
Insects in the City: Does Remnant Native Habitat Influence Insect Order Distributions?

 , , , , and
, , , , and
Abstract
:1. Introduction
2. Materials and Methods
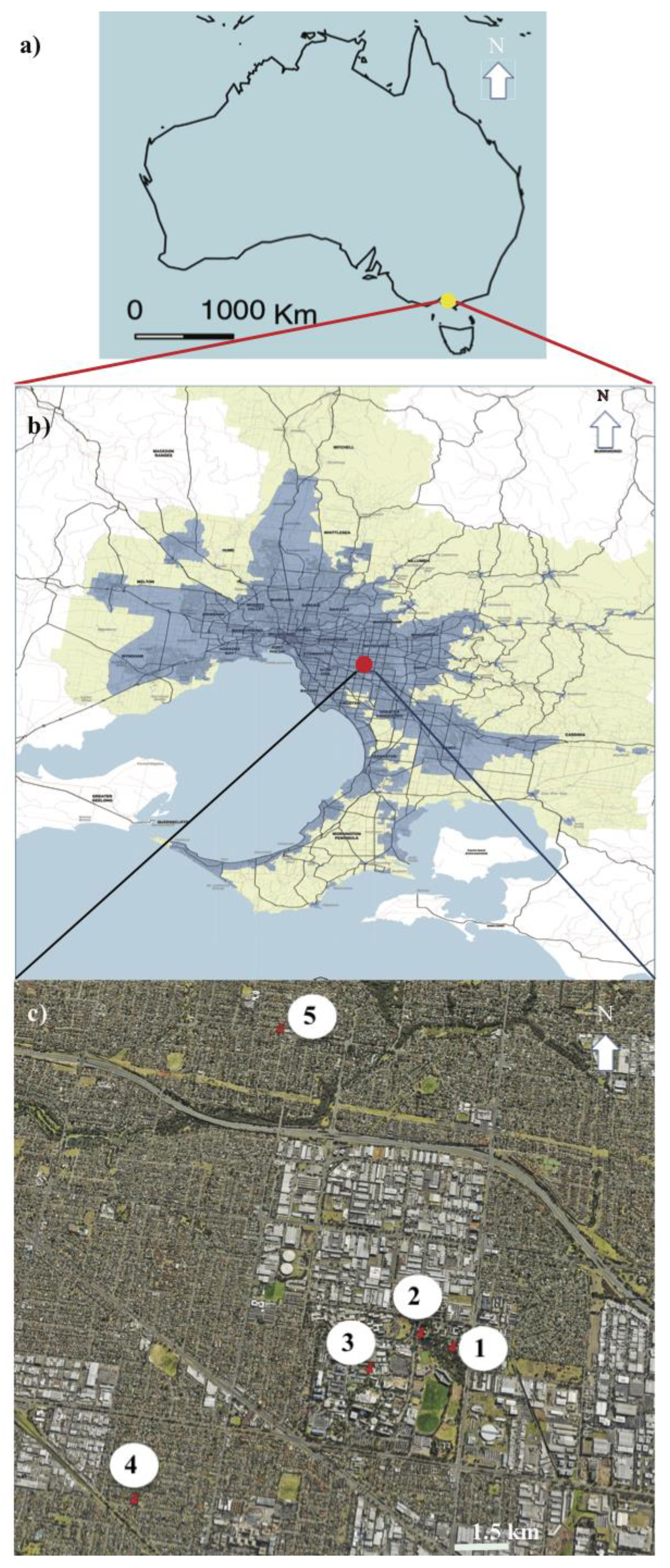
2.1. Study Area
2.2. Data Collection
2.3. Insect Identification
2.4. Data Analyses
3. Results
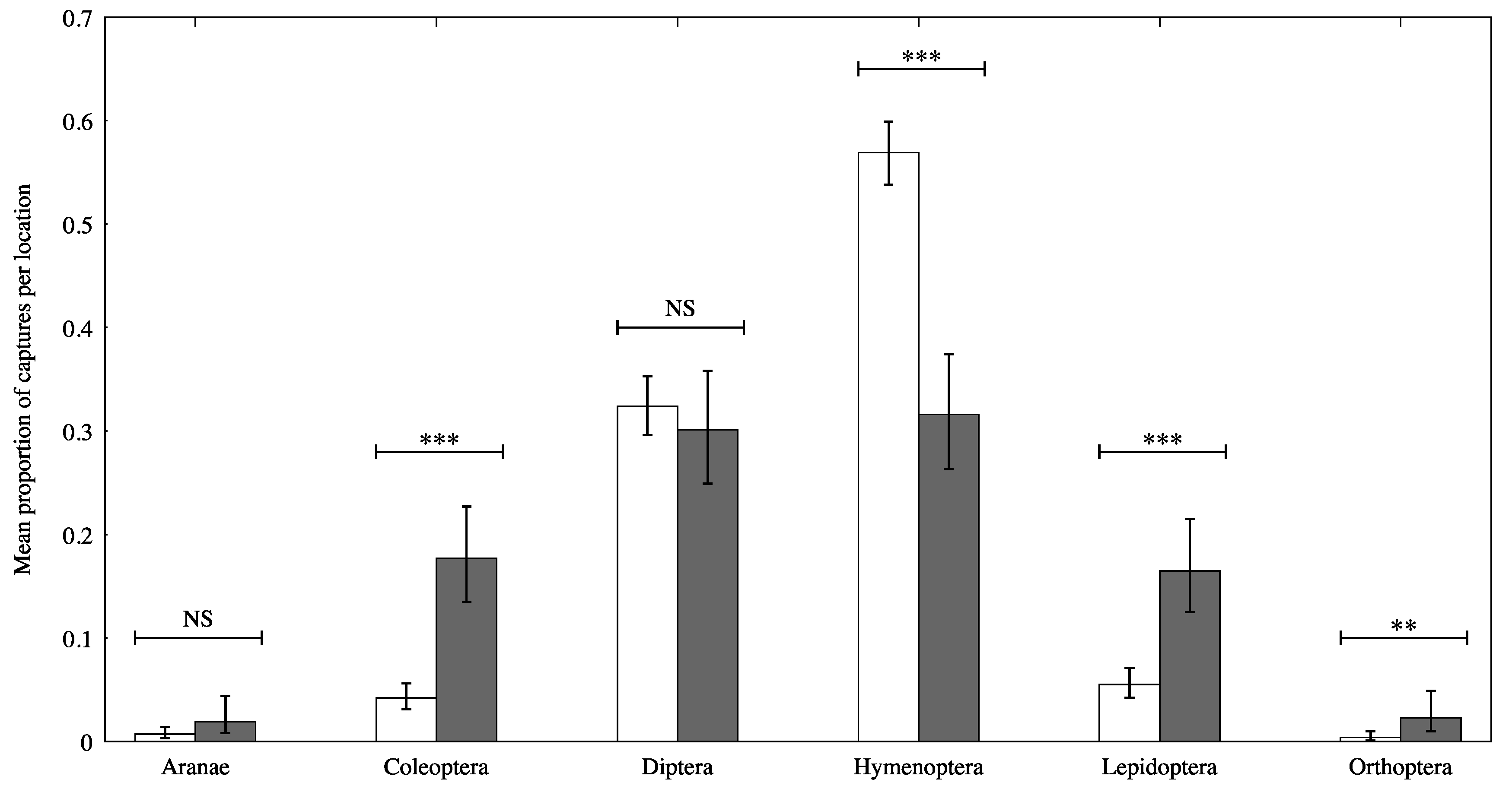
3.1. Data Overview
3.2. Model Parameters
3.3. Pairwise Post Hoc Comparisons
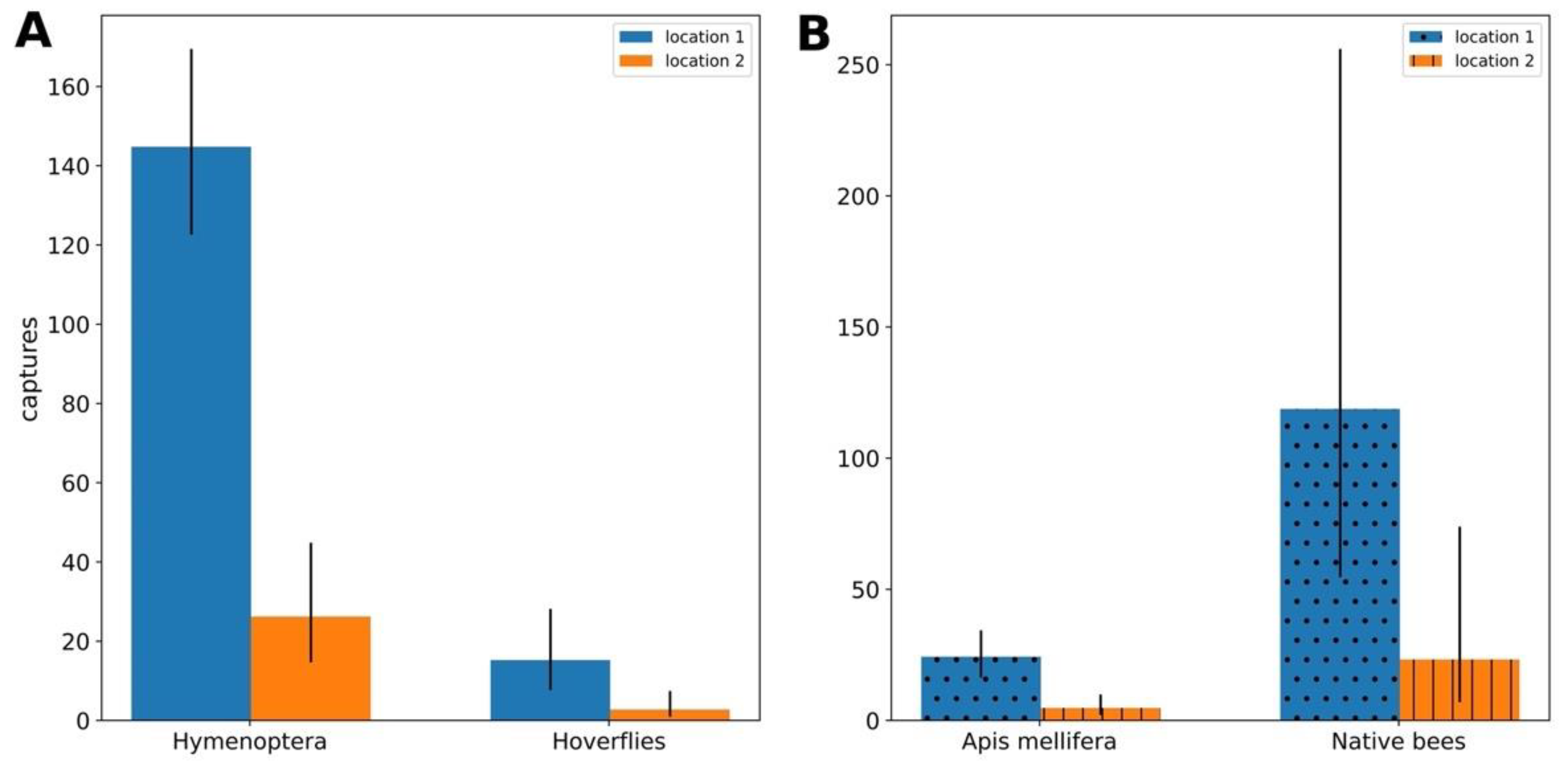
3.4. Hymenopteran Bee vs. Syrphidae Capture Comparison
3.5. Apis Mellifera vs. Native Bee Captures Comparison
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- United Nations. World Urbanization Prospects: The 2014 Revision, Highlights; United Nations, Department of Economic and Social Affairs, Population Division: New York, NY, USA, 2014. [Google Scholar]
- Elmqvist, T.; Michail, F.; Goodness, J.; Güneralp, B.; Marcotullio, P.J.; McDonald, R.I.; Parnell, S.; Schewenius, M.; Sendstad, M.; Seto, K.C.; et al. Urbanization, Biodiversity and Ecosystem Services: Challenges and Opportunities; Springer Nature: Basingstoke, UK, 2013; Volume 103, ISBN 0169-2046. [Google Scholar]
- Dearborn, D.C.; KARK, S. Motivations for Conserving Urban Biodiversity. Conserv. Biol. 2010, 24, 432–440. [Google Scholar] [CrossRef] [PubMed]
- Ives, C.D.; Lentini, P.E.; Threlfall, C.G.; Ikin, K.; Shanahan, D.F.; Garrard, G.E.; Bekessy, S.A.; Fuller, R.A.; Mumaw, L.; Rayner, L.; et al. Cities are hotspots for threatened species. Glob. Ecol. Biogeogr. 2016, 25, 117–126. [Google Scholar] [CrossRef]
- New, T.R. Promoting and developing insect conservation in Australia’s urban environments. Austral Entomol. 2018, 57, 182–193. [Google Scholar] [CrossRef]
- Cook, W.M.; Faeth, S.H. Irrigation and Land Use Drive Ground Arthropod Community Patterns in an Urban Desert. Environ. Entomol. 2006, 35, 1532–1540. [Google Scholar] [CrossRef]
- Hunter, M.R.; Hunter, M.D. Designing for conservation of insects in the built environment. Insect Conserv. Divers. 2008, 22, 189–196. [Google Scholar] [CrossRef] [Green Version]
- Ikin, K.; Le Roux, D.S.; Rayner, L.; Villaseñor, N.R.; Eyles, K.; Gibbons, P.; Manning, A.D.; Lindenmayer, D.B. Key lessons for achieving biodiversity-sensitive cities and towns. Ecol. Manag. Restor. 2015, 16, 206–214. [Google Scholar] [CrossRef]
- Garrard, G.E.; Williams, N.S.G.; Mata, L.; Thomas, J.; Bekessy, S.A. Biodiversity Sensitive Urban Design. Conserv. Lett. 2018, 11, e12411. [Google Scholar] [CrossRef]
- Sánchez-Bayo, F.; Wyckhuys, K.A.G. Worldwide decline of the entomofauna: A review of its drivers. Biol. Conserv. 2019, 232, 8–27. [Google Scholar] [CrossRef]
- Lindenmayer, D.B.; Fischer, J. Habitat Fragmentation and Landscape Change: An Ecological and Conservation Synthesis; Island Press: Washington, DC, USA, 2013; ISBN 159726606X. [Google Scholar]
- Battisti, C. Habitat fragmentation, fauna and ecological network planning: Toward a theoretical conceptual framework. Ital. J. Zool. 2003, 70, 241–247. [Google Scholar] [CrossRef]
- Salafsky, N.; Salzer, D.; Stattersfield, A.J.; Hilton-Taylor, C.; Neugarten, R.; Butchart, S.H.M.; Collen, B.; Cox, N.; Master, L.L.; O’Connor, S.; et al. A standard lexicon for biodiversity conservation: Unified classifications of threats and actions. Conserv. Biol. 2008, 22, 897–911. [Google Scholar] [CrossRef]
- Levin, M.D. Value of Bee Pollination to U.S. Agriculture. Bull. Entomol. Soc. Am. 1983, 29, 50–51. [Google Scholar] [CrossRef]
- Cook, D.F.; Voss, S.C.; Finch, J.T.D.; Rader, R.C.; Cook, J.M.; Spurr, C.J. The role of flies as pollinators of horticultural crops: An Australian case study with worldwide relevance. Insects 2020, 11, 341. [Google Scholar] [CrossRef]
- Shrestha, M.; Burd, M.; Garcia, J.E.; Dorin, A.; Dyer, A.G. Colour evolution within orchids depends on whether the pollinator is a bee or a fly. Plant Biol. 2019, 21, 745–752. [Google Scholar] [CrossRef]
- Hall, D.M.; Camilo, G.R.; Tonietto, R.K.; Ollerton, J.; Ahrné, K.; Arduser, M.; Ascher, J.S.; Baldock, K.C.R.; Fowler, R.; Frankie, G.; et al. The city as a refuge for insect pollinators. Conserv. Biol. 2017, 31, 24–29. [Google Scholar] [CrossRef] [PubMed]
- New, T.R. Insect Conservation and Urban Environments; Springer: Berlin/Heidelberg, Germany, 2015; ISBN 3319212230. [Google Scholar]
- Hostetler, N.E.; McIntyre, M.E.; McIntyre, N.E.; Hostetler, M.E. Effects of urban land use on pollinator (Hymenoptera: Apoidea) communities in a desert metropolis. Basic Appl. Ecol. 2001, 2, 209. [Google Scholar] [CrossRef] [Green Version]
- Goulson, D.; Lye, G.C.; Darvill, B. Decline and conservation of bumble bees. Annu. Rev. Entomol. 2008, 53, 191–208. [Google Scholar] [CrossRef]
- Matteson, K.C.; Ascher, J.S.; Langellotto, G.A. Bee Richness and Abundance in New York City Urban Gardens. Ann. Entomol. Soc. Am. 2008, 101, 140–150. [Google Scholar] [CrossRef]
- Winfree, R.; Aguilar, R.; Vázquez, D.P.; LeBuhn, G.; Aizen, M.A. A meta-analysis of bees’ responses to anthropogenic disturbance. Ecology 2009, 90, 2068–2076. [Google Scholar] [CrossRef]
- Leong, M.; Kremen, C.; Roderick, G.K. Pollinator Interactions with Yellow Starthistle (Centaurea solstitialis) across Urban, Agricultural, and Natural Landscapes. PLoS ONE 2014, 9, e086357. [Google Scholar]
- Baldock, K.C.R.; Goddard, M.A.; Hicks, D.M.; Kunin, W.E.; Mitschunas, N.; Osgathorpe, L.M.; Potts, S.G.; Robertson, K.M.; Scott, A.V.; Stone, G.N.; et al. Where is the UK’s pollinator biodiversity? The importance of urban areas for flower-visiting insects. Proc. R. Soc. Lond. B Biol. Sci. 2015, 282, 20142849. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Turrini, T.; Knop, E. A landscape ecology approach identifies important drivers of urban biodiversity. Glob. Chang. Biol. 2015, 21, 1652–1667. [Google Scholar] [CrossRef]
- Ziter, C. The biodiversity–ecosystem service relationship in urban areas: A quantitative review. Oikos 2016, 125, 761–768. [Google Scholar] [CrossRef]
- Ahrné, K.; Bengtsson, J.; Elmqvist, T. Bumble Bees (Bombus spp) along a Gradient of Increasing Urbanization. PLoS ONE 2009, 4, e05574. [Google Scholar] [CrossRef] [Green Version]
- Bates, A.J.; Sadler, J.P.; Fairbrass, A.J.; Falk, S.J.; Hale, J.D.; Matthews, T.J. Changing bee and hoverfly pollinator assemblages along an urban-rural gradient. PLoS ONE 2011, 6, e023459. [Google Scholar] [CrossRef] [PubMed]
- Yuen, S.W.; Bonebrake, T.C. Artificial night light alters nocturnal prey interception outcomes for morphologically variable spiders. PeerJ 2017, 5, e4070. [Google Scholar] [CrossRef] [PubMed]
- Czaczkes, T.J.; Bastidas-Urrutia, A.M.; Ghislandi, P.; Tuni, C. Reduced light avoidance in spiders from populations in light-polluted urban environments. Sci. Nat. 2018, 105, 64. [Google Scholar] [CrossRef] [PubMed]
- Harrison, T.; Winfree, R. Urban drivers of plant-pollinator interactions. Funct. Ecol. 2015, 29, 879–888. [Google Scholar] [CrossRef]
- Sands, D.P.A. Important issues facing insect conservation in Australia: Now and into the future. Austral Entomol. 2018, 57, 150–172. [Google Scholar] [CrossRef] [Green Version]
- McLoughlin, S. The breakup history of Gondwana and its impact on pre-Cenozoic floristic provincialism. Aust. J. Bot. 2001, 49, 271–300. [Google Scholar] [CrossRef]
- Exon, N.F.; Hill, P.J.; Lafoy, Y.; Heine, C.; Bernardel, G. Kenn Plateau off northeast Australia: A continental fragment in the southwest Pacific jigsaw. Aust. J. Earth Sci. 2006, 53, 541–564. [Google Scholar] [CrossRef]
- Armstrong, J.A. Biotic pollination mechanisms in the Australian flora—A review. N. Z. J. Bot. 1979, 17, 467–508. [Google Scholar] [CrossRef]
- Heard, T.A. The Australian Native Bee Book: Keeping Stingless Bee Hives for Pets, Pollination, and Sugarbag Honey; Sugarbag Bees: Brisbane, Australia, 2016. [Google Scholar]
- Gilpin, A.-M.; Denham, A.J.; Ayre, D.J. Are there magnet plants in Australian ecosystems: Pollinator visits to neighbouring plants are not affected by proximity to mass flowering plants. Basic Appl. Ecol. 2019, 35, 34–44. [Google Scholar] [CrossRef]
- Shrestha, M.; Dyer, A.G.; Bhattarai, P.; Burd, M. Flower colour and phylogeny along an altitudinal gradient in the Himalayas of Nepal. J. Ecol. 2014, 102, 126–135. [Google Scholar] [CrossRef]
- Dyer, A.G.; Boyd-Gerny, S.; McLoughlin, S.; Rosa, M.G.P.; Simonov, V.; Wong, B.B.M. Parallel evolution of angiosperm colour signals: Common evolutionary pressures linked to hymenopteran vision. Proc. R. Soc. B 2012, 279, 3606–3615. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Austin, A.D.; Yeates, D.K.; Cassis, G.; Fletcher, M.J.; Salle, J.L.; Lawrence, J.F.; McQuillan, P.B.; Mound, L.A.; Bickel, D.J.; Gullan, P.J.; et al. Insects ‘Down Under’—Diversity, endemism and evolution of the Australian insect fauna: Examples from select orders. Aust. J. Entomol. 2004, 43, 216–234. [Google Scholar] [CrossRef]
- Hahs, A.K.; McDonnell, M.J.; McCarthy, M.A.; Vesk, P.A.; Corlett, R.T.; Norton, B.A.; Clemants, S.E.; Duncan, R.P.; Thompson, K.; Schwartz, M.W.; et al. A global synthesis of plant extinction rates in urban areas. Ecol. Lett. 2009, 12, 1165–1173. [Google Scholar] [CrossRef]
- Victorian Planning Authority Announcement. Available online: https://vpa.vic.gov.au/ (accessed on 28 May 2019).
- Australian Bureau of Statistics, Australian Government. Available online: https://www.abs.gov.au/ (accessed on 28 May 2019).
- The Economist Intelligence Unit Limited. Available online: https://www.eiu.com/ (accessed on 28 May 2019).
- R Core Team R Core Team. R: A Language and Environment for Statistical Computing; Version 3.5.1.; R Foundation for Statistical Computing: Vienna, Austria, 2018; Available online: https://www.R-project.org/ (accessed on 28 May 2019).
- Heithaus, E.R. Community Structure of Neotropical Flower Visiting Bees and Wasps: Diversity and Phenology. Ecology 1979, 60, 190–202. [Google Scholar] [CrossRef]
- Peakall, R.; Beattie, A.J. Pollination of the Orchid Microtis parviflora R. Br. by Flightless Worker Ants. Funct. Ecol. 1989, 3, 515–522. [Google Scholar] [CrossRef]
- Shuttleworth, A.; Johnson, S. Specialized Pollination by Large Spider-Hunting Wasps and Self-Incompatibility in the African Milkweed Pachycarpus asperifolius. Int. J. Plant Sci. 2006, 167, 1177–1186. [Google Scholar] [CrossRef]
- Ollerton, J.; Liede-Schumann, S.; Endress, M.E.; Meve, U.; Rech, A.R.; Shuttleworth, A.; Keller, H.A.; Fishbein, M.; Alvarado-Cárdenas, L.O.; Amorim, F.W.; et al. The diversity and evolution of pollination systems in large plant clades: Apocynaceae as a case study. Ann. Bot. 2019, 123, 311–325. [Google Scholar] [CrossRef] [Green Version]
- Bawa, K.S.; Bullock, S.H.; Perry, D.R.; Coville, R.E.; Grayum, M.H. Reproductive biology of tropical lowland rain forest trees. II. Pollination systems. Am. J. Bot. 1985, 72, 346–356. [Google Scholar] [CrossRef]
- Paudel, B.R.; Shrestha, M.; Burd, M.; Adhikari, S.; Sun, Y.S.; Li, Q.J. Coevolutionary elaboration of pollination-related traits in an alpine ginger (Roscoea purpurea) and a tabanid fly in the Nepalese Himalayas. New Phytol. 2016, 211, 1402–1411. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paudel, B.R.; Shrestha, M.; Dyer, A.G.; Zhu, X.F.; Abdusalam, A.; Li, Q.J. Out of Africa: Evidence of the obligate mutualism between long corolla tubed plant and long-tongued fly in the Himalayas. Ecol. Evol. 2015, 5, 5240–5251. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lunau, K. Visual ecology of flies with particular reference to colour vision and colour preferences. J. Comp. Physiol. A 2014, 200, 497–512. [Google Scholar] [CrossRef] [PubMed]
- Shrestha, M.; Lunau, K.; Dorin, A.; Schulze, B.; Bischoff, M.; Burd, M.; Dyer, A.G. Floral colours in a world without birds and bees: The plants of Macquarie Island. Plant Biol. 2016, 18, 842–850. [Google Scholar] [CrossRef]
- Doyle, T.; Hawkes, W.L.S.; Massy, R.; Powney, G.D.; Menz, M.H.M.; Wotton, K.R. Pollination by hoverflies in the Anthropocene: Pollination by Hoverflies. Proc. R. Soc. B Biol. Sci. 2020, 287. [Google Scholar] [CrossRef]
- Orford, K.A.; Vaughan, I.P.; Memmott, J. The forgotten flies: The importance of non-syrphid Diptera as pollinators. Proc. R. Soc. B Biol. Sci. 2015, 282, doi. [Google Scholar] [CrossRef] [Green Version]
- Pauw, A.; Stofberg, J.; Waterman, R.J. Flies and Flowers in Darwin’s Race. Evolution 2009, 63, 268–279. [Google Scholar] [CrossRef]
- Gottsberger, G. Flowers and Beetles in the South American Tropics. Bot. Acta 1990, 103, 360–365. [Google Scholar] [CrossRef]
- Bernhardt, P. Convergent evolution and adaptive radiation of beetle-pollinated angiosperms. Plant Syst. Evol. 2000, 222, 293–320. [Google Scholar] [CrossRef]
- Paudel, B.R.; Shrestha, M.; Dyer, A.G.; Li, Q. Ginger and the beetle: Evidence of primitive pollination system in a Himalayan endemic alpine ginger (Roscoea alpina, Zingiberaceae). PLoS ONE 2017, 12, e0180460. [Google Scholar] [CrossRef] [Green Version]
- Micheneau, C.; Fournel, J.; Warren, B.H.; Hugel, S.; Gauvin-Bialecki, A.; Pailler, T.; Strasberg, D.; Chase, M.W. Orthoptera, a new order of pollinator. Ann. Bot. 2010, 105, 355–364. [Google Scholar] [CrossRef]
- Heiling, A.M.; Herberstein, M.E.; Chittka, L. Pollinator attraction: Crab-spiders manipulate flower signals. Nature 2003, 421, 334. [Google Scholar] [CrossRef] [PubMed]
- Huey, S.; Nieh, J.C. Foraging at a safe distance: Crab spider effects on pollinators. Ecol. Entomol. 2017, 42, 469–476. [Google Scholar] [CrossRef] [Green Version]
- Jackson, R.R.; Pollard, S.D.; Nelson, X.J.; Edwards, G.B.; Barrion, A.T. Jumping spiders (Araneae: Salticidae) that feed on nectar. J. Zool. 2001, 255, 25–29. [Google Scholar] [CrossRef]
- Shrestha, M.; Tscheulin, T.; Garcia, J.E.; Chua, J.H.J.; Howard, S.R.; Dorin, A.; Nielsen, A.; Dyer, A.G. Fluorescent pan traps affect the capture rate of insect orders in different ways. Insects 2019, 10, 40. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nielsen, A.; Steffan-Dewenter, I.; Westphal, C.; Messinger, O.; Potts, S.G.; Roberts, S.P.M.; Settele, J.; Szentgyörgyi, H.; Vaissière, B.E.; Vaitis, M. Assessing bee species richness in two Mediterranean communities: Importance of habitat type and sampling techniques. Ecol. Res. 2011, 26, 969–983. [Google Scholar] [CrossRef]
- LeBuhn, G.; Droege, S.; Connor, E.; Gemmill-Herren, B.; Azzu, N. Protocol to Detect and Monitor Pollinator Communities: Guidance for Practitioners; Food and Agriculture Organization: Quebec City, QC, Canada, 2016. [Google Scholar]
- Michener, C.D. The Bees of the World, 2nd ed.; Johns Hopkins: Baltimore, MD, USA, 2007. [Google Scholar]
- Zborowski, P.; Storey, R. Field Guide to Insects in Australia; New Holand Publisher, CSIRO: Clayton, Australia, 2010; ISBN 0730104141. [Google Scholar]
- Dollin, A.; Batley, M.; Robinson, M.; Faulkner, B. Native Bees of the Sydney Region, A Field Guide; Australian Native Bee Research Centre: North Richmond, Australia, 2000. [Google Scholar]
- Gullan, P.J.; Cranston, P.S. The Insects: An Outline of Entomology; John Wiley & Sons: Hoboken, NJ, USA, 2009; ISBN 1405144572. [Google Scholar]
- The Core Team, R. R: A Language and Environment for Statistical Computing; V 4.02.; R Foundation for Statistical Computing: Vienna, Austria, 2020; Available online: https://www.r-project.org/ (accessed on 28 May 2019).
- Fox, J.; Weisberg, S. An R Companion to Applied Regression, 3rd ed.; Sage: Thousand Oaks, CA, USA, 2019. [Google Scholar]
- Lenth, R. Emmeans: Estimated Marginal Means, Aka Least-Squares Means. R Package Version 1.5-2-1. Available online: https://cran.r-project.org/package=emmeans (accessed on 29 October 2020).
- Faraway, J.J. Extending the Linear Model with R; Carlin, B.P., Chatfield, C., Tanner, M., Zidek, J., Eds.; Chapman & Hall/CRC: Boca Raton, FL, USA, 2006. [Google Scholar]
- Zuur, A.F.; Hilbe, J.M.; Ieno, E.N. A Beginner’s Guide to GLM and GLMM with R: A Frequentist and Bayesian Perspective for Ecologists; Highland Statistics Limited: New Burgh, NY, USA, 2013; ISBN 0957174144. [Google Scholar]
- Zuur Ieno, E.N.; Walker, N.; Saveliev, A.A.; Smith, G.M.A.; Zuur, I.E.N.; Walker, N.; Saveliev, A.A.; Smith, G.M.A. Mixed Effects Models and Extensions in Ecology with R; Springer: New York, NY, USA, 2009. [Google Scholar]
- Hardin, J.W.; Hilbe, J.M.; Hilbe, J. Generalized Linear Models and Extensions; Second Stata Press: College Station, TX, USA, 2007. [Google Scholar]
- Nagelkerke, N.J.D. A note on a general definition of the coefficient of determination miscellanea a note on a general definition of the coefficient of determination. Biometrika 1991, 78, 691–692. [Google Scholar] [CrossRef]
- Searle, S.R.; Speed, F.M.; Milliken, G.A. Population Marginal Means in the Linear Model: An Alternative to Least Squares Means. Am. Stat. 1980, 34, 216–221. [Google Scholar]
- Briscoe, A.D.; Chittka, L. The evolution of color vision in insects. Annu. Rev. Entomol. 2001, 46, 471–510. [Google Scholar] [CrossRef] [Green Version]
- Giurfa, M.; Núñez, J.; Chittka, L.; Menzel, R. Colour preferences of flower-naive honeybees. J. Comp. Physiol. A 1995, 177, 247–259. [Google Scholar] [CrossRef]
- Dyer, A.G.; Boyd-Gerny, S.; Shrestha, M.; Lunau, K.; Garcia, J.E.; Koethe, S.; Wong, B.B.M.M. Innate colour preferences of the Australian native stingless bee Tetragonula carbonaria Sm. J. Comp. Physiol. A 2016, 202, 603–613. [Google Scholar] [CrossRef] [PubMed]
- Discovering and Documenting Australia’s Native Bees. Available online: https://www.taxonomyaustralia.org.au/discoverbees (accessed on 28 May 2019).
- Gautam, G.; Paudel, A.; Poudel, L.; Shrestha, M. Limno-plankton diversity and water quality in shallow Lake Ecosystem in central Nepal. Int. J. Adv. Res. Biol. Sci 2016, 3, 131–139. [Google Scholar]
- Burkett, D.A.; Butler, J.F.; Kline, D.L. Field evaluation of colored light-emitting diodes as attractants for woodland mosquitoes and other Diptera in north central Florida. J. Am. Mosq. Control Assoc. News 1998, 14, 186–195. [Google Scholar]
- Cohnstaedt, L.W.; Gillen, J.I.; Munstermann, L.E. Light-emitting diode technology improves insect trapping. J. Am. Mosq. Control Assoc. 2008, 24, 331–334. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Horváth, G.; Kriska, G.; Malik, P.; Robertson, B. Polarized light pollution: A new kind of ecological photopollution. Front. Ecol. Environ. 2009, 7, 317–325. [Google Scholar] [CrossRef] [Green Version]
- Souza de Medeiros, B.A.; Barghini, A.; Souza de Medeiros, B.A. UV radiation as an attractor for insects. Leukos 2012, 9, 47–56. [Google Scholar]
- Kaluza, B.F.; Wallace, H.; Heard, T.A.; Klein, A.-M.; Leonhardt, S.D. Urban gardens promote bee foraging over natural habitats and plantations. Ecol. Evol. 2016, 6, 1304–1316. [Google Scholar] [CrossRef] [Green Version]
- Palma, E.; Catford, J.A.; Corlett, R.T.; Duncan, R.P.; Hahs, A.K.; McCarthy, M.A.; McDonnell, M.J.; Thompson, K.; Williams, N.S.G.; Vesk, P.A. Functional trait changes in the floras of 11 cities across the globe in response to urbanization. Ecography 2016, 40, 875–886. [Google Scholar] [CrossRef]
- Barth, F.G. Insects and Flowers. The Biology of a Partnership; Princeton University Press USA: Princeton, NJ, USA, 1985; ISBN 0045740291. [Google Scholar]
- Chittka, L.; Thomson, J.D.; Waser, N.M. Flower constancy, insect psychology, and plant evolution. Naturwissenschaften 1999, 86, 361–377. [Google Scholar] [CrossRef]
- Cruz-Cárdenas, J.; Oleas, N.H. Private Urban Garden Satisfaction and Its Determinants in Quito, Ecuador. SAGE Open 2018, 8. [Google Scholar] [CrossRef] [Green Version]
- Fortel, L.; Henry, M.; Guilbaud, L.; Guirao, A.L.; Kuhlmann, M.; Mouret, H.; Rollin, O.; Vaissière, B.E. Decreasing abundance, increasing diversity and changing structure of the wild bee community (Hymenoptera: Anthophila) along an urbanization gradient. PLoS ONE 2014, 9, e0104679. [Google Scholar] [CrossRef] [PubMed]
- Sirohi, M.H.; Jackson, J.; Edwards, M.; Ollerton, J. Diversity and abundance of solitary and primitively eusocial bees in an urban centre: A case study from Northampton (England). J. Insect Conserv. 2015, 19, 487–500. [Google Scholar] [CrossRef] [Green Version]
- Hannah, L.; Dyer, A.G.; Garcia, J.E.; Dorin, A.; Burd, M. Psychophysics of the hoverfly: Categorical or continuous color discrimination? Curr. Zool. 2019, 65, 483–492. [Google Scholar] [CrossRef]
- van der Kooi, C.J.; Dyer, A.G.; Kevan, P.G.; Lunau, K. Functional significance of the optical properties of flowers for visual signalling. Ann. Bot. 2018, 123, 263–276. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pardee, G.L.; Philpott, S.M. Native plants are the bee’s knees: Local and landscape predictors of bee richness and abundance in backyard gardens. Urban Ecosyst. 2014, 17, 641–659. [Google Scholar] [CrossRef] [Green Version]
- Fukase, J.; Simons, A.M. Increased pollinator activity in urban gardens with more native flora. Appl. Ecol. Environ. Res. 2016, 14, 297–310. [Google Scholar] [CrossRef]
- Bennett, A.F. Habitat linkages: A key element in an integrated landscape approach to conservation. Parks 1997, 7, 43–49. [Google Scholar]
- Goulson, D.; Hughes, W.; Derwent, L.; Stout, J. Colony growth of the bumblebee, Bombus terrestris, in improved and conventional agricultural and suburban habitats. Oecologia 2002, 130, 267–273. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Order | Mean Proportion Difference (95% CI) | Odds Ratio | z | p |
|---|---|---|---|---|
| Araneae | −1.02 (−2.18, 0.133) | 0.360 | −1.73 | 0.083 |
| Coleoptera | −1.59 (−2.03, −1.15) | 0.224 | −7.09 | <0.001 |
| Diptera | 0.106 (−0.186, 0.398) | 1.11 | 0.711 | 0.477 |
| Hymenoptera | 1.05 (0.759, 1.34) | 2.86 | 7.18 | <0.001 |
| Lepidopterans | −1.23 (−1.65, −0.809) | 0.215 | −5.73 | <0.001 |
| Orthopterans | −1.77 (−3.04, −0.499) | 0.170 | −2.73 | 0.006 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shrestha, M.; Garcia, J.E.; Thomas, F.; Howard, S.R.; Chua, J.H.J.; Tscheulin, T.; Dorin, A.; Nielsen, A.; Dyer, A.G. Insects in the City: Does Remnant Native Habitat Influence Insect Order Distributions? Diversity 2021, 13, 148. https://doi.org/10.3390/d13040148
Shrestha M, Garcia JE, Thomas F, Howard SR, Chua JHJ, Tscheulin T, Dorin A, Nielsen A, Dyer AG. Insects in the City: Does Remnant Native Habitat Influence Insect Order Distributions? Diversity. 2021; 13(4):148. https://doi.org/10.3390/d13040148
Chicago/Turabian StyleShrestha, Mani, Jair E. Garcia, Freya Thomas, Scarlett R. Howard, Justin H. J. Chua, Thomas Tscheulin, Alan Dorin, Anders Nielsen, and Adrian G. Dyer. 2021. "Insects in the City: Does Remnant Native Habitat Influence Insect Order Distributions?" Diversity 13, no. 4: 148. https://doi.org/10.3390/d13040148