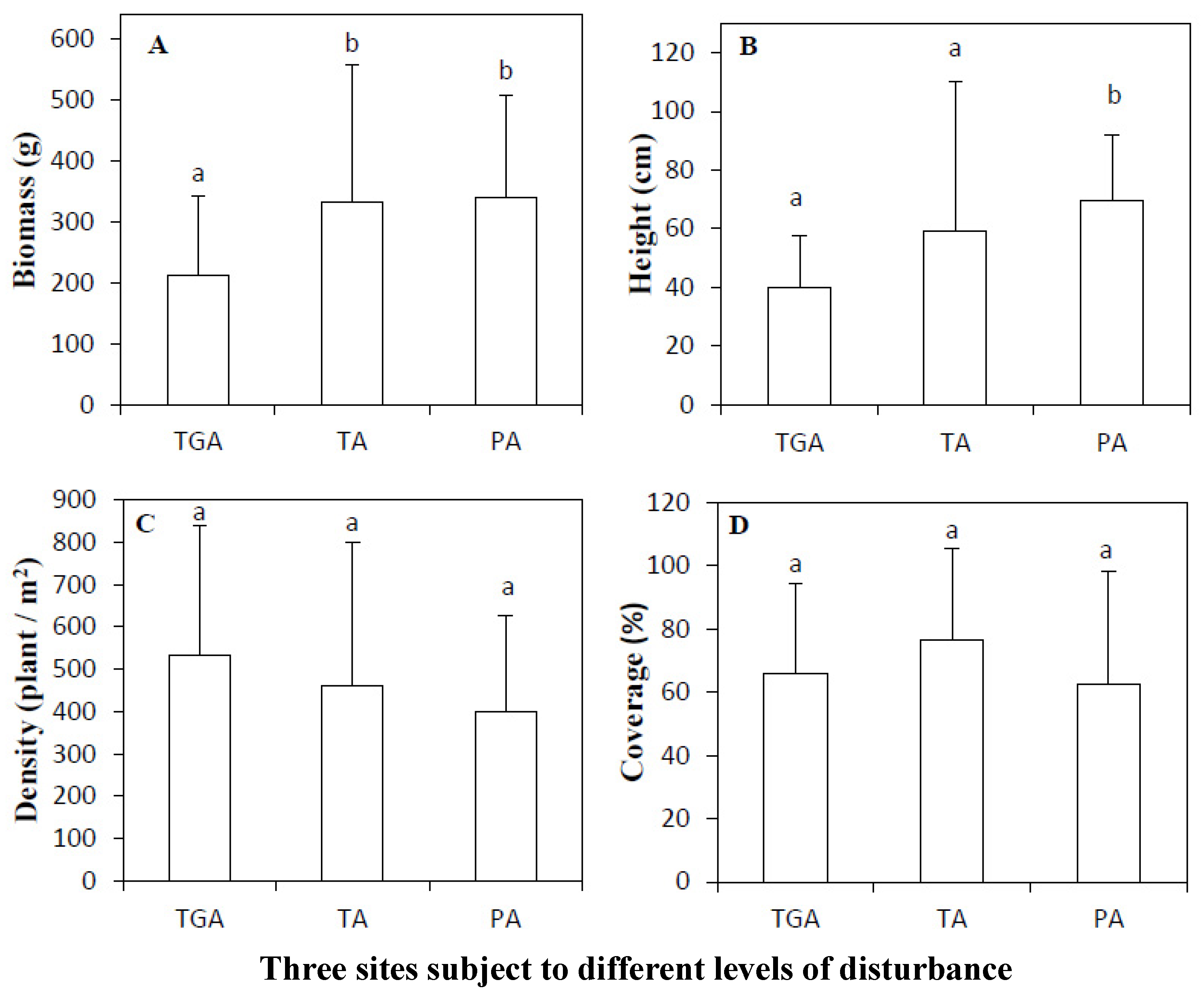
3.1. Comparison of Plant Growth in the Three Subareas
3.2. Composition of Plant Communities and Their Indicators in the Three Subareas
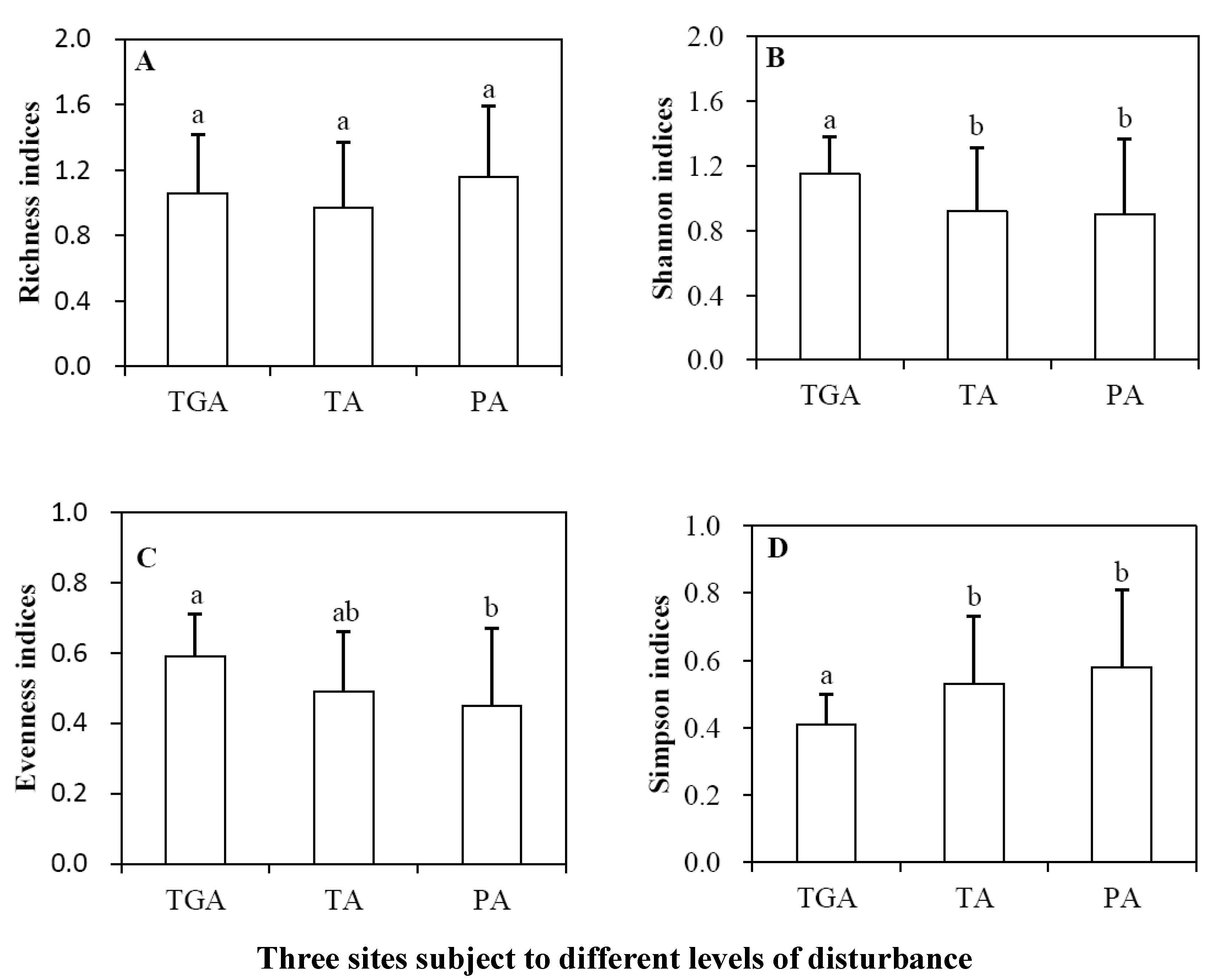
3.3. Plant Diversity of Three Subareas
3.4. Community Similarity among Three Subareas
3.5. Physical and Chemical Characteristics of the Soil of the Three Subareas
4. Discussion
4.1. The Rationality of the Division of Subareas
4.2. Responses and Indications of Plant Growth Parameters to Different Levels of Disturbance
4.3. Indicator Species in Plant Communities under Different Levels of Disturbance
4.4. Indicative Role of Diversity Indices under Different Levels of Disturbance
4.5. Indicators of Soil Characteristics under Different Levels of Disturbance
4.6. Management Applications
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zhong, L.-S.; Niu, Y.-F.; Liu, J.-M.; Chen, T. Development of Grassland Tourism Resource in Inner Mongolia Autonomous Region. J. Arid Land Resour. Environ. 2005, 19, 105–110. [Google Scholar]
- Shi, K.-B.; Wang, W.-R.; Yang, Y.-C.; Shao, R.; Shi, Y.-B. Impacts of tourists’activities on vegetation of Gannan grassland—A case study on tourist spot of Sangke prairie. Arid Zone Res. 2015, 32, 1220–1228. [Google Scholar]
- Mainguet, M. Desertification: Natural Background and Human Mismanagement; Speringer: Berlin/Heidelberg, Germany, 1994. [Google Scholar]
- Manzano, M.G.; Návar, J. Processes of desertification by goats overgrazing in the Tamaulipan thornscrub (matorral) in north-eastern Mexico. J. Arid Environ. 2000, 44, 1–17. [Google Scholar] [CrossRef]
- Su, Y.-Z.; Li, Y.-L.; Cui, J.-Y.; Zhao, W.-Z. Influences of continuous grazing and livestock exclusion on soil properties in a degraded sandy grassland, Inner Mongolia, northern China. Catena 2005, 59, 267–278. [Google Scholar] [CrossRef]
- Liu, L.-M.; Lv, J. The environmental influence of tourism on vegetation in typical grassland areas. Resour. Sci. 2009, 31, 442–449. [Google Scholar]
- Gao, X.-M.; Ma, K.-P.; Chen, L.-Z.; Li, D.-Q. The effects of tourism on species diversity of subalpine meadows in Dongling mountainous area, Beijing. Biodivers. Sci. 2002, 10, 189–195. [Google Scholar]
- Wu, J.-Z.; Shangguan, T.-L.; Zhang, J.; Li, J.-C.; Cao, T.-W.; Yue, J.-Y. Disturbing effects of tourism on plant species diversity of Malun subalpine meadow, Shanxi Province. J. MT Sci. 2007, 25, 534–540. [Google Scholar]
- Turner, C.L.; Seastedt, T.R.; Dyer, M.I. Maximization of aboveground grassland production—The role of defoliation frequency, intensity, and history. Ecol. Appl. 1993, 3, 175–186. [Google Scholar] [CrossRef]
- Virtanen, R. Effects of Grazing on Above-Ground Biomass on a Mountain Snowbed, NW Finland. Oikos 2000, 90, 295–300. [Google Scholar] [CrossRef]
- Patton, B.D.; Dong, X.J.; Nyren, P.E.; Nyren, A. Effects of Grazing Intensity, Precipitation, and Temperature on Forage Production. Rangel. Ecol. Manag. 2007, 60, 656–665. [Google Scholar] [CrossRef]
- de Mazancourt, C.; Loreau, M.; Abbadie, L. Grazing Optimization and Nutrient Cycling: When Do Herbivores Enhance Plant Production? Ecology 1998, 79, 2242–2252. [Google Scholar] [CrossRef]
- Semmartin, M.; Oesterheld, M. Effects of grazing pattern and nitrogen availability on primary productivity. Oecologia 2001, 126, 225–230. [Google Scholar] [CrossRef]
- Li, C.; Hao, X.; Zhao, M.; Han, G.; Willms, W.D. Influence of historic sheep grazing on vegetation and soil properties of a Desert Steppe in Inner Mongolia. Agric. Ecosyst. Environ. 2008, 128, 109–116. [Google Scholar] [CrossRef]
- Fedorov, N.I.; Zharkikh, T.L.; Mikhailenko, O.I.; Bakirova, R.T.; Martynenko, V.B. Forecast changes in the productivity of plant communities in the pre-urals steppe site of orenburg state nature reserve (russia) in extreme drought conditions using ndvi. Nat. Conserv. Res. 2019, 4 (Suppl. 2), 104–110. [Google Scholar] [CrossRef]
- Bulgakova, M.A.; Pyatina, E.V. The role of ungulates in soil zoocoenosis development of the steppe zone of the urals. Nat. Conserv. Res. 2019, 4 (Suppl. 2), 94–97. [Google Scholar] [CrossRef]
- Biondini, M.E.; Patton, B.D.; Nyren, P.E. Grazing Intensity and Ecosystem Processes in a Northern Mixed-Grass Prairie, USA. Ecol. Appl. 1998, 8, 469–479. [Google Scholar] [CrossRef]
- Hickman, K.R.; Hartnett, D.C.; Cochran, R.C.; Owensby, C.E. Grazing management effects on plant species diversity in tallgrass prairie. J. Range Manag. 2004, 57, 58–65. [Google Scholar] [CrossRef]
- Battisti, C.; Poeta, G.; Fanelli, G. An Introduction to Disturbance Ecology; Springer International Publishing: Cham, Switzerland, 2016. [Google Scholar]
- Waser, N.M.; Price, M.V. Effects of Grazing on Diversity of Annual Plants in the Sonoran Desert. Oecologia 1981, 50, 407–411. [Google Scholar] [CrossRef]
- Pykälä, J. Cattle Grazing Increases Plant Species Richness of Most Species Trait Groups in Mesic Semi-Natural Grasslands. Plant Ecol. 2004, 175, 217–226. [Google Scholar] [CrossRef]
- Marty, J.T. Effects of Cattle Grazing on Diversity in Ephemeral Wetlands. Conserv. Biol. 2005, 19, 1626–1632. [Google Scholar] [CrossRef]
- Schultz, N.L.; Morgan, J.W.; Lunt, I.D. Effects of grazing exclusion on plant species richness and phytomass accumulation vary across a regional productivity gradient. J. Veg. Sci. 2011, 22, 130–142. [Google Scholar] [CrossRef]
- Shi, X.-M.; Li, X.G.; Li, C.T.; Zhao, Y.; Shang, Z.H.; Ma, Q. Grazing exclusion decreases soil organic C storage at an alpine grassland of the Qinghai–Tibetan Plateau. Ecol. Eng. 2013, 57, 183–187. [Google Scholar] [CrossRef]
- Sasaki, T.; Okubo, S.; Okayasu, T.; Jamsran, U.; Ohkuko, T.; Takeuchi, K. Management Applicability of the Intermediate Disturbance Hypothesis across Mongolian Rangeland Ecosystems. Ecol. Appl. 2009, 19, 423–432. [Google Scholar] [CrossRef] [Green Version]
- Fensham, R.J.; Silcock, J.L.; Dwyer, J.M. Plant species richness responses to grazing protection and degradation history in a low productivity landscape. J. Veg. Sci. 2011, 22, 997–1008. [Google Scholar] [CrossRef]
- Deng, L.; Sweeney, S.; Shangguan, Z.P. Grassland responses to grazing disturbance: Plant diversity changes with grazing intensity in a desert steppe. Grass Forage Sci. 2014, 69, 524–533. [Google Scholar] [CrossRef]
- Ganjurjav, H.; Duan, M.-J.; Wan, Y.-F.; Zhang, W.-N.; Gao, Q.-Z.; Li, Y.; Jiangcun, W.-Z.; Danjiu, L.-B.; Guo, H.-B. Effects of grazing by large herbivores on plant diversity and productivity of semi-arid alpine steppe on the Qinghai-Tibetan Plateau. Rangel. J. 2015, 37. [Google Scholar] [CrossRef]
- Proulx, M.; Mazumder, A. reversal of grazing impact on plant species richness innutrient-poor vs. nutrient-rich ecosystems. Ecology 1998, 79, 2581–2592. [Google Scholar] [CrossRef]
- Osem, Y.; Perevolotsky, A.; Kigel, J. Grazing Effect on Diversity of Annual Plant Communities in a Semi-Arid Rangeland: Interactions with Small-Scale Spatial and Temporal Variation in Primary Productivity. J. Ecol. 2002, 90, 936–946. [Google Scholar] [CrossRef]
- Shrestha, G.; Stahl, P.D. Carbon accumulation and storage in semi-arid sagebrush steppe: Effects of long-term grazing exclusion. Agric. Ecosyst. Environ. 2008, 125, 173–181. [Google Scholar] [CrossRef]
- Austrheim, G.; Eriksson, O. Plant Species Diversity and Grazing in the Scandinavian Mountains: Patterns and Processes at Different Spatial Scales. Ecography 2001, 24, 683–695. [Google Scholar] [CrossRef]
- Fernández-Lugo, S.; de Nascimento, L.; Mellado, M.; Arévalo, J.R. Grazing effects on species richness depends on scale: A 5-year study in tenerife pastures (canary islands). Plant Ecol. 2011, 212, 423–432. [Google Scholar] [CrossRef]
- Jing, Z.; Cheng, J.; Su, J.; Bai, Y.; Jin, J. Changes in plant community composition and soil properties under 3-decade grazing exclusion in semiarid grassland. Ecol. Eng. 2014, 64, 171–178. [Google Scholar] [CrossRef]
- O’Connor, T.G.; Martindale, G.; Morris, C.D.; Short, A.; Witkowski, E.T.F.; Scott-Shaw, R. Influence of Grazing Management on Plant Diversity of Highland Sourveld Grassland, KwaZulu-Natal, South Africa. Rangel. Ecol. Manag. 2011, 64, 196–207. [Google Scholar] [CrossRef]
- Fenu, G.; Cogoni, D.; Ulian, T.; Bacchetta, G. The impact of human trampling on a threatened coastal Mediterranean plant: The case of Anchusa littorea Moris (Boraginaceae). Flora Morphol. Distrib. Funct. Ecol. Plants 2013, 208, 104–110. [Google Scholar] [CrossRef]
- Santoro, R.; Jucker, T.; Prisco, I.; Carboni, M.; Battisti, C.; Acosta, A.T.R. Effects of Trampling Limitation on Coastal Dune Plant Communities. Environ. Manag. 2012, 49, 534–542. [Google Scholar] [CrossRef]
- Du, G.-S. The study of trophic condition of Kwangting reservoir. J. Teach. Coll. (Nat. Sci. E) 1989, 10, 56–61. [Google Scholar]
- Du, G.S.; Wang, J.T.; Zhang, W.H.; Feng, L.Q.; Liu, J. On the nutrient status of guanting reservoir, 2001–2002. J. Lake Sci. 2004, 16, 277–281. [Google Scholar]
- Zhao-Ning, G.; Gong, H.-L.; Zhao, W.-J. The Ecological Evolution of Beijing Wetland: A Case Study in Beijing Wild Duck Lake; Beijing Science &Technology Press: Beijing, China, 2007. [Google Scholar]
- Wang, Z.; Gong, H.; Zhang, J. Receding water line and interspecific competition determines plant community composition and diversity in wetlands in Beijing. PLoS ONE 2015, 10, e0124156. [Google Scholar] [CrossRef]
- Zhao, W.J.; Wang, Y.H.; Gong, Z.N.; Hu, D.Y. 3S Integrated Practice Instruction-Taking Yeyahu Wetland as an Example; China Environmental Science Press: Beijing, China, 2011. [Google Scholar]
- Wang, Y.; Gong, H.L.; Zhao, W.J.; Li, X.J.; Zhang, Z.F.; Zhao, W. The change of Yeyahu wetland resources in Beijing. Acta Geogr. Sin. 2005, 60, 656–664. [Google Scholar]
- Beijing Municipal Bureau of Statistics. Statistical Bulletin of the National Economic and Social Development of Yanqing County, Beijing. 2004. Available online: http://www.tjcn.org/tjgb/01bj/247.html (accessed on 5 October 2020).
- Bureau of Statistics of Yanqing County, Beijing. Statistical Bulletin of the National Economic and Social Development of Yanqing County, Beijing. 2011. Available online: http://www.tjcn.org/tjgb/01bj/25878_2.html (accessed on 5 October 2020).
- Xie, X.M. Soil and Plant Nutrition Experiment; Zhejiang University Press: Hangzhou, China, 2014. [Google Scholar]
- Bao, S.D. Soil and Agricultural Chemistry Analysis; China Agricultural Press: Beijing, China, 2000. [Google Scholar]
- Clifford, H.T.; Stephenson, W. An Introduction to Numerical Classification; Academic Press: London, UK, 1975. [Google Scholar]
- Shannon, C.E.; Weaver, W. The Mathematical Theory of Communication; University of Illinois Press: Urbana, IL, USA, 1949. [Google Scholar]
- Magurran, A.E. Measuring Biological Diversity; Blackwell Publishing Company: Malden, MA, USA, 2004. [Google Scholar]
- Pielou, E.C. An Introduction to Mathematical Ecology; Wiley InterScience: New York, NY, USA, 1969. [Google Scholar]
- Pielou, E.C. Ecological Diversity; Wiley InterScience: New York, NY, USA, 1975. [Google Scholar]
- Simpson, E.H. Measurement of diversity. Nature 1949, 163, 688. [Google Scholar] [CrossRef]
- Magurran, A.E. Ecological Diversity and Its Measurement; Princeton University Press: Princeton, NJ, USA, 1988. [Google Scholar]
- Krebs, C.J. Ecological Methodology; Benjamin/Cummings: Menlo Park, CA, USA, 1999. [Google Scholar]
- Wang, B. Phytocoenology; Higher Education Press: Beijing, China, 1987. [Google Scholar]
- Chen, W.; Hu, D.; Fu, B.Q. Research on Biodiversity in Beijing Wetland; Science Press: Beijing, China, 2007. [Google Scholar]
- Gong, Z.N.; Gong, H.L.; Hu, D. Wetlnd Plants in Wild Duke Lake, Beijing; China Environmental Science Press: Beijing, China, 2012. [Google Scholar]
- Walters, C.M.; Martin, M.C. An Examination of the Effects of Grazing on Vegetative and Soil Parameters in the tallgrass praire. Trans. Kans. Acad. Sci. 2003, 106, 59–70. [Google Scholar] [CrossRef]
- Pei, S.; Fu, H.; Wan, C. Changes in soil properties and vegetation following exclosure and grazing in degraded Alxa desert steppe of Inner Mongolia, China. Agric. Ecosyst. Environ. 2008, 124, 33–39. [Google Scholar] [CrossRef]
- Wu, G.L.; Du, G.Z.; Liu, Z.H.; Thirgood, S. Effect of fencing and grazing on a kobresia-dominated meadow in the qinghai-tibetan plateau. Plant Soil 2009, 319, 115–126. [Google Scholar] [CrossRef]
- Gao, J.J.; Carmel, Y. Can the intermediate disturbance hypothesis explain grazing–diversity relations at a global scale? Oikos 2020, 129, 493–502. [Google Scholar] [CrossRef]
- Li, J.; Loneragan, W.A.; Duggin, J.A.; Grant, C.D. Issues affecting the measurement of disturbance response patterns in herbaceous vegetation—A test of the intermediate disturbance hypothesis. Plant Ecol. 2004, 172, 11–26. [Google Scholar] [CrossRef]
- Pueyoa, Y.; Aladosa, C.L.; Ferrer-Benimelib, C. Is the analysis of plant community structure better than common species-diversity indices for assessing the effects of livestock grazing on a Mediterranean arid ecosystem? J. Arid Environ. 2006, 64, 698–712. [Google Scholar] [CrossRef]
- Battisti, C.; Contoli, L. Diversity Indices as ‘Magic’ Tools inLandscape Planning: A Cautionary Note on their Uncritical Use. Landsc. Res. 2011, 36, 111–117. [Google Scholar] [CrossRef]
- Weber, K.T.; Gokhale, B.S. Effect of grazing on soil-water content in semiarid rangelands of southeast Idaho. J. Arid Environ. 2011, 75, 464–470. [Google Scholar] [CrossRef]
- Willms, W.D.; McGinn, S.M.; Dormaar, J.F. Influence of litter on herbage production in the mixed prairie. J. Range Manag. 1993, 46, 320–324. [Google Scholar] [CrossRef] [Green Version]
- Snyman, H.A. Root studies on grass species in a semi-arid South Africa along a degradation gradient. Agric. Ecosyst. Environ. 2009, 130, 100–108. [Google Scholar] [CrossRef]
- Battisti, C.; Amori, G.; Luiselli, L. Toward a new generation of effective problem solvers and project-oriented applied ecologists. Web Ecol. 2020, 20, 11–17. [Google Scholar] [CrossRef]
- Wu, G.-L.; Liu, Z.-H.; Zhang, L.; Chen, J.-M.; Hu, T.-M. Long-term fencing improved soil properties and soil organic carbon storage in an alpine swamp meadow of western China. Plant Soil 2010, 332, 331–337. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Site | N | Mesophytic Species | Hygrophytic Species | ||
|---|---|---|---|---|---|
| nm (Percentage) | IVm (%) | nh (Percentage) | IVh (%) | ||
| TGA | 36 | 28 (77.8%) | 53 | 8 (22.2%) | 47 |
| TA | 49 | 37 (75.5%) | 49.7 | 12 (24.5%) | 50.3 |
| PA | 43 | 31 (72.1%) | 35.7 | 12 (27.9%) | 64.3 |
| Site | nm (Percentage) | IVm (%) (Percentage) | ||
|---|---|---|---|---|
| Dominant Species | Rare Species | Dominant Species | Rare Species | |
| TGA | 6 (67%) | 30 (80%) | 77.5 (45%) | 22.5 (80%) |
| TA | 6 (50%) | 43 (79%) | 64.6 (36.8%) | 35.4 (75%) |
| PA | 5 (40%) | 38 (76%) | 62.6 (15.8%) | 37.4 (69%) |
| TGA | TA | PA |
|---|---|---|
| Salsola collina Ixeris chinensis Elymus dahuricus Artemisia scoparia Lespedeza davurica Astragalus adsurgens | Calamagrostis epigejos * | Cynanchum chinensis Inula japonica * Sonchus brachyotus * Artemisia japonica Kummerowia stipulacea |
| Species Groups | PA and TGA | PA and TA | TGA and TA |
|---|---|---|---|
| Dominant species | 0.38 | 0.38 | 0.71 |
| Rare species | 0.31 | 0.37 | 0.46 |
| All species | 0.41 | 0.48 | 0.55 |
| Site | Water Content (%) | Organic Matter Content (g/kg) | pH |
|---|---|---|---|
| TGA | 4.85 ± 1.69 a | 13.97 ± 4.32 a | 8.79 ± 0.22 a |
| TA | 10.58 ± 11.56 b | 10.67 ± 7.47 b | 8.59 ± 0.18 b |
| PA | 13.58 ± 8.72 c | 14.22 ± 7.01 a | 8.57 ± 0.29 b |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Duan, T.; Zhang, J.; Wang, Z. Responses and Indicators of Composition, Diversity, and Productivity of Plant Communities at Different Levels of Disturbance in a Wetland Ecosystem. Diversity 2021, 13, 252. https://doi.org/10.3390/d13060252
Duan T, Zhang J, Wang Z. Responses and Indicators of Composition, Diversity, and Productivity of Plant Communities at Different Levels of Disturbance in a Wetland Ecosystem. Diversity. 2021; 13(6):252. https://doi.org/10.3390/d13060252
Chicago/Turabian StyleDuan, Tingting, Jing Zhang, and Zhengjun Wang. 2021. "Responses and Indicators of Composition, Diversity, and Productivity of Plant Communities at Different Levels of Disturbance in a Wetland Ecosystem" Diversity 13, no. 6: 252. https://doi.org/10.3390/d13060252