
Invasion at the Edge: The Case of Rosa rugosa (Rosaceae) in Italy


 ,
,  , , and
, , and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Survey Method for the Literature Review
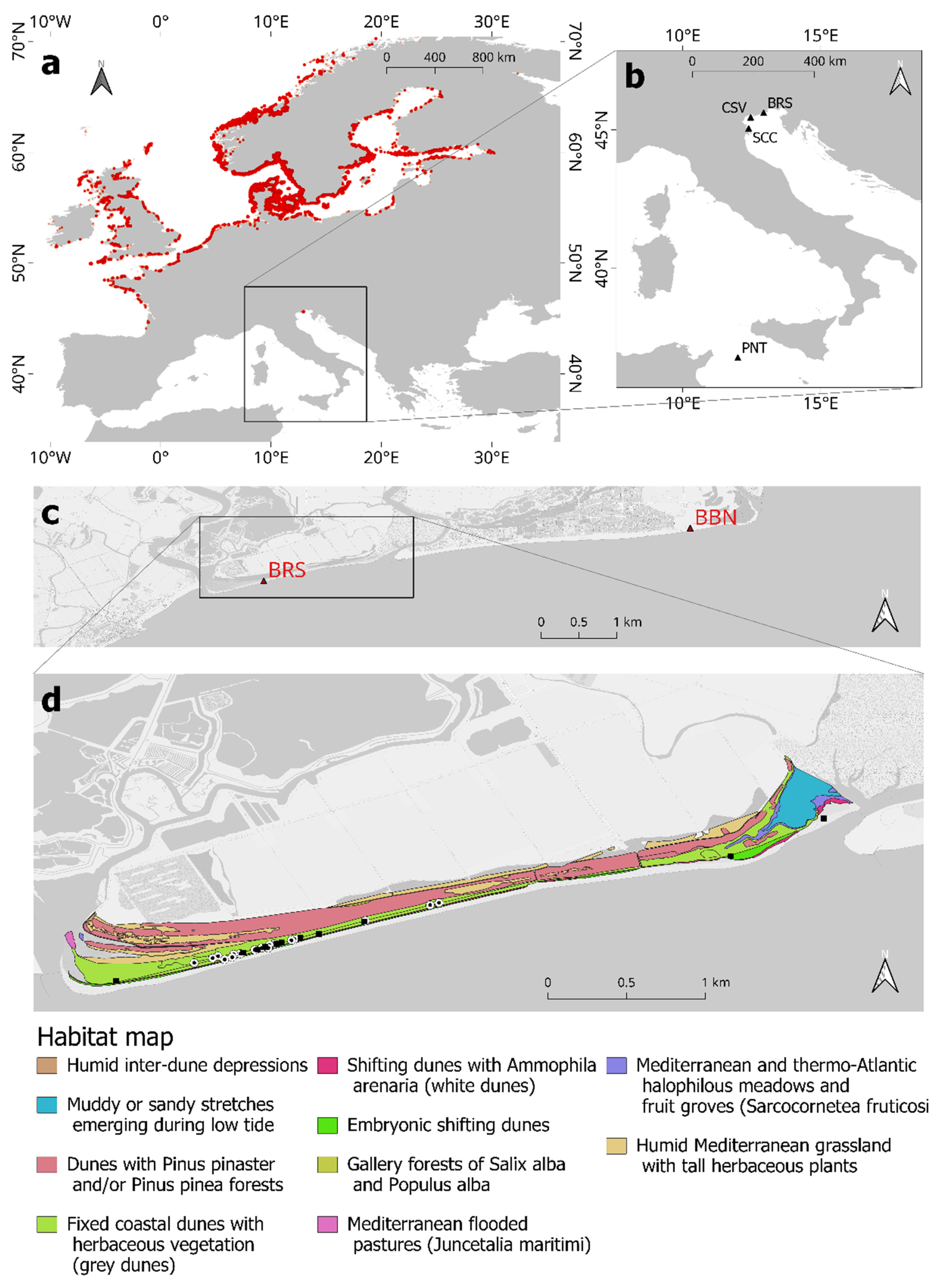
2.2. Collection of New Data on R. rugosa in Italy: Study Area
2.3. Data Collection and Sampling Design
2.4. Data Analysis
3. Results
3.1. From the Far East to West: The Establishment of Rosa rugosa in Europe
3.2. Functional Characteristics of Rosa rugosa in Invaded and Native Ranges
3.3. Genetic Characterisation of R. rugosa
3.3.1. The Genome of R. rugosa
3.3.2. Genetic Diversity of R. rugosa from Native to Invaded Areas
3.4. Mycorrhizal Association in R. rugosa
3.5. Impacts on Ecosystem Services
3.5.1. Habitat and Native Resident Communities
3.5.2. Commercial Implication in Relation to Cultural Ecosystem Services
3.5.3. Human Health
3.6. Management and Control
3.6.1. Prevention
3.6.2. Eradication
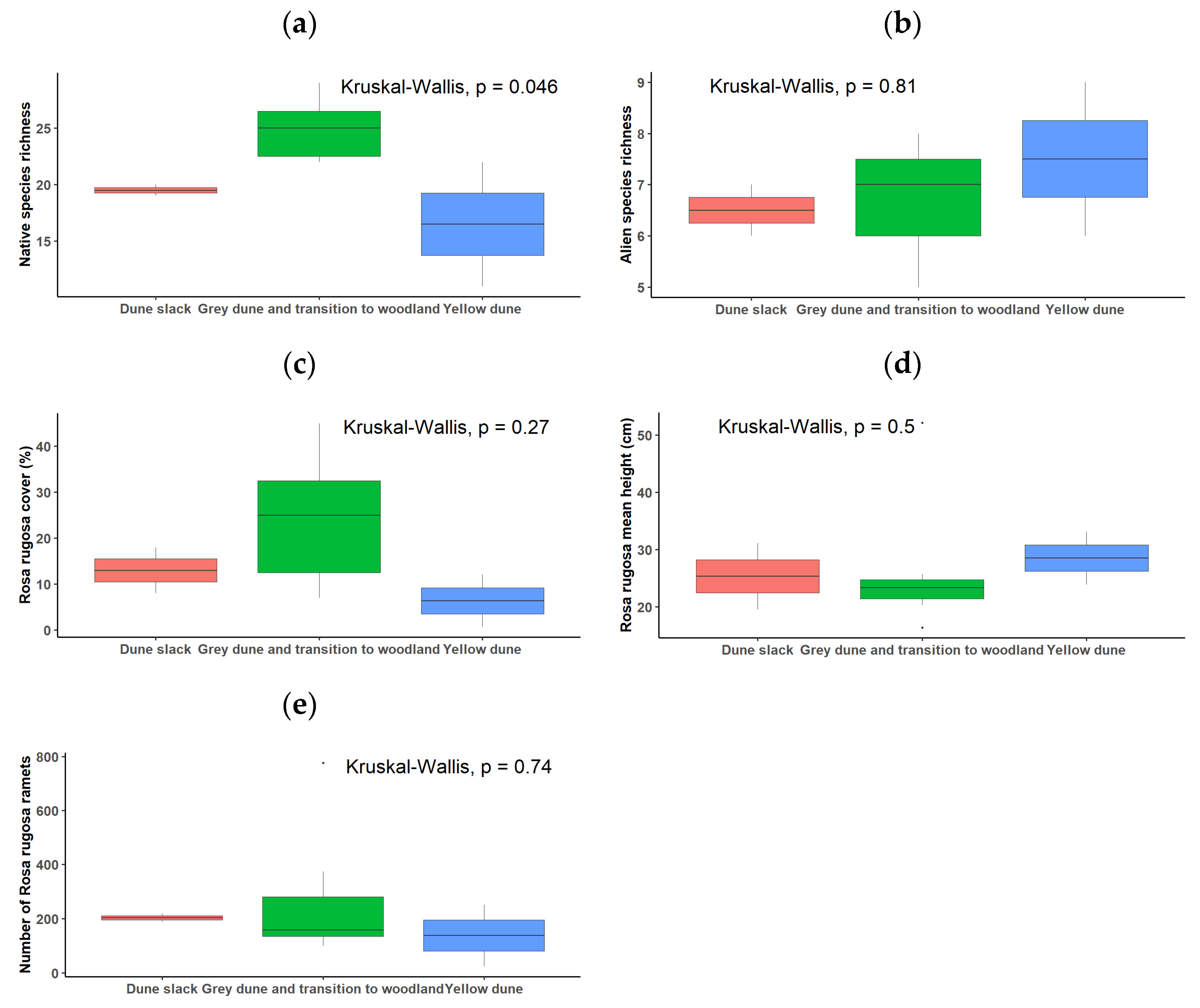
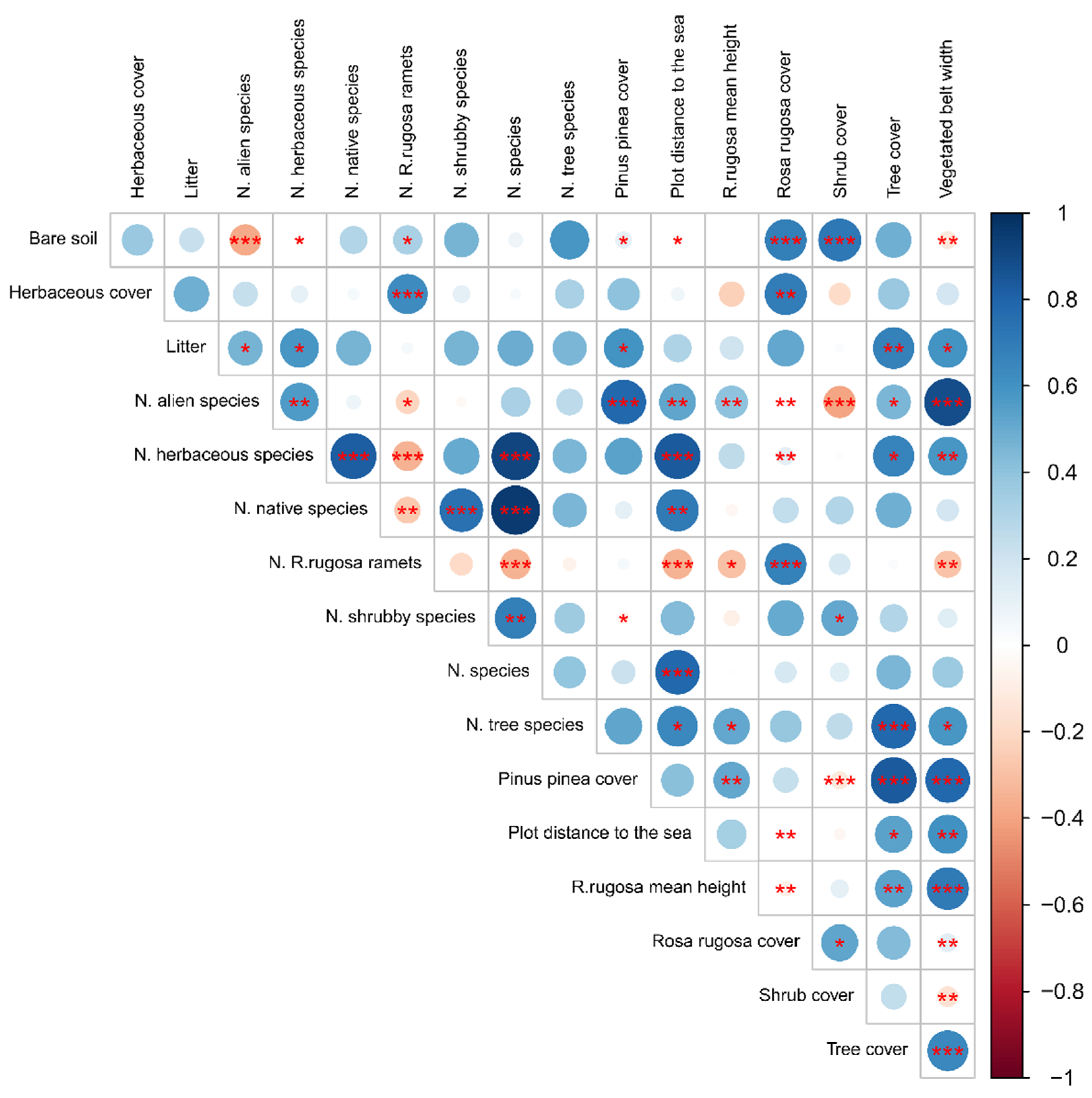
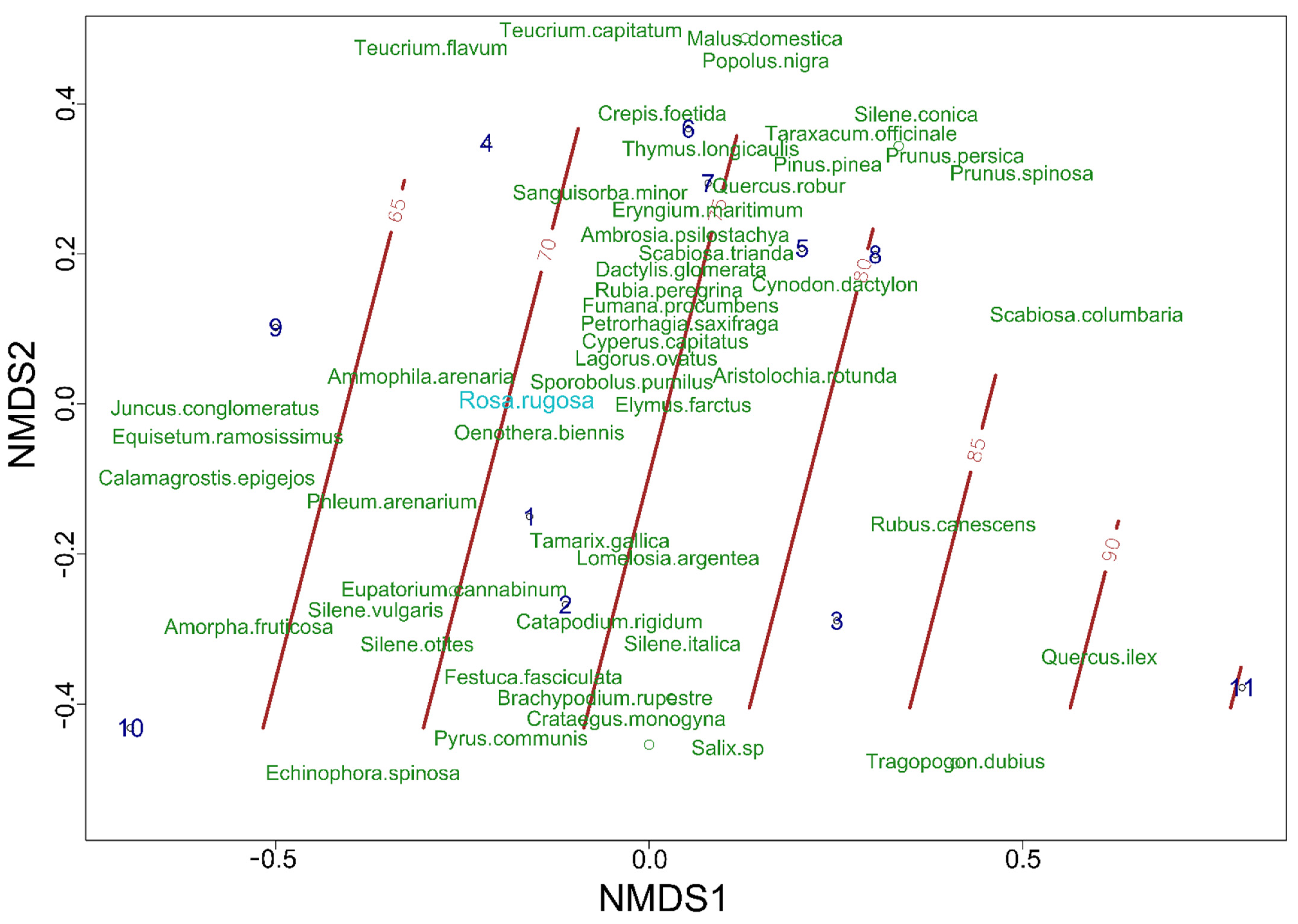
3.7. Rosa rugosa Establishment in Italy: Quantitative Characterisation of Its Populations on North-Eastern Italian Coasts
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Simberloff, D.; Martin, J.-L.; Genovesi, P.; Maris, V.; Wardle, D.A.; Aronson, J.; Courchamp, F.; Galil, B.; García-Berthou, E.; Pascal, M.; et al. Impacts of Biological Invasions: What’s What and the Way Forward. Trends Ecol. Evol. 2013, 28, 58–66. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodríguez-Labajos, B.; Binimelis, R.; Monterroso, I. Multi-Level Driving Forces of Biological Invasions. Ecol. Econ. 2009, 69, 63–75. [Google Scholar] [CrossRef]
- Early, R.; Bradley, B.A.; Dukes, J.S.; Lawler, J.J.; Olden, J.D.; Blumenthal, D.M.; Gonzalez, P.; Grosholz, E.D.; Ibañez, I.; Miller, L.P.; et al. Global Threats from Invasive Alien Species in the Twenty-First Century and National Response Capacities. Nat. Commun. 2016, 7, 12485. [Google Scholar] [CrossRef] [PubMed]
- Robinson, T.B.; Martin, N.; Loureiro, T.G.; Matikinca, P.; Robertson, M.P. Double Trouble: The Implications of Climate Change for Biological Invasions. NeoBiota 2020, 62, 463–487. [Google Scholar] [CrossRef]
- Vilà, M.; Espinar, J.L.; Hejda, M.; Hulme, P.E.; Jarošík, V.; Maron, J.L.; Pergl, J.; Schaffner, U.; Sun, Y.; Pyšek, P. Ecological Impacts of Invasive Alien Plants: A Meta-Analysis of Their Effects on Species, Communities and Ecosystems: Ecological Impacts of Invasive Alien Plants. Ecol. Lett. 2011, 14, 702–708. [Google Scholar] [CrossRef]
- Liao, C.; Peng, R.; Luo, Y.; Zhou, X.; Wu, X.; Fang, C.; Chen, J.; Li, B. Altered Ecosystem Carbon and Nitrogen Cycles by Plant Invasion: A Meta-analysis. New Phytol. 2008, 177, 706–714. [Google Scholar] [CrossRef] [PubMed]
- Morris, T.L.; Barger, N.N.; Cramer, M.D. Ecophysiological Traits of Invasive Alien Acacia Cyclops Compared to Co-Occuring Native Species in Strandveld Vegetation of the Cape Floristic Region: Ecophysiological Traits of Invasive Acacia. Austral Ecol. 2020, 45, 48–59. [Google Scholar] [CrossRef]
- Siniscalco, C.; Barni, E.; Bacaro, G. Non-native species distribution along the elevation gradient in the western Italian Alps. Plant Biosyst. 2011, 145, 150–158. [Google Scholar] [CrossRef]
- Hobbie, S.E. Effects of Plant Species on Nutrient Cycling. Trends Ecol. Evol. 1992, 7, 336–339. [Google Scholar] [CrossRef]
- van Breemen, N. Plant-soil interactions: Ecological aspects and evolutionary implications. Biogeochemistry 1998, 42, 1–4. [Google Scholar] [CrossRef]
- Ehrenfeld, J.G.; Scott, N. Invasive Species and the Soil: Effects on Organisms and Ecosystem Processes 1. Ecol. Appl. 2001, 11, 1259–1260. [Google Scholar] [CrossRef]
- Tordoni, E.; Bacaro, G.; Weigelt, P.; Cameletti, M.; Janssen, J.A.; Acosta, A.T.; Bagella, S.; Filigheddu, R.; Bergmeier, E.; Buckley, H.L.; et al. Disentangling native and alien plant diversity in coastal sand dune ecosystems worldwide. J. Veg. Sci. 2021, 32, e12861. [Google Scholar] [CrossRef]
- Ciccarelli, D. Mediterranean Coastal Dune Vegetation: Are Disturbance and Stress the Key Selective Forces That Drive the Psammophilous Succession? Estuar. Coast. Shelf Sci. 2015, 165, 247–253. [Google Scholar] [CrossRef]
- Tordoni, E.; Petruzzellis, F.; Nardini, A.; Savi, T.; Bacaro, G. Make It Simpler: Alien Species Decrease Functional Diversity of Coastal Plant Communities. J. Veg. Sci. 2019, 30, 498–509. [Google Scholar] [CrossRef]
- Tordoni, E.; Petruzzellis, F.; Nardini, A.; Bacaro, G. Functional Divergence Drives Invasibility of Plant Communities at the Edges of a Resource Availability Gradient. Diversity 2020, 12, 148. [Google Scholar] [CrossRef] [Green Version]
- Martínez, M.L.; Psuty, N.P. (Eds.) Coastal Dunes; Ecological Studies; Springer: Berlin/Heidelberg, Germany, 2004; Volume 171, ISBN 978-3-540-74001-8. [Google Scholar]
- Bajocco, S.; De Angelis, A.; Perini, L.; Ferrara, A.; Salvati, L. The Impact of Land Use/Land Cover Changes on Land Degradation Dynamics: A Mediterranean Case Study. Environ. Manag. 2012, 49, 980–989. [Google Scholar] [CrossRef] [PubMed]
- Malavasi, M.; Santoro, R.; Cutini, M.; Acosta, A.T.R.; Carranza, M.L. What Has Happened to Coastal Dunes in the Last Half Century? A Multitemporal Coastal Landscape Analysis in Central Italy. Landsc. Urban Plan. 2013, 119, 54–63. [Google Scholar] [CrossRef]
- Marignani, M.; Bruschi, D.; Astiaso Garcia, D.; Frondoni, R.; Carli, E.; Pinna, M.S.; Cumo, F.; Gugliermetti, F.; Saatkamp, A.; Doxa, A.; et al. Identification and Prioritisation of Areas with High Environmental Risk in Mediterranean Coastal Areas: A Flexible Approach. Sci. Total Environ. 2017, 590–591, 566–578. [Google Scholar] [CrossRef]
- Maccherini, S.; Bacaro, G.; Tordoni, E.; Bertacchi, A.; Castagnini, P.; Foggi, B.; Gennai, M.; Mugnai, M.; Sarmati, S.; Angiolini, C. Enough Is Enough? Searching for the Optimal Sample Size to Monitor European Habitats: A Case Study from Coastal Sand Dunes. Diversity 2020, 12, 138. [Google Scholar] [CrossRef] [Green Version]
- Marcantonio, M.; Rocchini, D.; Ottaviani, G. Impact of Alien Species on Dune Systems: A Multifaceted Approach. Biodivers. Conserv. 2014, 23, 2645–2668. [Google Scholar] [CrossRef]
- Gallien, L.; Carboni, M. The Community Ecology of Invasive Species: Where Are We and What’s Next? Ecography 2017, 40, 335–352. [Google Scholar] [CrossRef] [Green Version]
- Tordoni, E.; Napolitano, R.; Maccherini, S.; Da Re, D.; Bacaro, G. Ecological Drivers of Plant Diversity Patterns in Remnants Coastal Sand Dune Ecosystems along the Northern Adriatic Coastline. Ecol. Res. 2018, 33, 1157–1168. [Google Scholar] [CrossRef]
- Giulio, S.; Acosta, A.T.R.; Carboni, M.; Campos, J.A.; Chytrý, M.; Loidi, J.; Pergl, J.; Pyšek, P.; Isermann, M.; Janssen, J.A.M.; et al. Alien Flora across European Coastal Dunes. Appl. Veg. Sci. 2020, 23, 317–327. [Google Scholar] [CrossRef]
- Isermann, M. Classification and Habitat Characteristics of Plant Communities Invaded by the Non-Native Rosa rugosa Thunb. in NW Europe. Phytocoenologia 2008, 38, 133–150. [Google Scholar] [CrossRef]
- Kelager, A.; Pedersen, J.S.; Bruun, H.H. Multiple Introductions and No Loss of Genetic Diversity: Invasion History of Japanese Rose, Rosa rugosa, in Europe. Biol. Invasions 2013, 15, 1125–1141. [Google Scholar] [CrossRef]
- Kottek, M.; Grieser, J.; Beck, C.; Rudolf, B.; Rubel, F. World Map of the Köppen-Geiger Climate Classification Updated. Meteorol. Z. 2006, 15, 259–263. [Google Scholar] [CrossRef]
- Pignatti, S. Flora d’Italia; Edagricole: Bologna, Italy, 1982. [Google Scholar]
- Pignatti, S.; Guarino, R.; La Rosa, M. Flora d’Italia, 2nd ed.; Edagricole di New Business Media: Bologna, Italy, 2017–2019.
- Celesti-Grapow, L.; Alessandrini, A.; Arrigoni, P.V.; Banfi, E.; Bernardo, L.; Bovio, M.; Brundu, G.; Cagiotti, M.R.; Camarda, I.; Carli, E.; et al. Inventory of the Non-native Flora of Italy. Plant Biosyst. 2009, 143, 386–430. [Google Scholar] [CrossRef]
- Galasso, G.; Conti, F.; Peruzzi, L.; Ardenghi, N.M.G.; Banfi, E.; Celesti-Grapow, L.; Albano, A.; Alessandrini, A.; Bacchetta, G.; Ballelli, S.; et al. An Updated Checklist of the Vascular Flora Alien to Italy. Plant Biosyst. 2018, 152, 556–592. [Google Scholar] [CrossRef]
- Wei, T.; Simko, V. R Package ‘Corrplot’: Visualization of a Correlation Matrix (Version 0.90). 2021. Available online: https://github.com/taiyun/corrplot (accessed on 1 September 2021).
- RStudio Team. RStudio: Integrated Development for R; RStudio PBC: Boston, MA, USA, 2020; Available online: http://www.rstudio.com/ (accessed on 1 September 2021).
- McCune, B.; Grace, J.B. Analysis of Ecological Communities; MjM Software: Gleneden Beach, OR, USA, 2002. [Google Scholar]
- Oksanen, F.J.; Blanchet, G.; Friendly, M.; Kindt, R.; Legendre, P.; McGlinn, D.; Minchin, P.R.; O’Hara, R.B.; Simpson, G.L.; Solymos, P.; et al. R Package Version 2.5-7 ‘Vegan’. 2020. Available online: https://CRAN.R-project.org/package=vegan (accessed on 1 September 2021).
- Weidema, I. NOBANIS—Invasive Alien Species Fact Sheet—Rosa rugosa. From: Online Database of the European Network on Invasive Alien Species—NOBANIS. 2006. Available online: www.nobanis.org (accessed on 1 September 2021).
- Bruun, H.H. Rosa rugosa Thunb. ex Murray. J. Ecol. 2005, 93, 441–470. [Google Scholar] [CrossRef]
- Nakata, Y.; Hioki, Y.; Oguchi, T. Evaluation of Beach Vegetation for Conservation/Rehabilitation in Tottori Prefecture, Southwest Japan. Landsc. Ecol. Eng. 2018, 14, 231–243. [Google Scholar] [CrossRef]
- Landcare. Rosa rugosa. Landcare Plants Database: Auckland, New Zealand: Landcare Research. Available online: https://nzflora.landcareresearch.co.nz/ (accessed on 1 September 2021).
- Garbary, D.; Hill, N. Invasion of Rosa rugosa (Rugosa Rose) into Coastal Plant Communities of Brier Island, Nova Scotia. Can. Field-Nat. 2013, 127, 319–331. [Google Scholar] [CrossRef] [Green Version]
- CABI. Rosa rugosa. Invasive Species Compendium; CAB International: Wallingford, UK; Available online: www.cabi.org/isc (accessed on 1 September 2021).
- GBIF. Global Biodiversity Information Facility. Available online: www.gbif.org (accessed on 1 September 2021).
- Vladimirov, V.; Petrova, A.; Stoyanov, S.; Bancheva, S.; Delcheva, M. Rosa rugosa (Rosaceae): An Alien Species in the Bulgarian Flora. Phytol. Balc. 2018, 24, 337–341. [Google Scholar]
- Lægaard, S. Forurening af Floraen. Flora Fauna 1987, 93, 87–143. [Google Scholar]
- Kowarik, I. Biologische Invasionen: Neophyten und Neozoenin Mitteleuropa; Ulmer: Stuttgart, Germany, 2003. (In German) [Google Scholar]
- Svart, H.E.; Lyck, G. Introducerede Planter—Forvildede og Adventive Arter, 2nd ed.; Inst. for Økologisk Botanik, Københavns Universitet, Skov-og Naturstyrelsen: København, Denmark, 1991. [Google Scholar]
- Nilsson, O. Wild roses in Norden: Taxonomic discussion. Acta Bot. Fenn. 1999, 162, 169–173. [Google Scholar]
- Isermann, M. Expansion of Rosa rugosa and Hippophaë Rhamnoides in Coastal Grey Dunes: Effects at Different Spatial Scales. Flora Morphol. Distrib. Funct. Ecol. Plants 2008, 203, 273–280. [Google Scholar] [CrossRef]
- MacDougall, A.S.; Gilbert, B.; Levine, J.M. Plant Invasions and the Niche. J. Ecol. 2009, 97, 609–615. [Google Scholar] [CrossRef]
- Zhang, S.; Isermann, M.; Gan, W.; Breed, M. Invasive Rosa rugosa Populations Outperform Native Populations, but Some Populations Have Greater Invasive Potential than Others. Sci. Rep. 2018, 8, 5735. [Google Scholar] [CrossRef] [Green Version]
- Helsen, K.; Van Cleemput, E.; Bassi, L.; Somers, B.; Honnay, O. Optical Traits Perform Equally Well as Directly-measured Functional Traits in Explaining the Impact of an Invasive Plant on Litter Decomposition. J. Ecol. 2020, 108, 2000–2011. [Google Scholar] [CrossRef]
- Helsen, K.; Matsushima, H.; Somers, B.; Honnay, O. A Trait-Based Approach across the Native and Invaded Range to Understand Plant Invasiveness and Community Impact. Oikos 2021, 130, 1001–1013. [Google Scholar] [CrossRef]
- van Kleunen, M.; Weber, E.; Fischer, M. A Meta-Analysis of Trait Differences between Invasive and Non-Invasive Plant Species. Ecol. Lett. 2010, 13, 235–245. [Google Scholar] [CrossRef] [Green Version]
- Funk, J.L.; Vitousek, P.M. Resource-Use Efficiency and Plant Invasion in Low-Resource Systems. Nature 2007, 446, 1079–1081. [Google Scholar] [CrossRef] [PubMed]
- Chen, F.; Su, L.; Hu, S.; Xue, J.-Y.; Liu, H.; Liu, G.; Jiang, Y.; Du, J.; Qiao, Y.; Fan, Y.; et al. A Chromosome-Level Genome Assembly of Rugged Rose (Rosa rugosa) Provides Insights into Its Evolution, Ecology, and Floral Characteristics. Hortic. Res. 2021, 8, 141. [Google Scholar] [CrossRef] [PubMed]
- Jiang, H.; He, J.; Meng, J. Characterization of the Complete Plastid Genome of a Chinese Endangered Species Rosa rugosa Thunb. Mitochondrial DNA Part B 2019, 4, 1679–1680. [Google Scholar] [CrossRef]
- Kim, Y.; Heo, K.-I.; Nam, S.; Xi, H.; Lee, S.; Park, J. The Complete Chloroplast Genome of Candidate New Species from Rosa rugosa in Korea (Rosaceae). Mitochondrial DNA Part B 2019, 4, 2433–2435. [Google Scholar] [CrossRef] [Green Version]
- Park, J.; Xi, H.; Kim, Y.; Nam, S.; Heo, K.-I. The Complete Mitochondrial Genome of New Species Candidate of Rosa rugosa (Rosaceae). Mitochondrial DNA Part B 2020, 5, 3435–3437. [Google Scholar] [CrossRef]
- Hibrand Saint-Oyant, L.; Ruttink, T.; Hamama, L.; Kirov, I.; Lakhwani, D.; Zhou, N.N.; Bourke, P.M.; Daccord, N.; Leus, L.; Schulz, D.; et al. A High-Quality Genome Sequence of Rosa Chinensis to Elucidate Ornamental Traits. Nat. Plants 2018, 4, 473–484. [Google Scholar] [CrossRef] [Green Version]
- Guo, J.; Wang, Q.; Liu, L.; Ren, S.; Li, S.; Liao, P.; Zhao, Z.; Lu, C.; Jiang, B.; Sunkar, R.; et al. Analysis of MicroRNAs, Phased Small Interfering RNAs and Their Potential Targets in Rosarugosa Thunb. BMC Genom. 2019, 19, 983. [Google Scholar] [CrossRef]
- Fe’r, T.; Vasa’k, P.; Vojta, J.; Marhold, K. Out of the Alps or Carpathians? Origin of Central European populations of Rosa pendulina. Preslia 2007, 79, 367–376. [Google Scholar]
- Wang, L.-Y.; Ikeda, H.; Liu, T.-L.; Wang, Y.-J.; Liu, J.-Q. Repeated Range Expansion and Glacial Endurance of Potentilla Glabra (Rosaceae) in the Qinghai-Tibetan Plateau. J. Integr. Plant Biol. 2009, 51, 698–706. [Google Scholar] [CrossRef]
- Bossdorf, O.; Auge, H.; Lafuma, L.; Rogers, W.E.; Siemann, E.; Prati, D. Phenotypic and Genetic Differentiation between Native and Introduced Plant Populations. Oecologia 2005, 144, 1–11. [Google Scholar] [CrossRef]
- Wilson, J.R.U.; Dormontt, E.E.; Prentis, P.J.; Lowe, A.J.; Richardson, D.M. Something in the Way You Move: Dispersal Pathways Affect Invasion Success. Trends Ecol. Evol. 2009, 24, 136–144. [Google Scholar] [CrossRef]
- van der Heijden, M.G.A.; Bardgett, R.D.; van Straalen, N.M. The Unseen Majority: Soil Microbes as Drivers of Plant Diversity and Productivity in Terrestrial Ecosystems. Ecol. Lett. 2008, 11, 296–310. [Google Scholar] [CrossRef]
- Smith, S.E.; Read, D.J. Mycorrhizal Symbiosis; Academic Press: London, UK, 1997. [Google Scholar]
- Santolamazza-Carbone, S.; Durán-Otero, M.; Calviño-Cancela, M. Context Dependency, Co-Introductions, Novel Mutualisms, and Host Shifts Shaped the Ectomycorrhizal Fungal Communities of the Alien Tree Eucalyptus Globulus. Sci. Rep. 2019, 9, 7121. [Google Scholar] [CrossRef] [Green Version]
- Molina, R.; Massicotte, H.; Trappe, J.M. Specificity phenomena in mycorrhizal symbioses: Community ecological consequencesand practical implications. In Mycorrhizal Functioning: An Integrative Plant-Fungal Process; Allen, M.F., Ed.; Chapman and Hall: New York, NY, USA, 1992; pp. 357–423. [Google Scholar]
- Moyano, J.; Dickie, I.A.; Rodriguez-Cabal, M.A.; Nuñez, M.A. Patterns of Plant Naturalization Show That Facultative Mycorrhizal Plants Are More Likely to Succeed Outside Their Native Eurasian Ranges. Ecography 2020, 43, 648–659. [Google Scholar] [CrossRef] [Green Version]
- van Kleunen, M.; Pyšek, P.; Dawson, W.; Essl, F.; Kreft, H.; Pergl, J.; Weigelt, P.; Stein, A.; Dullinger, S.; König, C.; et al. The Global Naturalized Alien Flora (GloNAF) database. Ecology 2019, 100, e02542. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gemma, J.N.; Koske, R.E. Arbuscular Mycorrhizae in Sand Dune Plants of the North Atlantic Coast of the U.S.: Field and Greenhouse Inoculation and Presence of Mycorrhizae in Planting Stock. J. Environ. Manag. 1997, 50, 251–264. [Google Scholar] [CrossRef] [Green Version]
- Menkis, A.; Ihrmark, K.; Stenlid, J.; Vasaitis, R. Root-Associated Fungi of Rosa rugosa Grown on the Frontal Dunes of the Baltic Sea Coast in Lithuania. Microb. Ecol. 2014, 67, 769–774. [Google Scholar] [CrossRef]
- Blaszkowski, J.; Madej, T.; Tadych, M. Entrophospora balticasp. nov. and Glomus fuegianum, two species in the glomales from Poland. Mycotaxon 1998, 68, 165–184. [Google Scholar]
- Rodriguez, R.; Redman, R. More than 400 million years of evolution and some plants still can’t make it on their own: Plant stress tolerance via fungal symbiosis. J. Exp. Bot. 2008, 59, 1109–1114. [Google Scholar] [CrossRef]
- Isermann, M. Effects of Rosa rugosa invasion in different coastal dune vegetation types. In Plant Invasions: Human Perception, Ecological Impacts and Management; Tokarska-Guzik, B., Brock, J.H., Brundu, G., Child, L., Daehler, C.C., Pyšek, P., Eds.; Backhuys Publishers: Leiden, The Netherlands, 2008; pp. 289–306. [Google Scholar]
- Lanta, V.; Hyvönen, T.; Norrdahl, K. Non-Native and Native Shrubs Have Differing Impacts on Species Diversity and Composition of Associated Plant Communities. Plant Ecol. 2013, 214, 1517–1528. [Google Scholar] [CrossRef]
- Stefanowicz, A.M.; Zubek, S.; Stanek, M.; Grześ, I.M.; Rożej-Pabijan, E.; Błaszkowski, J.; Woch, M.W. Invasion of Rosa rugosa Induced Changes in Soil Nutrients and Microbial Communities of Coastal Sand Dunes. Sci. Total Environ. 2019, 677, 340–349. [Google Scholar] [CrossRef]
- Dassonville, N.; Vanderhoeven, S.; Vanparys, V.; Hayez, M.; Gruber, W.; Meerts, P. Impacts of Alien Invasive Plants on Soil Nutrients Are Correlated with Initial Site Conditions in NW Europe. Oecologia 2008, 157, 131–140. [Google Scholar] [CrossRef]
- Elleriis, P.; Pedersen, M.L.; Toft, S. Impact of Invasive Rosa rugosa on the Arthropod Fauna of Danish Yellow Dunes. Biol. Invasions 2015, 17, 3289–3302. [Google Scholar] [CrossRef] [Green Version]
- Toft, S. Habitat Specialist Spiders in Coastal Dunes Benefit from Eradication of the Invasive Shrub Rosa rugosa. J. Insect Conserv. 2020, 24, 993–1003. [Google Scholar] [CrossRef]
- Grześ, I.M.; Rożej-Pabijan, E.; Stefanowicz, A.M.; Woch, M.W. Community Composition of Ants beneath Invasive Plant Rosa rugosa. Entomol. Sci. 2021, 24, 361–365. [Google Scholar] [CrossRef]
- Lu, J.; Wang, C. Medicinal Components and Pharmacological Effects of Rosa rugosa. Rec. Nat. Prod. 2018, 12, 535–543. [Google Scholar] [CrossRef]
- Miljøministeriet, Skov og Naturstyrelsen; Danmarks Naturfredningsforening; Friluftsrådet. Beskyt den Vilde Flora Langs Kysterne. 2004. Available online: https://naturstyrelsen.dk/media/243017/beskyt-den-vilde-flora-langs-kysterne-2008.pdf (accessed on 1 September 2021).
- Bruun, H.H. Prospects for Biocontrol of Invasive Rosa rugosa. Biocontrol 2006, 51, 141–181. [Google Scholar] [CrossRef]
- Simberloff, D. Risks of Biological Control for Conservation Purposes. BioControl 2012, 57, 263–276. [Google Scholar] [CrossRef]
- Portal to the Flora of Italy. Available online: http:/dryades.units.it/floritaly (accessed on 1 September 2021).
- Celesti-Grapow, L.; Pretto, F.; Carli, E.; Blasi, C. Flora Vascolare Alloctona e Invasiva Delle Regioni d’Italia; Casa Editrice Università La Sapienza: Roma, Italy, 2010. [Google Scholar]
- Banfi, E.; Galasso, G. La flora spontanea della città di Milano alle soglie del terzo millennio e i suoi cambiamenti a partire dal 1700. In Memorie Della Società Italiana di Scienze Naturali e del Museo Civico di Storia Naturale di Milano; G. Bernardoni: Milano, Italy, 1998; Volume 28. [Google Scholar]
- Wikiplantbase #Italia. Available online: http://bot.biologia.unipi.it/wpb/italia/search_segn.html?stype=floritaly&id=8600&taxon=Rosa%20rugosa%20Thunb (accessed on 1 September 2021).
- Masin, R.; Scortegagna, S. Flora alloctona del Veneto centromeridionale (province di Padova, Rovigo, Venezia e Vicenza—Veneto—NE Italia). Nat. Vicentina 2012, 15, 5–54. [Google Scholar]
- Di Gregorio, B.; Bellone, G.; Cassanego, E.; Longo, D.; Montalto, S.; Montoleone, E.; Tomasello, G. Rassegna di segnalazioni notevoli riguardanti la Sicilia comparse nel forum Acta Plantarum. Acta Plant. Notes 2017, 5, 70–74. [Google Scholar]
- Bartolucci, F.; Galasso, G.; Peruzzi, L.; Conti, F. Report 2020 on Plant Biodiversity in Italy: Native and Alien Vascular Flora. Nat. Hist. Sci. 2021, 8, 41–54. [Google Scholar] [CrossRef]
- Acosta, A.; Carranza, M.L.; Izzi, C.F. Are There Habitats That Contribute Best to Plant Species Diversity in Coastal Dunes? Biodivers. Conserv. 2009, 18, 1087–1098. [Google Scholar] [CrossRef]
- Kollmann, J.; Frederiksen, L.; Vestergaard, P.; Bruun, H.H. Limiting Factors for Seedling Emergence and Establishment of the Invasive Non-Native Rosa rugosa in a Coastal Dune System. Biol. Invasions 2006, 9, 31–42. [Google Scholar] [CrossRef]
- Jørgensen, R.H.; Kollmann, J. Invasion of Coastal Dunes by the Alien Shrub Rosa rugosa Is Associated with Roads, Tracks and Houses. Flora Morphol. Distrib. Funct. Ecol. Plants 2009, 204, 289–297. [Google Scholar] [CrossRef]
- Khapugin, A.A.; Sołtys-Lelek, A.; Fedoronchuk, N.M.; Muldashev, A.A.; Agafonov, V.A.; Kazmina, E.S.; Vasjukov, V.M.; Baranova, O.G.; Buzunova, I.O.; Teteryuk, L.V.; et al. Taxon-level assessment of the data collection quality in Atlas Florae Europaeae: Insights from the case of Rosa (Rosaceae) in Eastern Europe. Nord. J. Bot. 2021, 39. [Google Scholar] [CrossRef]
- Rocchini, D.; Garzon-Lopez, C.X.; Marcantonio, M.; Amici, V.; Bacaro, G.; Bastin, L.; Brummitt, N.; Chiarucci, A.; Foody, G.M.; Hauffe, H.C.; et al. Anticipating species distributions: Handling sampling effort bias under a Bayesian framework. Sci. Total Environ. 2017, 584, 282–290. [Google Scholar] [CrossRef] [PubMed] [Green Version]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ribotta, S.; Liccari, F.; Muggia, L.; Pallavicini, A.; Bagnolini, F.; Tordoni, E.; Bacaro, G. Invasion at the Edge: The Case of Rosa rugosa (Rosaceae) in Italy. Diversity 2021, 13, 645. https://doi.org/10.3390/d13120645
Ribotta S, Liccari F, Muggia L, Pallavicini A, Bagnolini F, Tordoni E, Bacaro G. Invasion at the Edge: The Case of Rosa rugosa (Rosaceae) in Italy. Diversity. 2021; 13(12):645. https://doi.org/10.3390/d13120645
Chicago/Turabian StyleRibotta, Sara, Francesco Liccari, Lucia Muggia, Alberto Pallavicini, Francesco Bagnolini, Enrico Tordoni, and Giovanni Bacaro. 2021. "Invasion at the Edge: The Case of Rosa rugosa (Rosaceae) in Italy" Diversity 13, no. 12: 645. https://doi.org/10.3390/d13120645