
Taxonomy and Phylogeny Reveal Two New Potential Edible Ectomycorrhizal Mushrooms of Thelephora from East Asia

 ,
,  and
and
Abstract
:1. Introduction
2. Materials and Methods
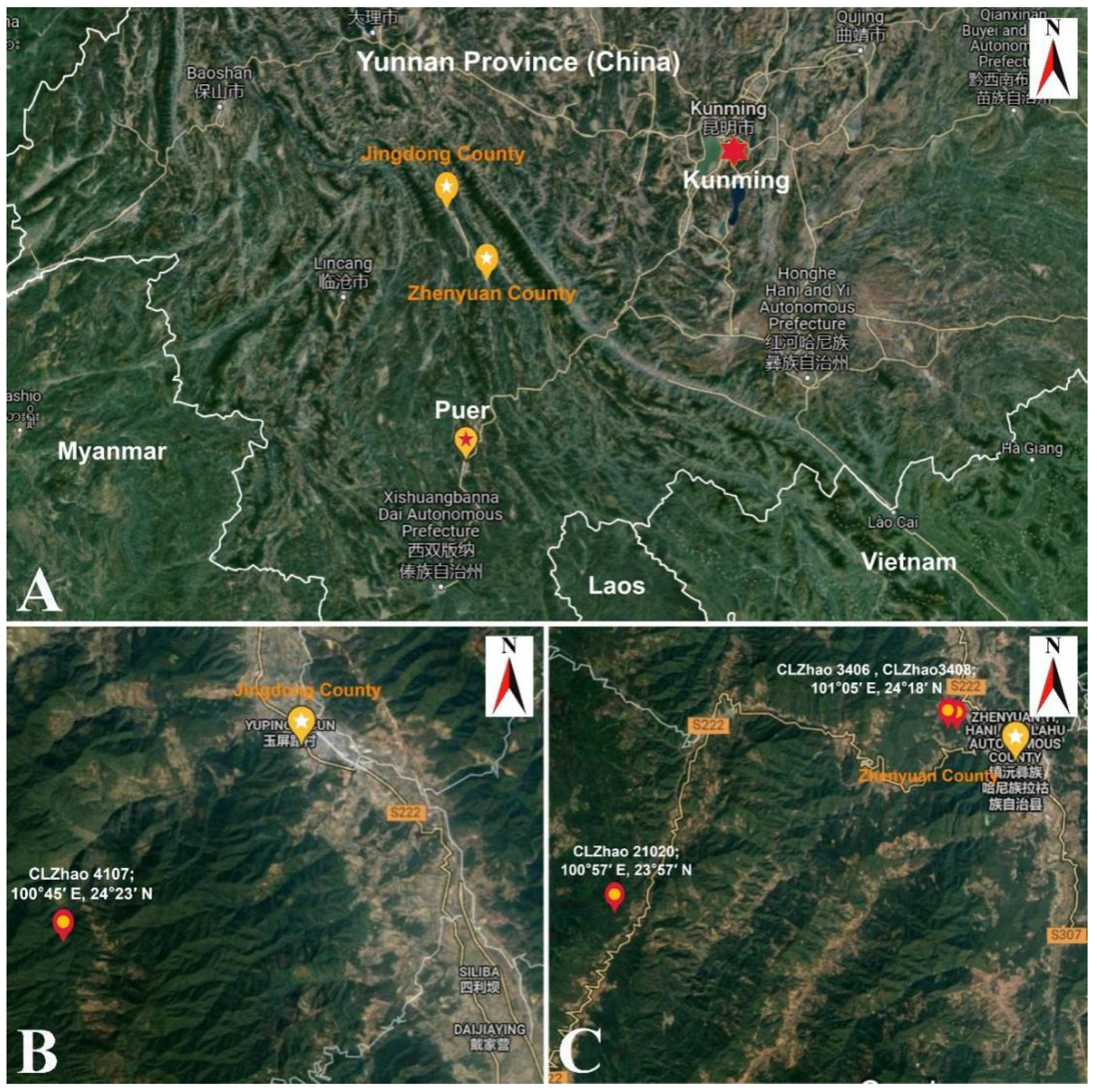
2.1. Specimens Collection and Herbarium Specimen Preparation
2.2. Morphology
2.3. Molecular Phylogeny
2.4. Pairwise Homoplasy Test
3. Results
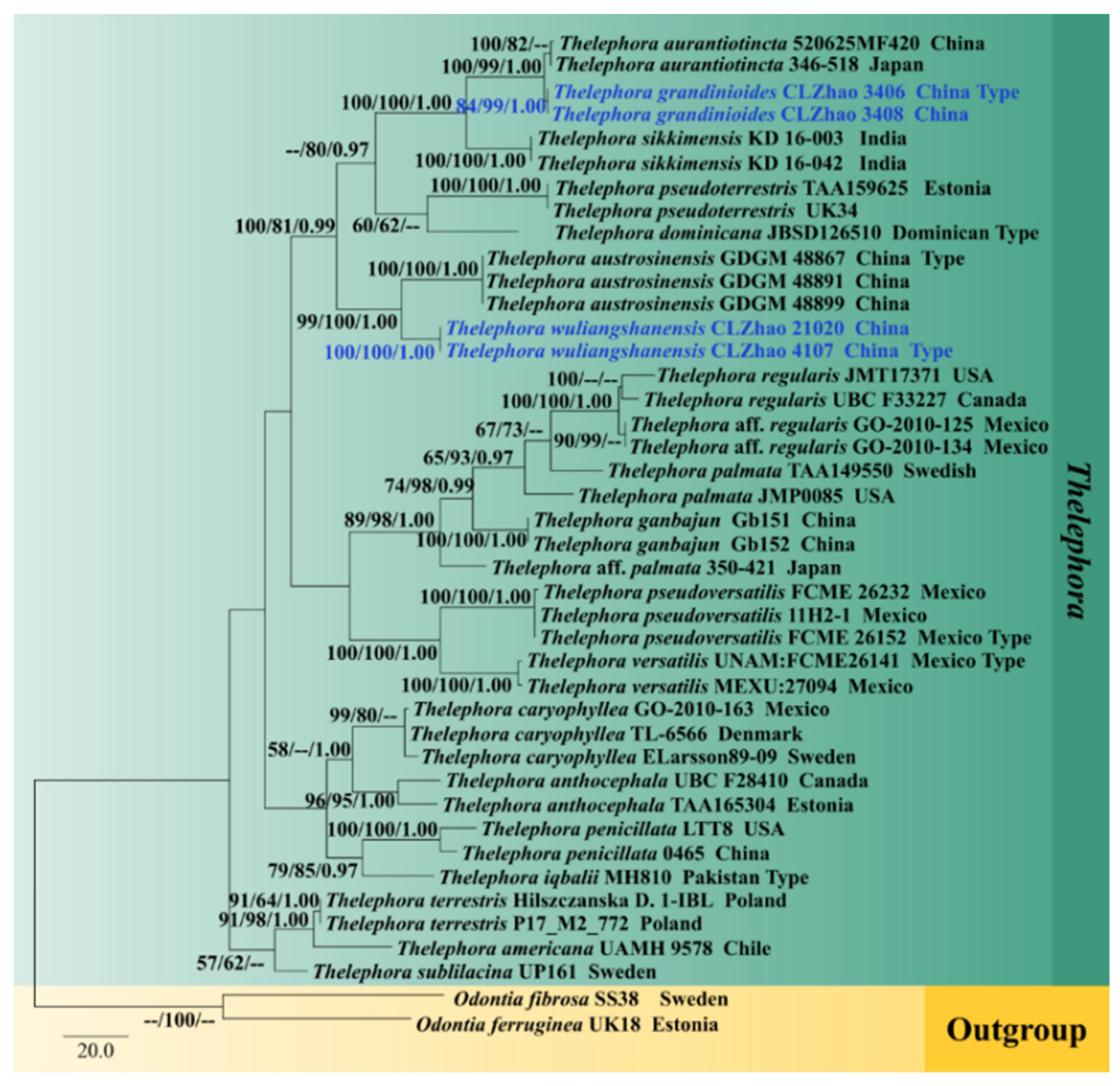
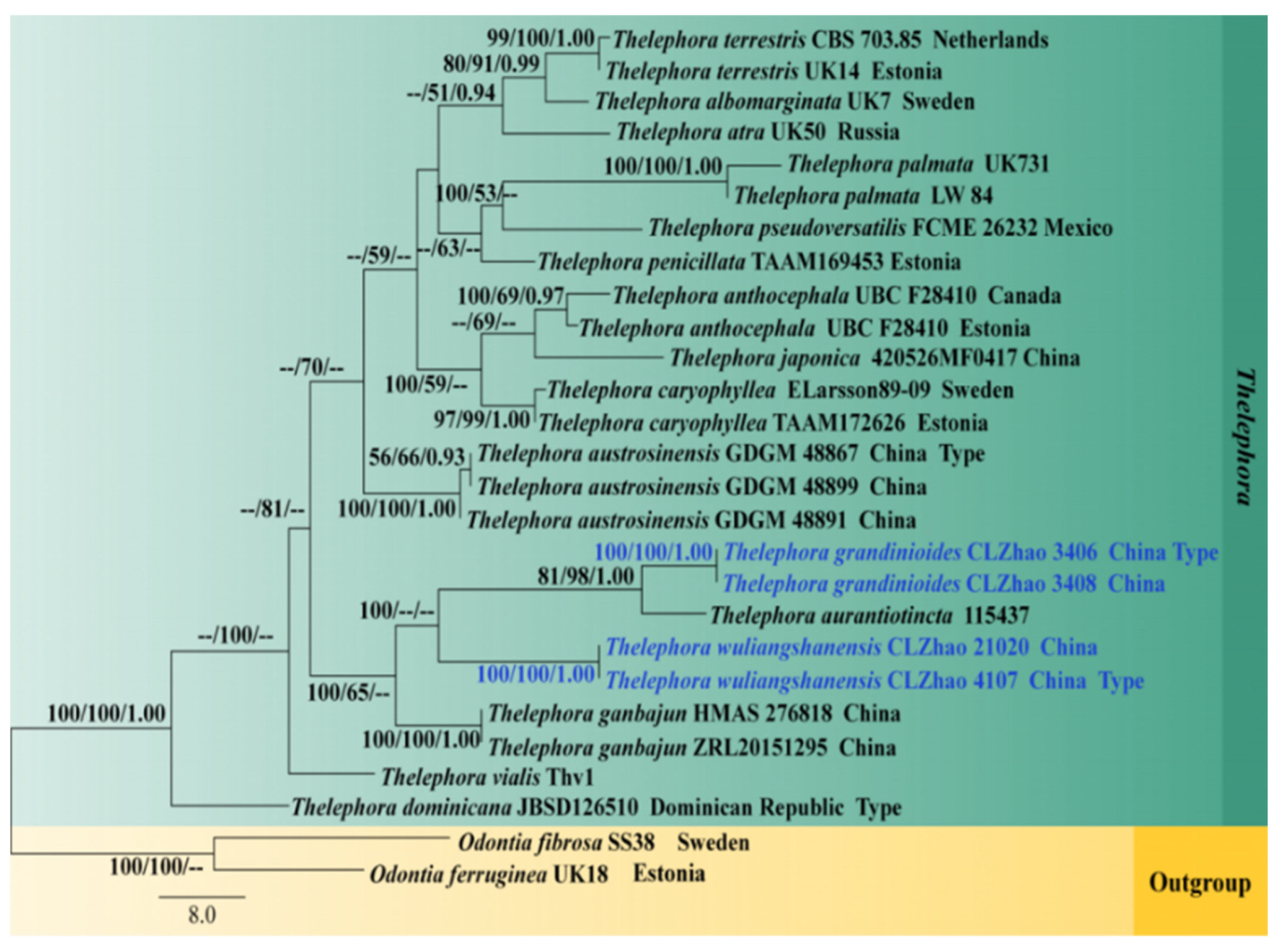
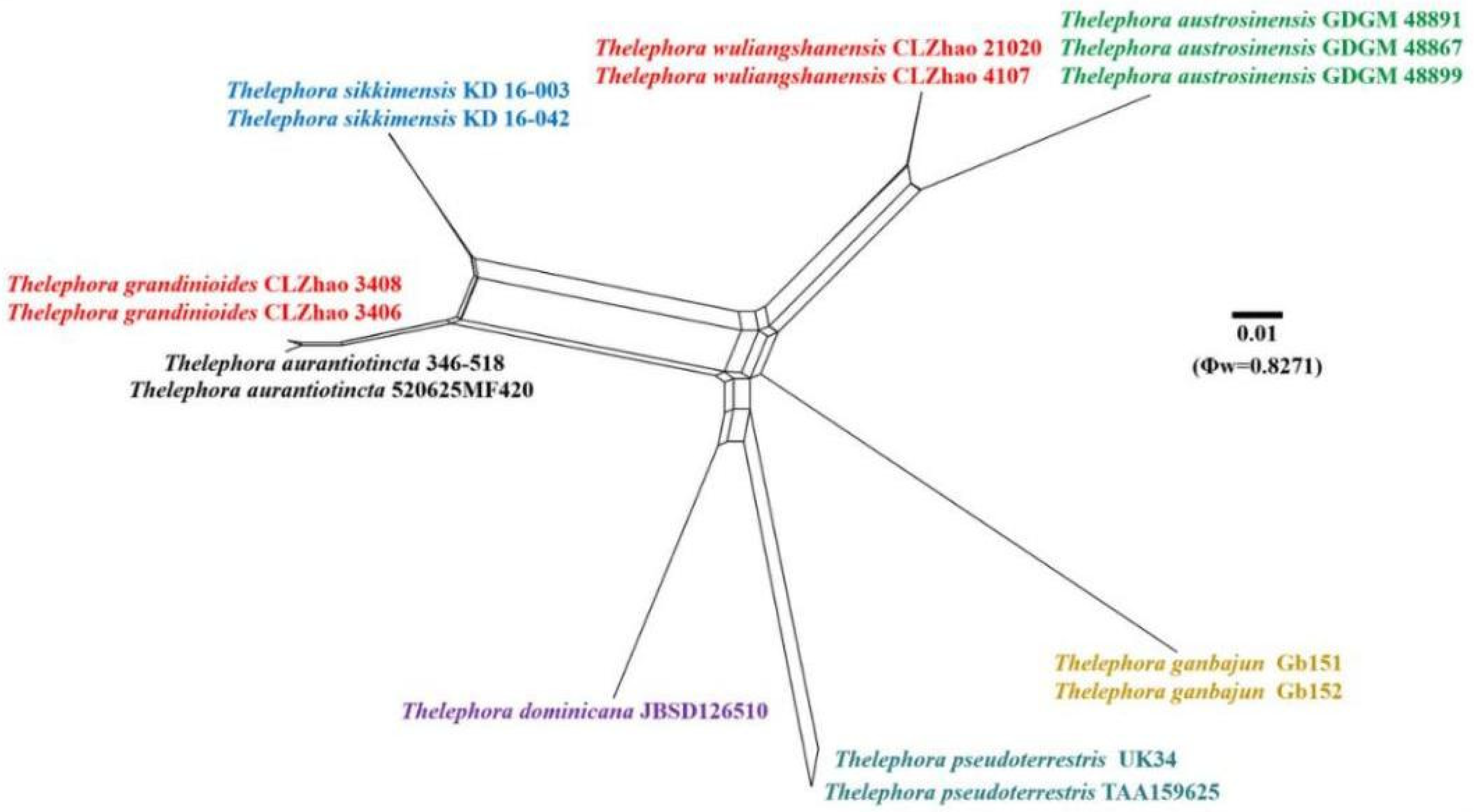
3.1. Molecular Phylogeny
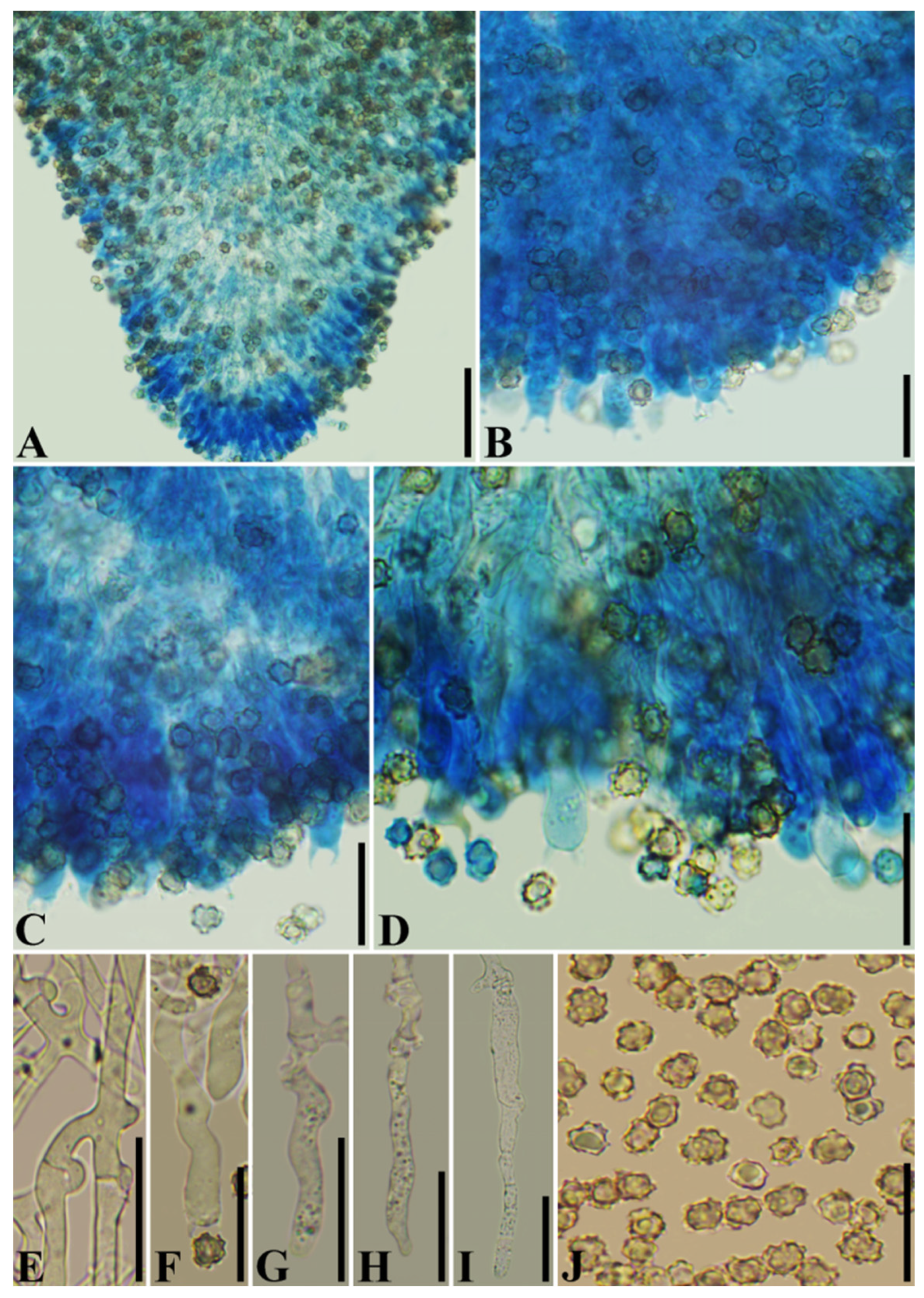
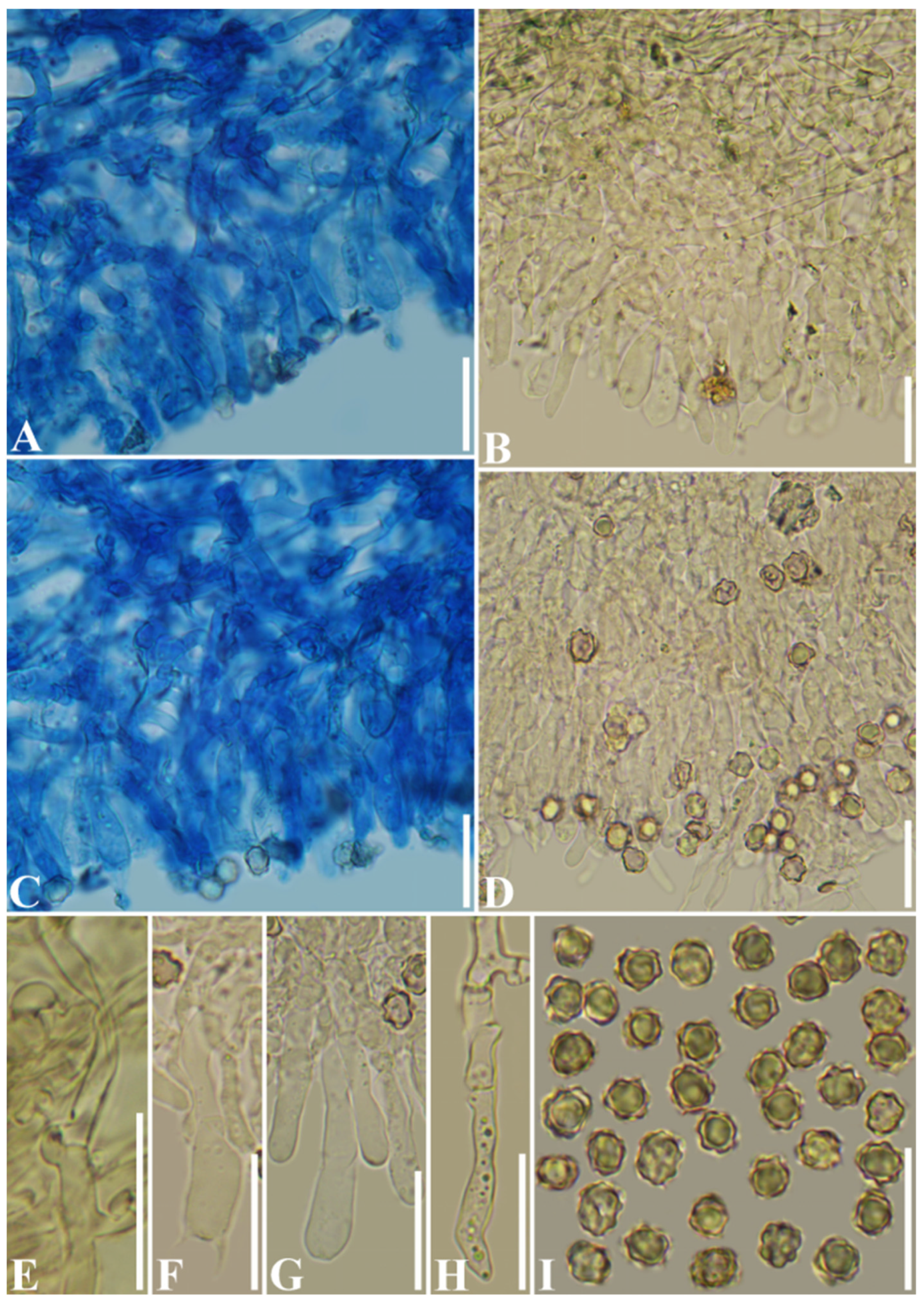
3.2. Taxonomy
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Cunningham, G.H. The Thelephoraceae of Australia and New Zealand; New Zealand Department of Science and Industrial Research: Wellington, New Zealand, 1963; pp. 228–239. [Google Scholar] [CrossRef]
- Burt, E.A. The Thelephoraceae of North America I-XV; Hafner Publishing Company, Inc.: New York, NY, USA; London, UK, 1966; pp. 199–229. [Google Scholar] [CrossRef]
- Corner, E.J.H. A monograph of Thelephora (Basidiomycetes). Nova Hedwig. 1968, 27, 1–110. [Google Scholar]
- Ramírez-López, I.; Villegas-Ríos, M.; Salas-Lizana, R. Thelephora versatilis and Thelephora pseudoversatilis: Two new cryptic species with polymorphic basidiomes inhabiting tropical deciduous and sub-perennial forests of the Mexican Pacific coast. Mycologia 2015, 107, 346–358. [Google Scholar] [CrossRef]
- Li, T.; Li, T.H.; Song, B.; Hosen, M.I. Thelephora austrosinensis (Thelephoraceae), a new species close to T. ganbajun from southern China. Phytotaxa 2020, 471, 208–220. [Google Scholar] [CrossRef]
- Hilszczanska, D.; Sierota, Z. Persistence of ectomycorrhizas by Thelephora terrestris on outplanted Scots pine seedlings. Acta Mycol. 2006, 41, 313–318. [Google Scholar] [CrossRef] [Green Version]
- Iwanski, M.; Rudawski, M.; Leski, T. Mycorrhizal associations of nursery grown scots pine (Pinus sylvestris L.) seedlings in poland. Ann. For. Sci. 2006, 63, 715–723. [Google Scholar] [CrossRef]
- Flykt, E.; Timonen, S.; Pennanen, T. Variation of ectomycorrhizal colonisation in Norway spruce seedlings in Finnish forest nurseries. China Pharm. 2008, 42, 571–585. [Google Scholar] [CrossRef] [Green Version]
- Sha, T.; Xu, J.; Palanichamy, M.G.; Zhang, H.B.; Li, T.; Zhao, Z.W.; Zhang, Y.P. Genetic diversity of the endemic gourmet mushroom Thelephora ganbajun from south-western China. Microbiology 2008, 154, 3460–3468. [Google Scholar] [CrossRef] [Green Version]
- Yorou, S.N. Miscellaneous Contributions to the Anatomy and Molecular Phylogeny of Tropical African Resupinate Thelephorales. Ph.D. Thesis, University of Munich, Munich, Germany, 2008; p. 120. [Google Scholar]
- He, J.; Zhou, Z.; Yang, H.; Xu, J. Integrative management of commercialized wild mushroom: A case study of Thelephora ganbajun in Yunnan, southwest China. Environ. Manag. 2011, 48, 98–108. [Google Scholar] [CrossRef] [PubMed]
- Yangdo, R.; Kumar, S.; Sharma, Y.P. Three hitherto unreported macrofungi from cold arid region of Ladakh Province, Jammu and Kashmir, India. Kavaka 2015, 44, 42–44. [Google Scholar]
- Ezhov, O.N.; Zmitrovich, I.V. Lignotrophic basidiomycetes from pioneering microsites in boreal forests of the White Sea Region. Byulleten Moskovskogo Obshchestva Ispytateley Prirody. Otd. Biol. 2017, 122, 44–50. [Google Scholar]
- Zmitrovich, I.V.; Shchepin, O.N.; Malysheva, V.F.; Kalinovskaya, N.I.; Volobuev, S.V.; Myasnikov, A.G.; Ezhov, O.N.; Novozhilov, Y.K. Basidiome reduction in litter-inhabiting Thelephorales in boreal forest environments: Morphological and molecular evidence. Curr. Res. Environ. Appl. Mycol. 2018, 8, 360–371. [Google Scholar] [CrossRef]
- Bi, Z.S.; Zheng, G.Y.; Li, T.H. The Macrofungus Flora of China’s Guangdong Province; Chinese University Press: Hong Kong, 1993; p. 734. [Google Scholar]
- Teng, S.C. Notes on the Thelephoraceae and Hydnaceae from China. Sinensia 1934, 6, 9–13. [Google Scholar]
- Tai, F.L. Sylloge Fungorum Sinicorum; Science Press, Academia Sinica: Beijing, China, 1979; pp. 740–741. [Google Scholar]
- Zang, M. Some new and noteworthy higher fungi from eastern Himalayas. Acta Bot. Yunnanica 1987, 9, 81–88. [Google Scholar]
- Dai, Y.C.; Wei, Y.L.; Zhang, X.Q. An annotated checklist of non-poroid Aphyllophorales in China. Ann. Bot. Fenn. 2004, 41, 233–247. [Google Scholar] [CrossRef]
- Qin, W.M.; Dai, Y.C.; Wu, X.L. Thelephora caryophyllea-a corticioid fungus new to China. Guizhou Sci. 2009, 27, 19–21. [Google Scholar]
- Li, Y.; Li, T.H.; Yang, Z.L.; Bau, T.; Dai, Y.C. Atlas of Chinese Macrofungal Resources; Central China Farmers Publishing House: Zhengzhou, China, 2015; p. 1351. [Google Scholar]
- Willdenow, C.L. Florae Berolinensis Prodromus: Secundum Systema Linneanum ab Illustr. Viro ac eq. C.P. Thunbergio EMENDATUM Conscriptus; Impensis Wilhelmi Viewegii, Berolini: Berlin, Germany, 1787; p. 490. [Google Scholar]
- Kirk, P.M.; Cannon, P.; Stalpers, J. (Eds.) Dictionary of the Fungi, 10th ed.; CABI: Wallingford, UK, 2008; p. 686. [Google Scholar]
- Zecchin, G. Ⅱ genere Thelephora in Friuli-Ottavo contributo. Riv. Micol. 2008, 51, 117–125. [Google Scholar]
- Vizzini, A.; Angelini, C.; Losi, C.; Ercole, E. Thelephora dominicana (Basidiomycota, Thelephorales), a new species from the Dominican Republic, and preliminary notes on thelephoroid genera. Phytotaxa 2016, 265, 27–38. [Google Scholar] [CrossRef]
- Das, K.; Hembrom, M.E.; Ghosh, A.; Parihar, A.; Kuhar, F. Thelephora sikkimensis sp. nov. (Thelephoraceae) from the Eastern Himalayas (India). Nova Hedwig. 2018, 107, 337–347. [Google Scholar] [CrossRef]
- Khalid, N.A.; Hanif, M. Thelephora iqbalii sp. nov. from the himalayan moist temperate forests of Pakistan. Mycotaxon 2018, 132, 943–950. [Google Scholar] [CrossRef]
- Index Fungorum. 2021. Available online: http://www.indexfungorum.org/Names/Names.asp (accessed on 3 December 2021).
- Stalpers, J.A. The Aphyllophoraceous Fungi I. Keys to the species of the Thelephorales. Stud. Mycol. 1993, 35, 1–168. [Google Scholar]
- Yorou, N.S.; Agerer, R. Tomentella furcata, a new species from Benin (West Africa) with basidia forming internal hyphae. Mycol. Prog. 2007, 6, 239–247. [Google Scholar] [CrossRef]
- Larsson, K.H.; Larsson, E.; Kljalg, U. High phylogenetic diversity among corticioid homobasidiomycetes. Mycol. Res. 2004, 108, 983–1002. [Google Scholar] [CrossRef]
- Kuhar, F.; Barroetaveña, C.; Rajchenberg, M. New species of Tomentella (Thelephorales) from the patagonian andes forests. Mycologia 2016, 108, 780–790. [Google Scholar] [CrossRef]
- Kõljalg, U. Tomentella (Basidiomycota) and related genera in temperate Eurasia. Synop. Fungorum 1996, 9, 1–213. [Google Scholar]
- Patouillard, N. Les Hyménomycètes d’Europe; Paul Klincksieck: Paris, France, 1887; p. 166. [Google Scholar]
- Petersen, J.H. Farvekort. The Danish Mycological Society’s Colour-Chart; Foreningen til Svampekundskabens Fremme: Greve, Denmark, 1996; pp. 1–6. [Google Scholar]
- Dai, Y.C. Polypore diversity in China with an annotated checklist of Chinese polypores. Mycoscience 2012, 53, 49–80. [Google Scholar] [CrossRef]
- Zhao, C.L.; Wu, Z.Q. Ceriporiopsis kunmingensis sp. nov. (Polyporales, Basidiomycota) evidenced by morphological characters and phylogenetic analysis. Mycol. Prog. 2017, 16, 93–100. [Google Scholar] [CrossRef]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. PCR Protoc. Guide Methods Appl. 1990, 18, 315–322. [Google Scholar] [CrossRef]
- Vilgalys, R.; Hester, M. Rapid genetic identification and mapping of enzymatically amplified ribosomal DNA from several Cryptococcus species. J. Bacteriol. 1990, 172, 4239–4246. [Google Scholar] [CrossRef] [Green Version]
- Hall, T.A. BioEdit: A user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucleic Acids Symp. Ser. 1999, 41, 95–98. [Google Scholar] [CrossRef]
- Kernaghan, G.; Sigler, L.; Khasa, D. Mycorrhizal and root endophytic fungi of containerized Picea glauca seedlings assessed by rDNA sequence analysis. Microb. Ecol. 2003, 45, 128–136. [Google Scholar] [CrossRef]
- Larsson, K.H.; Svantesson, S.; Miscevic, D.; Kljalg, U.; Larsson, E. Reassessment of the generic limits for Hydnellum and Sarcodon (thelephorales, basidiomycota). MycoKeys 2019, 54, 31–47. [Google Scholar] [CrossRef]
- Zhao, R.L.; Li, G.J.; Sánchez-Ramírez, S.; Stata, M.; Yang, Z.L.; Wu, G.; Dai, Y.C.; He, S.H.; Cui, B.K.; Zhou, J.L.; et al. A six-gene phylogenetic overview of Basidiomycota and allied phyla with estimated divergence times of higher taxa and a phyloproteomics perspective. Fungal Divers. 2017, 84, 1–32. [Google Scholar] [CrossRef]
- Palmer, J.M.; Lindner, D.L.; Volk, T.J. Ectomycorrhizal characterization of an american chestnut (Castanea dentata)-dominated community in western wisconsin. Mycorrhiza 2008, 19, 27–36. [Google Scholar] [CrossRef]
- Weiß, M.; Oberwinkler, F. Phylogenetic relationships in auriculariales and related groups-hypotheses derived from nuclear ribosomal dna sequences. Mycol. Res. 2001, 105, 403–415. [Google Scholar] [CrossRef]
- Taylor, D.L.; Bruns, T.D. Independent, specialized invasions of ectomycorrhizal mutualism by two nonphotosyntheti corchids. Proc. Natl. Acad. Sci. USA 1997, 94, 4510–4515. [Google Scholar] [CrossRef] [Green Version]
- Vu, D.; Groenewald, M.; De, V.M.; Gehrmann, T.; Stielow, B.; Eberhardt, U.; Al-Hatmi, A.; Groenewald, J.Z.; Cardinali, G.; Houbraken, J.; et al. Large-Scale generation and analysis of filamentous fungal dna barcodes boosts coverage for kingdom fungi and reveals thresholds for fungal species and higher taxon delimitation. Stud. Mycol. 2019, 92, 135–154. [Google Scholar] [CrossRef]
- Swofford, D.L. PAUP*: Phylogenetic Analysis Using Parsimony (* and Other Methods); Version 4.0b10; Sinauer Associates: Sunderland, MA, USA, 2002. [Google Scholar]
- Felsenstein, J. Confidence intervals on phylogenetics: An approach using bootstrap. Evolution 1985, 39, 783–791. [Google Scholar] [CrossRef]
- Miller, M.A.; Holder, M.T.; Vos, R.; Midford, P.E.; Liebowitz, T.; Chan, L.; Hoover, P.; Warnow, T. The CIPRES Portals. CIPRES. 2009. Available online: http://www.phylo.org/sub_sections/portal (accessed on 3 December 2021).
- Nylander, J.A.A. MrModeltest v2. Program Distributed by the Author; Evolutionary Biology Centre, Uppsala University: Uppsala, Sweden, 2004. [Google Scholar]
- Ronquist, F.; Huelsenbeck, J.P. MrBayes 3: Bayesian phylogenetic inference under mixed models. Bioinformatics 2003, 19, 1572–1574. [Google Scholar] [CrossRef] [Green Version]
- Bruen, T.C.; Philippe, H.; Bryant, D. A simple and robust statistical test for detecting the presence of recombination. Genetics 2006, 172, 2665–2681. [Google Scholar] [CrossRef] [Green Version]
- Huson, D.H.; Bryant, D. Application of phylogenetic networks in evolutionary studies. Mol. Biol. Evol. 2006, 23, 254–267. [Google Scholar] [CrossRef]
- Quaedvlieg, W.; Binder, M.; Groenewald, J.Z.; Summerell, B.A.; Carnegie, A.J.; Burgess, T.I.; Crous, P.W. Introducing the consolidated species concept to resolve species in the Teratosphaeriaceae. Persoonia 2014, 33, 1–40. [Google Scholar] [CrossRef] [Green Version]
- Coker, W.C. Notes on the Thelephoraceae of North Carolina. J. Elisha Mitchell Sci. Soc. 1921, 36, 146–196. [Google Scholar]
- Lentz, P.L. The genus Thelephora in Iowa. Proc. Iowa Acad. Sci. 1942, 49, 175–184. [Google Scholar]
- Global Biodiversity Information Facility (GBIF). 2021. Available online: https://www.gbif.org/species/search?q=fungi&dataset_key (accessed on 3 December 2021).
- Vidal, J.M.; Siquier, J.L.; Constantino, C. Alguns macromicets nous o interessants de l’illa de Mallorca (Balears). Rev. Catalana Micol. 1994, 16, 135–144. [Google Scholar]
- Dai, Y.C. A revised checklist of corticioid and hydnoid fungi in China for 2010. Mycoscience 2011, 52, 69–79. [Google Scholar] [CrossRef]
- Landi, M.; Salerni, E.; Ambrosio, E.; D’Aguanno, M.; Nucci, A.; Saveri, C.; Perini, C.; Angiolini, C. Concordance between vascular plant and macrofungal community composition in broadleaf deciduous forests in central Italy. Iforest Biogeosci. For. 2014, 3, 279–286. [Google Scholar] [CrossRef] [Green Version]
- Czernyadjeva, I.V.; Afonina, O.M.; Davydov, E.A.; Doroshina, G.Y.; Zyatnina, V. New cryptogamic records. 5. Nov. Sist. Nizshikh Rastenii 2020, 54, 261–286. [Google Scholar] [CrossRef]
- Hernández-López, L.; Rodríguez-Alcántar, O.; Figueroa-García, D.; Reynoso-Dueñas, J.J.; Arias, A. Threatened and endemic flora and mycobiota in the municipality San Sebastián del Oeste, Jalisco, Mexico. Acta Bot. Mex. 2021, 128, 2448–7589. [Google Scholar] [CrossRef]
- Vila, J.; Angel, F.; Mayoral, A.; Hoyo, P. Segona aportació al coneixement dels macromicets de la zona de Garraf. Rev. Catalana Micol. 1998, 21, 45–61. [Google Scholar]
- Mleczko, P. Mycorrhizal and saprobic macrofungi of two zinc wastes in southern poland. Acta Biol. Crac. Ser. Bot. 2004, 46, 25–38. [Google Scholar] [CrossRef] [Green Version]
- Abrahão, M.C.; Gugliotta, A.D.M.; Bononi, V.L.R. Xylophilous Agaricomycetes (Basidiomycota) of the Brazilian Cerrado. Check List 2012, 8, 1102–1116. [Google Scholar] [CrossRef] [Green Version]
- Liu, T.Z.; Li, G.L. Atlas of Macrofungi in Saihanwula National Nature Reserve of Inner Mongolia; Inner Mongolia Science Press: Hohhot, China, 2019; p. 341. [Google Scholar]
- Chanonagómez, F.; Andradegallegos, R.H.; Castellanosalbores, J.; Sánchez, J.E. Macromicetos del parque educativo laguna bélgica, muncipio de ocozocoautla de espinosa, chiapas, méxico. Rev. Mex. Biodivers. 2007, 78, 369–381. [Google Scholar] [CrossRef]
- Chacón, S.; Tapia, F.; Jarvio, D. Four interesting aphyllophoroid species in the tropical northern region of Veracruz, Mexico. Mycotaxon 2018, 133, 153–163. [Google Scholar] [CrossRef]
- Budathoki, U.S.H.A. A new species of Periconiella from Kathmandu valley, Nepal. Recent Mycol. Res. 2009, 77–80. [Google Scholar]
- Roberts, P.J.; Spooner, B.M. Cantharelloid, clavarioid and thelephoroid fungi from Brunei Darussalam. Kew Bull. 2000, 55, 843–851. [Google Scholar] [CrossRef]
- Welden, A.L. West Indian species of dark-spored Thelephoraceae. Sydowia 1968, 22, 269–273. [Google Scholar]
- An, G. The excellent edible mushrooms in Yunnan—The exploitation and utilization of Thelephora vialis schw and T. aurantiotincta Corner. J. Southwest For. Coll. 1998, 18, 105–108. [Google Scholar]
- Kõljalg, U.; Dahlberg, A.; Taylor, A.F.S.; Larsson, E.; Hallenberg, J.; Stenlid, K.H.; Larsson, P.M.; Fransson, O.; Kårén, L.; Jonsson, L. Diversity and abundance of resupinate thelephoroid fungi as ectomycorrhizal symbionts in swedish boreal forests. Mol. Ecol. 2000, 9, 1985–1996. [Google Scholar] [CrossRef]
- Cunningham, G.H. Thelephoraceae of New Zealand (parts XII and XIII). Part XII: The genera Thelephora and Tomentella. Trans. R. Soc. N. Z. 1957, 84, 479–496. [Google Scholar]
- Ramirez-Lópezl, I.; Villegas-Ríos, M.; Cano-Santana, Z. Phenotypic plasticity of the basidiomata of Thelephora sp. (Thelephoraceae) in tropical forest habitats. Rev. Biol. Trop. 2013, 61, 343–350. [Google Scholar] [CrossRef] [Green Version]
- Erland, S.; Taylor, A.F.S. Resupinate ectomycorrhizal fungal genera. In Ectomycorrhizal Fungi: Key Genera in Profile; Springer: Berlin/Heidelberg, Germany, 1999; p. 369. [Google Scholar] [CrossRef]
- Zmitrovich, I.V.; Wasser, S.P.; Ţura, D. Wood-Inhabiting fungi. In Fungi from Different Substrates; Misra, J.K., Tewari, J.P., Deshmukh, S.K., Vágvölgyi, C., Eds.; CRC Press, Taylor and Francis Group: New York, NY, USA, 2015; pp. 17–74. [Google Scholar]
- Teng, S.C. Fungi of China; Science Press: Beijing, China, 1963; p. 808. [Google Scholar]
- Lin, H.; Liu, J.K. p-Terphenyls from the basidiomycete Thelephora aurantiotincta. Z. Nat. C J. Biosci. 2003, 58, 452–454. [Google Scholar] [CrossRef]
- Wang, X.H.; Liu, P.G. Resources investigation and study on the wild commercial fungi in Yunnan Province. Biodivers. Sci. 2002, 10, 318–3325. [Google Scholar] [CrossRef]
- Barad, A.A. British Columbian Wild Mushrooms as Potent Source of Novel Natural Anti-Cancer Compounds. Ph.D. Thesis, University of Northern British Columbia, Prince George, BC, Canada, 2017. [Google Scholar]
- Liu, J.K.; Lin, H.; Dong, Z.J.; Hu, Q. Dpph radical scavenging activity of ten natural p-terphenyl derivatives obtained from three edible mushrooms indigenous to china. Chem. Biodivers. 2010, 1, 601–605. [Google Scholar] [CrossRef] [PubMed]
- Norikura, T.; Fujiwara, K.; Narita, T.; Yamaguchi, S.; Morinaga, Y.; Kunihisa, I.; Matsue, H. Anticancer activities of thelephantin o and vialinin an isolated from Thelephora aurantiotincta. J. Agric. Food Chem. 2011, 59, 6974–6979. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.H.; Meng, W.W.; Liu, R.C.; Bai, Y.X.; Xu, H.Q.; Ding, R.; Shao, S.C. Complete mitochondrial genome of the edible Basidiomycete mushroom Thelephora aurantiotincta (Aphyllophorales: Thelephoraceae) from China. Mitochondrial DNA Part B 2021, 6, 606–607. [Google Scholar] [CrossRef]
- Hu, L.; Gao, J.M.; Liu, J.K. From fruiting bodies of the basidiomycete Thelephora ganbajun. Helv. Chim. Acta 2001, 84, 3342–3349. [Google Scholar] [CrossRef]
- Hu, L.; Liu, J.K. Two novel phenylacetoxylated p-terphenyls from Thelephora ganbajun Zang. Z. Nat. Sect. C J. Biosci. 2001, 56, 983–987. [Google Scholar] [CrossRef]
- Xu, D.P.; Jie, Z.; Yue, Z.; Li, Y.; Li, S.; Li, H.B. Extraction of natural antioxidants from the Thelephora ganbajun mushroom by an ultrasound-assisted extraction technique and evaluation of antiproliferative activity of the extract against human cancer cells. Int. J. Mol. Sci. 2016, 17, 1664. [Google Scholar] [CrossRef] [Green Version]
- Zhang, L.; Zhang, A.L.; Li, X.M.; Wang, J.; Gao, J.M. Preliminary study of the anti-microbial activities of fermentation broth of Thelephora japonica. J. Northwest For. Univ. 2007, 22, 135–137. [Google Scholar]
- Radulovic, N.; Dang, N.Q.; Hashimoto, T.; Nukada, M.; Asakawa, Y. Terrestrins A–G: Pmilerphenyl derivatives from the inedible mushroom Thelephora terrestris. Phytochemistry 2005, 66, 1052–1059. [Google Scholar] [CrossRef]
- Xie, C.; Koshino, H.; Esumi, Y.; Onose, J.; Yoshikawa, K.; Abe, N. Vialinin B, a novel potent inhibitor of tnf-alpha production, isolated from an edible mushroom, Thelephora vialis. Bioorg. Med. Chem. Lett. 2006, 16, 5424–5426. [Google Scholar] [CrossRef] [PubMed]
- Xie, C.; Koshino, H.; Esumi, Y.; Takahashi, S.; Abe, N. Vialinins A and B: Novel bioactive compounds from Thelephora vialis, an edible mushroom in China. ACS Symp. 2008, 993, 465–472. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
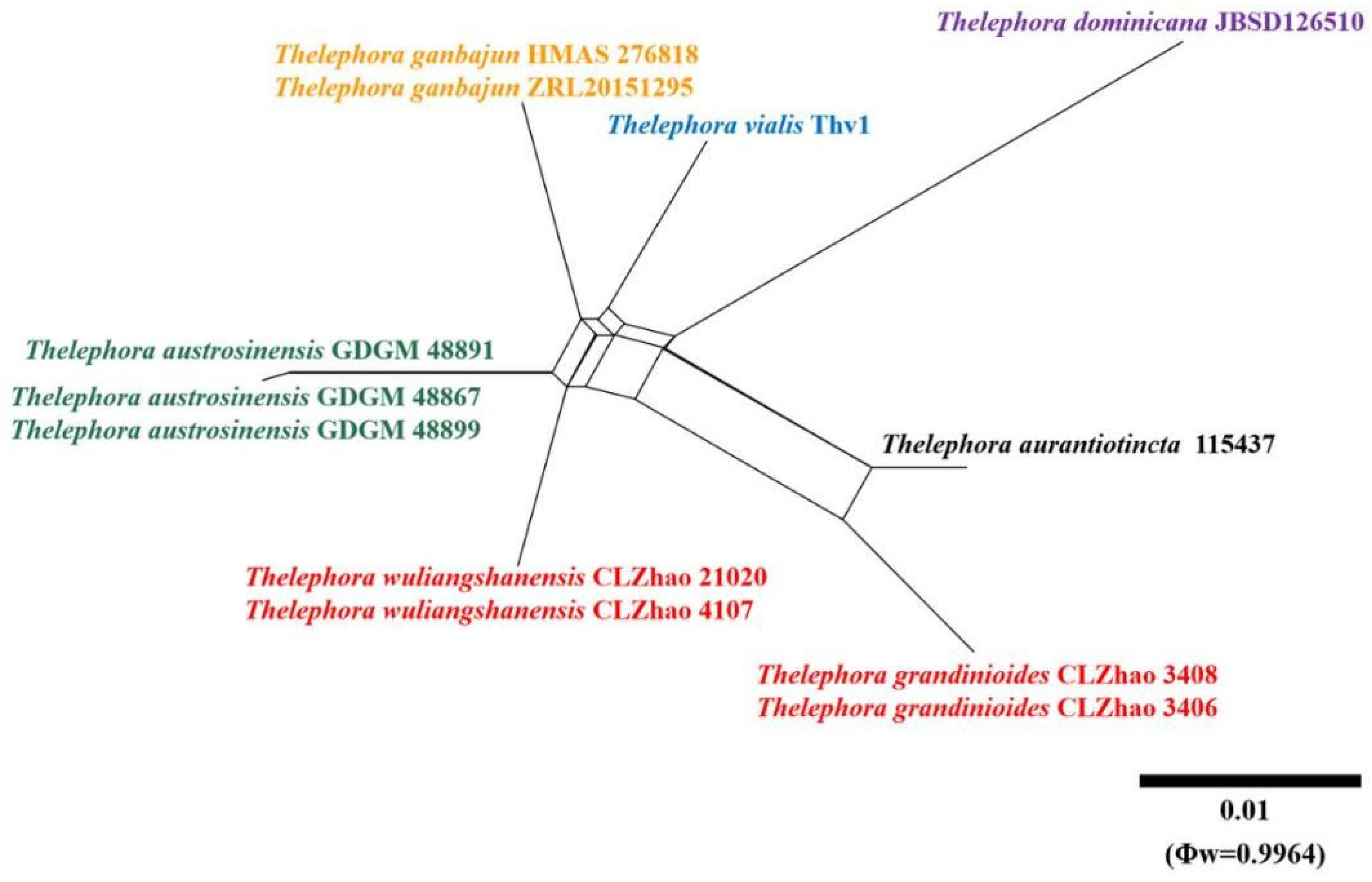{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Taxon Names | Voucher | Location | GenBank/UNITE Accession Number | ||
|---|---|---|---|---|---|
| ITS | nrLSU | Reference | |||
| Odontia ferruginea | UK18 | Estonia | UDB000285 | UDB018691 | [25] |
| O. fibrosa | SS38 | Sweden | MH310788 | UDB018463 | [25,26] |
| Thelephora albomarginata | KHL8457 | Sweden | – | UDB018707 | [5] |
| T. americana | UAMH 9578 | Chile | AY219838 | – | [41] |
| T. anthocephala | UBC F28410 | Canada | KP454019 | KP454019 | [26] |
| T. anthocephala | TAA165304 | Estonia | AF272927 | UDB018693 | [27] |
| T. atra | UK50 | Russia | – | UDB018697 | UNITE |
| T. aurantiotincta | 115437 | – | – | TU115437 | UNITE |
| T. aurantiotincta | 520625MF420 | China | MZ057686 | – | GenBank |
| T. aurantiotincta | 346–518 | Japan | AB509809 | – | GenBank |
| T. austrosinensis | GDGM 48867 T | China | MF593265 | MF593265 | [5] |
| T. austrosinensis | GDGM 48891 | China | MF593266 | MF593266 | [5] |
| T. austrosinensis | GDGM 48899 | China | MF593267 | MF593267 | [5] |
| T. caryophyllea | ELarsson89-09 | Sweden | MK602776 | MK602776 | [42] |
| T. caryophyllea | TAAM172626 | Estonia | – | UDB018694 | [5] |
| T. caryophyllea | TL-6566 | Denmark | AJ889980 | – | [27] |
| T. caryophyllea | GO-2010-163 | Mexico | KC152242 | – | [26] |
| T. caryophyllea | TAAM172626 | Estonia | UDB018694 | – | [5] |
| T. dominicana | JBSD126510 T | Dominican Republic | KX216400 | KX216400 | [25] |
| T. ganbajun | Gb151 | China | EU696873 | – | [9] |
| T. ganbajun | Gb152 | China | EU696874 | – | [9] |
| T. ganbajun | HMAS 276818 | China | – | LC164937 | GenBank |
| T. ganbajun | ZRL20151295 | China | – | KY418908 | [43] |
| T. grandinioides | CLZhao 3406 T | China | MZ400673 | MZ400675 | Present study |
| T. grandinioides | CLZhao 3408 | China | MZ400674 | MZ400676 | Present study |
| T. iqbalii | MH810 T | Pakistan | JX241471 | – | [27] |
| T. japonica | 420526MF0417 | China | – | MG712350 | GenBank |
| T. palmata | JMP0085 | USA | EU819443 | – | [44] |
| T. palmata | LW 84 | – | – | AF291265 | [45] |
| T. palmata | TAA149550 | Swedish | AF272919 | [44] | |
| T. palmata | Telpa31/38 | – | AJ406477 | [5] | |
| T. aff. palmata | 350–421 | Japan | AB509755 | – | [27] |
| T. penicillata | 0465 | China | MT325773 | – | [26] |
| T. penicillata | LTT8 | USA | U83484 | – | [46] |
| T. penicillata | TAAM169453 | Estonia | – | UDB018695 | [5] |
| T. pseudoterrestris | TAA159625 | Estonia | AF272907 | – | [27] |
| T. pseudoterrestris | UK34 | – | UDB000209 | – | [25] |
| T. pseudoversatilis | 11H2-1 | Mexico | KU530339 | – | [26] |
| T. pseudoversatilis | FCME 26152 T | Mexico | KJ462486 | – | [4] |
| T. pseudoversatilis | FCME 26232 | Mexico | JX075890 | JX514167 | [4] |
| T. regularis | UBC F33227 | Canada | MG953966 | – | [26] |
| T. regularis | JMT17371 | USA | U83485 | – | [46] |
| T. aff. regularis | GO-2010-125 | Mexico | KC152240 | – | [26] |
| T. aff. regularis | GO-2010-134 | Mexico | KC152241 | – | [26] |
| T. sikkimensis | KD 16-003 | India | MF684017 | – | [26] |
| T. sikkimensis | KD 16-042 | India | MF684018 | – | [26] |
| T. sublilacina | UP161 | Sweden | EF493288 | – | [27] |
| T. terrestris | CBS 703.85 | Netherlands | – | MH873600 | [47] |
| T. terrestris | Hilszczanska D. 1-IBL | Poland | FJ532478 | – | [4] |
| T. terrestris | P17_M2_772 | Poland | KM409440 | – | [26] |
| T. terrestris | UK14 | Estonia | – | DB018696 | [5] |
| T. versatilis | MEXU:27094 | Mexico | KC595628 | – | [4] |
| T. versatilis | UNAM:FCME26141 T | Mexico | NR154492 | – | [4] |
| T. vialis | Thv1 | – | – | AJ406478 | [28] |
| T. wuliangshanensis | CLZhao 4107 T | China | MZ400671 | MZ400677 | Present study |
| T. wuliangshanensis | CLZhao 21020 | China | MZ400672 | MZ400678 | Present study |
| Species | Thelephora grandinioides | Thelephora wuliangshanensis | ||||||
|---|---|---|---|---|---|---|---|---|
| Specimens | CLZhao 3406 | CLZhao 3408 | CLZhao 4107 | CLZhao 21020 | ||||
| Gene | ITS (bp) | nrLSU (bp) | ITS (bp) | nrLSU (bp) | ITS (bp) | nrLSU (bp) | ITS (bp) | nrLSU (bp) |
| T. aurantiotincta 115437 | NA | 13 | NA | 13 | NA | 24 | NA | 24 |
| T. aurantiotincta 346–518 | 1 | NA | 1 | NA | 42 | NA | 40 | NA |
| T. aurantiotincta 520625MF420 | 8 | NA | 8 | NA | 78 | NA | 76 | NA |
| T. austrosinensis GDGM 48891 | 89 | 28 | 89 | 28 | 36 | 17 | 36 | 17 |
| T. austrosinensis GDGM 48867 | 89 | 29 | 89 | 29 | 37 | 18 | 37 | 18 |
| T. austrosinensis GDGM 48899 | 87 | 30 | 87 | 30 | 35 | 18 | 35 | 18 |
| T. dominicana JBSD126510 | 84 | 32 | 83 | 32 | 79 | 26 | 78 | 26 |
| T. ganbajun Gb151 | 98 | NA | 98 | NA | 53 | NA | 52 | NA |
| T. ganbajun Gb152 | 85 | NA | 85 | NA | 78 | NA | 77 | NA |
| T. ganbajun HMAS 276818 | NA | 29 | NA | 29 | NA | 18 | NA | 18 |
| T. ganbajun ZRL20151295 | NA | 27 | NA | 27 | NA | 18 | NA | 18 |
| T. pseudoterrestris UK34 | 87 | NA | 87 | NA | 48 | NA | 46 | NA |
| T. pseudoterrestris TAA159625 | 64 | NA | 64 | NA | 48 | NA | 46 | NA |
| T. sikkimensis KD1603 | 45 | NA | 45 | NA | 76 | NA | 75 | NA |
| T. sikkimensis KD1642 | 43 | NA | 43 | NA | 77 | NA | 76 | NA |
| T. vialis Thv1 | NA | 26 | NA | 26 | NA | 20 | NA | 20 |
| Thelephora grandinioides (Holotype CLZhao 3406) | Thelephora wuliangshanensis (Holotype CLZhao 4107) | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| ITS | ITS | ||||||||||
| Species | Max Score | Total Score | Query Cover | E Value | Ident | Species | Max Score | Total Score | Query Cover | E Value | Ident |
| T. aurantiotincta | 1109 | 1109 | 94% | 0.0 | 98.87% | T. ganbajun | 1011 | 1011 | 97% | 0.0 | 95.32% |
| T. sikkimensis | 907 | 907 | 96% | 0.0 | 92.36% | T. ganbajun | 1005 | 1005 | 97% | 0.0 | 95.16% |
| T. sikkimensis | 872 | 872 | 91% | 0.0 | 92.61% | T. ganbajun | 1000 | 1000 | 97% | 0.0 | 95.01% |
| To. lateritia | 806 | 806 | 98% | 0.0 | 89.12% | T. ganbajun | 1000 | 1000 | 97% | 0.0 | 95.01% |
| To. cf. ramosissima | 802 | 802 | 97% | 0.0 | 89.13% | T. ganbajun | 1000 | 1000 | 97% | 0.0 | 95.01% |
| To. ramosissima | 802 | 802 | 97% | 0.0 | 89.16% | T. ganbajun | 1000 | 1000 | 97% | 0.0 | 95.01% |
| To. bryophila | 798 | 798 | 98% | 0.0 | 88.84% | T. cf. ganbajun | 985 | 985 | 95% | 0.0 | 95.22% |
| To. ramosissima | 793 | 793 | 96% | 0.0 | 89.15% | T. ganbajun | 976 | 976 | 97% | 0.0 | 94.24% |
| To. fuscocinerea | 793 | 793 | 96% | 0.0 | 88.96% | T. ganbajun | 976 | 976 | 97% | 0.0 | 94.25% |
| T. sp. | 987 | 987 | 82% | 0.0 | 99.27% | T. cf. ganbajun | 974 | 974 | 95% | 0.0 | 94.42% |
| nrLSU | nrLSU | ||||||||||
| Species | Max Score | Total Score | Query Cover | E Value | Ident | Species | Max Score | Total Score | Query Cover | E Value | Ident |
| To. stuposa | 2375 | 2375 | 98% | 0.0 | 97.89% | T. terrestris | 2342 | 2342 | 99% | 0.0 | 97.32% |
| T. terrestris | 2342 | 2342 | 98% | 0.0 | 97.45% | T. terrestris | 2331 | 2331 | 99% | 0.0 | 97.17% |
| T. caryophyllea | 2340 | 2340 | 98% | 0.0 | 97.38% | T. caryophyllea | 2329 | 2329 | 99% | 0.0 | 97.10% |
| T. terrestris | 2331 | 2331 | 98% | 0.0 | 97.31% | To. stuposa | 2320 | 2320 | 99% | 0.0 | 97.03% |
| Pseudotomentella griseopergamacea | 2183 | 2183 | 98% | 0.0 | 95.35% | P. griseopergamacea | 2180 | 2180 | 99% | 0.0 | 95.15% |
| P. flavovirens | 2165 | 2165 | 98% | 0.0 | 95.14% | P. humicola | 2172 | 2172 | 99% | 0.0 | 95.01% |
| Odontia parvospora | 2165 | 2165 | 96% | 0.0 | 95.77% | P. flavovirens | 2170 | 2170 | 99% | 0.0 | 95.08% |
| O. parvospora | 2165 | 2165 | 96% | 0.0 | 95.77% | To. pulchella | 2156 | 2156 | 99% | 0.0 | 94.94% |
| P. humicola | 2161 | 2161 | 98% | 0.0 | 94.99% | P. tristis | 2128 | 2128 | 99% | 0.0 | 94.50% |
| To. pulchella | 2139 | 2139 | 98% | 0.0 | 94.84% | P. tristis | 2122 | 2122 | 98% | 0.0 | 94.67% |
| Species | Basidiospore (μm) | Basidia (μm) | Cystidia (μm) | Host/Substrate | Location | Reference | |
|---|---|---|---|---|---|---|---|
| Sterigmata | |||||||
| Thelephora albidobrunnea | 8.5–10.5(–11) × 6–8.5 | 55–80 × 11–13 | 4 | On the ground; Acer rubrum L., Amelanchier canadensis (L.) Medik., Amelanchier sp., Carpinus caroliniana Walter | Canada, USA | [3,56,57,58] | |
| T. alta | 7.6–8.5 × 6–7 | On the ground | Borneo, Brunei Darussalam, Indonesia, Malaysia | [3,58,59] | |||
| T. anthocephala | (7–)8–10(–11) × (5–)6–8.5 | 40–80 × 7–11 | 2–4 | On the ground in woods; Fagus sp., Queercus sp. | Austria, China, Denmark, Italy, Netherlands, North Temperate, Norway, Russia, Spain, Slovenia, Sweden, USA, UK | [3,58,60,61,62] | |
| T. arbuscula | 6–7 × 5.5–6 | On the ground in forest | India, Mexico, Papua New Guinea; Ukraine | [3,63] | |||
| T. atra | 9–13 × 8–11 | 50–100 × 9–12 | 2–3 | On the ground | Spain, Poland | [3,64,65] | |
| T. atrocitrina | 8–13 × 6.5–9 | 45–75 × 8–13 | 2–4 | On the ground in woods (Abies sp., Carpinus sp., Fagus sp., Quercus sp.) | Austria, Belgium, Brazilian, Czechoslovakia, France, Germany, Netherlands, Spain | [3,66] | |
| T. aurantiotincta | 6.5–9 × 5.5–6.5 | 43–55 × 6.5–8 | 4 | on the ground in humus in mountain in forest | China, Malaysia | [3,67] | |
| T. austrosinensis | (5.2–)5.7–6.3(–6.7) × (4.6–)5.0–5.4(–5.8) | 15–24 × 5–7 | 4 | Castanopsis chinensis (Spreng.) Hance, C. fabri Hance, C. fissa (Champ. ex Benth.) Rehder and E.H.Wilson, C. hystrix Hook.f. and Thomson ex A.DC., Lithocarpus polystachyus (Wall. ex A.DC.) Rehder, L. uvariifolius (Hance) Rehder, Schima superba Gardner and Champ. | China | [5] | |
| T. bresadolae | 5–7 | On the ground in woods | Hungary, Czechoslovakia | [3] | |||
| T. brunneoviolacea | 7.5–11 × 6–8.5 | 2–4 | On the ground in the forest | Congo | [3] | ||
| T. caespitulans | 7–8 × 5–6 | On the ground | Canada, USA | [3] | |||
| T. caryophyllea | 5–10 × 5–8 | 47–90 × 8–12 | 2–4 | On sandy ground in coniferous woods; Betula nigra L., Larix occidentalis Nutt., Pinus silvestris L., Salix sitchensis Sanson ex Bong. | Canada, China, Georgia, India, Mexico, North temperate, Poland, Russia, Spain, USA | [3,20,60,62,64,67] | |
| T. cerberea | 8–11 × 6–8.3 | 2–4 | On the ground in dry forest | Congo | [3] | ||
| T. cervicornis | 7–8.5 × 6.5–7.5 | On the ground | Bahamas, Mexico, USA | [3,68] | |||
| T. cervina | 6–7.5 × 5–6 | On the ground | China | [3] | |||
| T. congesta | 8.5–11 × 5.5–8.3 | 30–40 × 7–8 | 4 | On the ground | Australia | [3] | |
| T. crassitexta | 9–11 × 6–9 | 65–100 × 10–12 | 2–4 | On the wood | Borneo | [3] | |
| T. cuticularis | 7.5–12.5 × 6–9 | On mossy bark at the base of trees and on fallen twigs; Juniperus virginiana L. | UK, USA | [3,56,57] | |||
| T. cylindrica | 8–11 × 6.5–9 | 80–110 × 11–14 | 4 | On the ground in the forest | Japen, Java, Sumatra, | [3] | |
| T. dactylites | 6.5–8 × 5–7 | 4 | On the ground | China | [3] | ||
| T. dentosa | 7–9 × 6.5–8 | 18–46 × 8–13 | 4 | 19–36 × 4.5–7.5 | On dead twigs and leaves and encrusting parts pf living plants | Brazilian, Cuba; Haiti, Indian, Jamaica, Mexico | [3,66,69] |
| dominicana | 8–9.6 × 7.2–8.8 | (30–)50–60 × 10–12 | 4 | on deciduous forest litter; On the ground of deciduous forest; Coccoloba spp.; Gymnopodium floribundum Rolfe | Dominican Republic, Mexico | [14,25] | |
| T. erebia | 9–12 × 7–10 | 45–60 × 10–12 | 2 | On the ground in the forest | Malaysia | [3] | |
| T. fragilis | 6–7 × 4–5 | 14–28 × 5–7 | 2–4 | On the ground in the forest | Malaysia, Philippines | [3] | |
| T. fucoides | 7–9 × 6–8 | On the ground in the forest | India, Malaysia, Pakistan, USA | [3] | |||
| T. fuscella | 6–8 × 4.5–6 | 35–45 × 6–7 | 2–4 | On the ground; Symbiotic with plants | China, Europe, India, Japan, Malaysia, Nepal, North America, Singapore | [3,11,70] | |
| T. ganbajun | 7–12 × 6–8 | 25–35 × 9–12 | 4 | 52–80 × 7–14 | In the root of Pinus yunnanensis Fr. and Pinus kesiya var. langbianensis | China | [5,18,21] |
| T. gelatinoidea | 7–9.5 × 6–9 | 45–70 × 8–10 | 4 | On the ground in the forest | China, India, Malaysia | [3] | |
| gelidioides | 6–8 × 4.5–6 | On the ground in the forest | Singapore | [3] | |||
| T. grandinioides | (5–)5.3–7.4(–7.8) × (3.8–)4–6.5(–7) | 27–62 × 5–7.5 | 4 | 35–60 × 5–7.5 | On the ground of pine-broadleaved mixed forest | China | Present study |
| T. griseozonata | 8–12 × 5–8 | on sandy ground in pine wooods (Pinus sp.) | Germany, Puerto Rico, New Zealand, USA, Virgin Islands | [3,56] | |||
| T. intybacea | 8–12 × 6–9 | 45–90 × 9–12 | 2–4 | In pine woods; Cedrus deodara (Lamb.) G.Don, Pinus canariensis C.Sm. | European, New Zealand, North America, Southern Africa, Uruguay, USA | [3,56] | |
| T. investiens | 8.5–10 × 7–9.5 | 4 | On the ground in forest | Malaysia | [3] | ||
| T. japonica | (6–)7–10 × (5.5–)6–8 | 40–55 × 8–10 | 2–4 | On the ground, often encrusting small living plants; Ectomycorrhizal, humicolous, gregarious in mixed forest of Populus nigra L. and Salix alba L. | China, India, Japan, Malaysia | [3,12] | |
| T. lutosa | 5–6 × 3.5–4 | On the ground in roads and in woods | USA | [3,56] | |||
| T. luzonensis | 5–6.5 × 4.7–5.7 | 2–3 | China; Philippines, USA | [3] | |||
| T. magnifica | 9–12 × 7–9 | 60–75 × 8–10 | 2–3–4 | On ground in hill dipterocarp forest | Brunei, Malaysia | [3,71] | |
| T. magnispora | 11–12 × 9–11 | 50–70 × 10–11 | 4 | On mossy ground | Indian, Jamaica, USA | [3,72] | |
| T. mollissima | 8.5–10.7 × 6–7.7 | In woods | European, China | [3,60] | |||
| T. multipartita | 6–8.5(–9) × 4.7–7 | On the ground in frondose woods; Quercus sp. | Canada, China, Japan, USA, Venezuela | [3,56,60] | |||
| T. nigrescens | 6–9 × 5–8 | 30–50 × 7–10 | 4, rarely 2–4 | On the ground | Brunei Darussalam, China, Indonesia, Japan, Philippines | [3,58] | |
| T. palmata | 8–12 × 7–9 | 70–100 × 9–12 | 2–4 | On the ground in coniferous woods; Pinus sp.; Quercus humboldtii Bonpl. | China, Colombia, France, North temperate, Russia, Sweden, UK, USA | [3,56] | |
| T. paraguayensis | 6.5–8.5 × 4.5–7 | 4 | On the ground in woods | Brazil; Columbia; Paraguay | [3,58] | ||
| T. pendens | 8–10 × 7–8 | 40–50 × 10–12 | 4 | Rotten wood in the forest | Malaysia | [3] | |
| T. penicillata | 7–10 × 5–8 | 30–75 × 7–11 | On the ground and crusted leaves, branches, grass; Quercus coccifera L. | Canada, China, Galapagos Islands, India, Russia, UK | [3,60] | ||
| T. phyllophoroides | 5–7.5 × 4.5–5.5 | On the ground | Japan | [3] | |||
| T. pseudoterrestris | 9–11.5 × 7–9 | 50–60 × 12–14 | On the ground in the forest, leaves | Malaysia | [3] | ||
| T. pseudoversatilis | (6–)7–8(–8.5) × (5–)5.5–6(–7) | 47–55 × 9–12 | 4 | Sub-perennial tropical forest | Mexico | [4] | |
| T. ramarioides | 7–12.5 × 5–8.5 | 46–70 × 5–12 | 2–4, rarely 6–8 | 50–150 × 5–12 | On the ground under Casuarina equisetifolia L. | Australia, Borneo, Java, Malaysia | [3] |
| T. regularis | 6–8 × 4.5–6.5 | 35.2–102.4 × 6.4–9.6 | 4 | In moss in wet places and in humus; Ectomycorrhizal, humicolous, scattered under Salix excelsa J.F.Gmel., S. alba L. and Populus nigra L. | Canada, India, USA | [3,12,56] | |
| T. scissilis | 6–8.5 × 5–7 | 4 | On the ground in oak forest | Borneo, India, USA | [3] | ||
| T. sikkimensis | 6–(7.3)–8.8 × 5–(6.26)–7 | 35–65 × 7–9 | 4 | 30–50 × 3–10 | On the ground of Castanopsis hystrix | India | [26] |
| T. spiculosa | 8–12 × 7.5–9 | 60–70 × 9–12 | Encrusting conferous needles of Pinus sp. | France, Japan, Sweden, UK | [3,57] | ||
| T. tenuis | 7–8 × 6–7 | On sandy ground | China, Russia | [3] | |||
| T. terrestris | 8–12 × 6–9 | 40–90 × 8–12 | 2–4 | On the ground in coniferous, on roots, syumps and seedlings; Picea abies L.; Picea sitchensis (Bong.) Carr. | Australia, Bavaria, Brazil, China, Europe, Germany, Indian, Jamaica, Japan, Mexico, New Zealand, North America, South Africa, Spain, UK, Uruguay, USA | [3,21,60,64,67,68,72] | |
| T. versatilis | 6–7(8.5) × (4) 5–6(6.5) | 30–86 × (7)8–11 | 4 | Deciduous and sub-perennial tropical forest | Mexico | [4] | |
| T. vialis | 4.5–7(–8) × 4.5–6(–6.5) | 26.4–64.0 × 5.6–10.4 | On the ground in frondose woods; Ectomycorrhizal, humicolous, scattered to gregarious in the mixed forest of Populus nigra L., Salix alba L. and Hippophae rhamnoides L. | China, India, Japan, North America, USA | [3,12,56,60,73] | ||
| T. wakefieldiae | 40–65 × 7.5–12 | 40–65 × 7.5–12 | 4 | Colonizes all kinds of wood debris | Denmark, Estonia, France, Germany, North temperate, Norway, Spain, Russia, Sweden, UK, USA | [14] | |
| T. wuliangshanensis | (5–)5.2–8.7(–9.3) × (3.7–)4.5–7.2(–7.6) | 30–60 × 5–9.5 | 4 | 28–55 × 3–7.5 | On the ground of pine-broadleaved mixed forest | China | Present study |
| T. zeylanica | 8–12 × 4–8.5 | On the ground | Ceylon | [3] | |||
| Species | Edible Value | Medicinal Value | Medicinal Efficacy | Reference |
|---|---|---|---|---|
| Thelephora anthocephala | P | Anticancer | [80] | |
| T. aurantiotincta | Y | Y | Anticancer | [73,80,81,82,83,84,85] |
| T. austrosinensis | Y | [5] | ||
| T. caespitulans | P | Anticancer | [82] | |
| T. caryophyllea | P | Anticancer | [82] | |
| T. fuscella | Y | [81] | ||
| T. ganbajun | Y | Y | Treat leukemia, anticancer and boost immunity | [5,83,85,86,87,88] |
| T. intybacea | P | Anticancer | [80] | |
| T. japonica | Y | P | Anti-microbial activities | [20,81,88,89] |
| T. palmata | Y | P | Anticancer | [81,82] |
| T. penicillata | P | Anticancer | [80] | |
| T. regularis | P | Anticancer | [80] | |
| T. scissilis | P | Anticancer | [80] | |
| T. sikkimensis | P | [26] | ||
| T. terrestris | Y | Anticancer | [90] | |
| T. vialis | Y | Y | Anticancer and anti-allergic agent | [73,91,92] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, X.-F.; Tibpromma, S.; Xu, J.-C.; Kumla, J.; Karunarathna, S.C.; Zhao, C.-L. Taxonomy and Phylogeny Reveal Two New Potential Edible Ectomycorrhizal Mushrooms of Thelephora from East Asia. Diversity 2021, 13, 646. https://doi.org/10.3390/d13120646
Liu X-F, Tibpromma S, Xu J-C, Kumla J, Karunarathna SC, Zhao C-L. Taxonomy and Phylogeny Reveal Two New Potential Edible Ectomycorrhizal Mushrooms of Thelephora from East Asia. Diversity. 2021; 13(12):646. https://doi.org/10.3390/d13120646
Chicago/Turabian StyleLiu, Xiang-Fu, Saowaluck Tibpromma, Jian-Chu Xu, Jaturong Kumla, Samantha Chandranath Karunarathna, and Chang-Lin Zhao. 2021. "Taxonomy and Phylogeny Reveal Two New Potential Edible Ectomycorrhizal Mushrooms of Thelephora from East Asia" Diversity 13, no. 12: 646. https://doi.org/10.3390/d13120646