
Coral Communities on Marginal High-Latitude Reefs in West Australian Marine Parks

 ,
,  ,
,
Abstract
:1. Introduction
- (1)
- Estimate abundance of hard coral in terms of % cover for five marine parks in the south-west of WA and, where possible, identify temporal trends in total coral coverage between 1999 and 2019;
- (2)
- Where trends in coral cover estimates are detected, delineate sites that drive these trends within each marine park; and
- (3)
- Where coral is sufficiently abundant, determine whether the composition of different hard-coral genera vary inter-annually and whether there is a sequential change in coral genera through time, in each marine park.
2. Materials and Methods
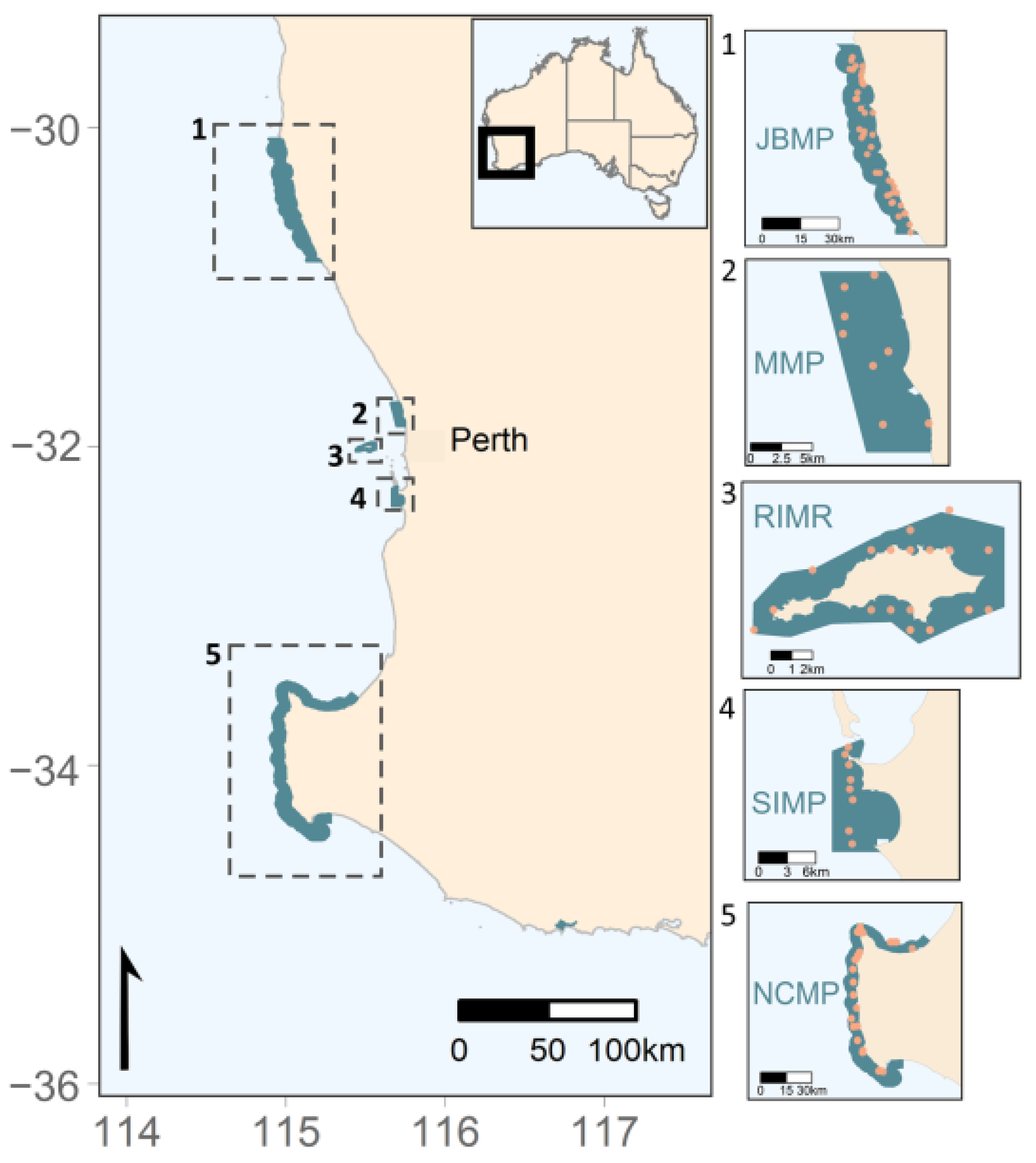
2.1. Data Collection and Collation
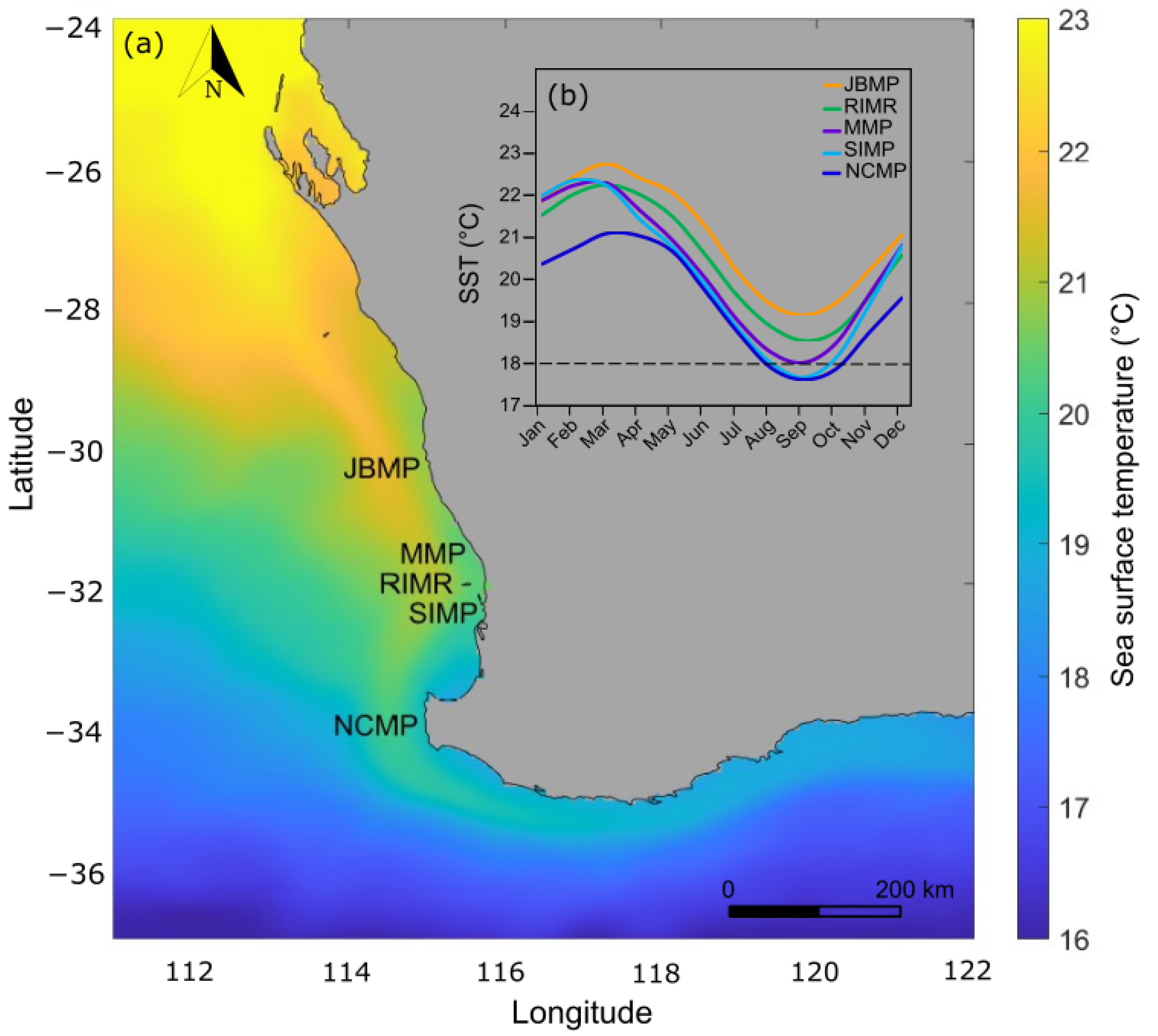
2.2. Temperature Profiles
2.2.1. JBMP
2.2.2. MMP and SIMP
2.2.3. RIMR
2.2.4. NCMP
2.3. Analysis of Total Hard-Coral Coverage
2.4. Analysis of Genera Assemblages
3. Results
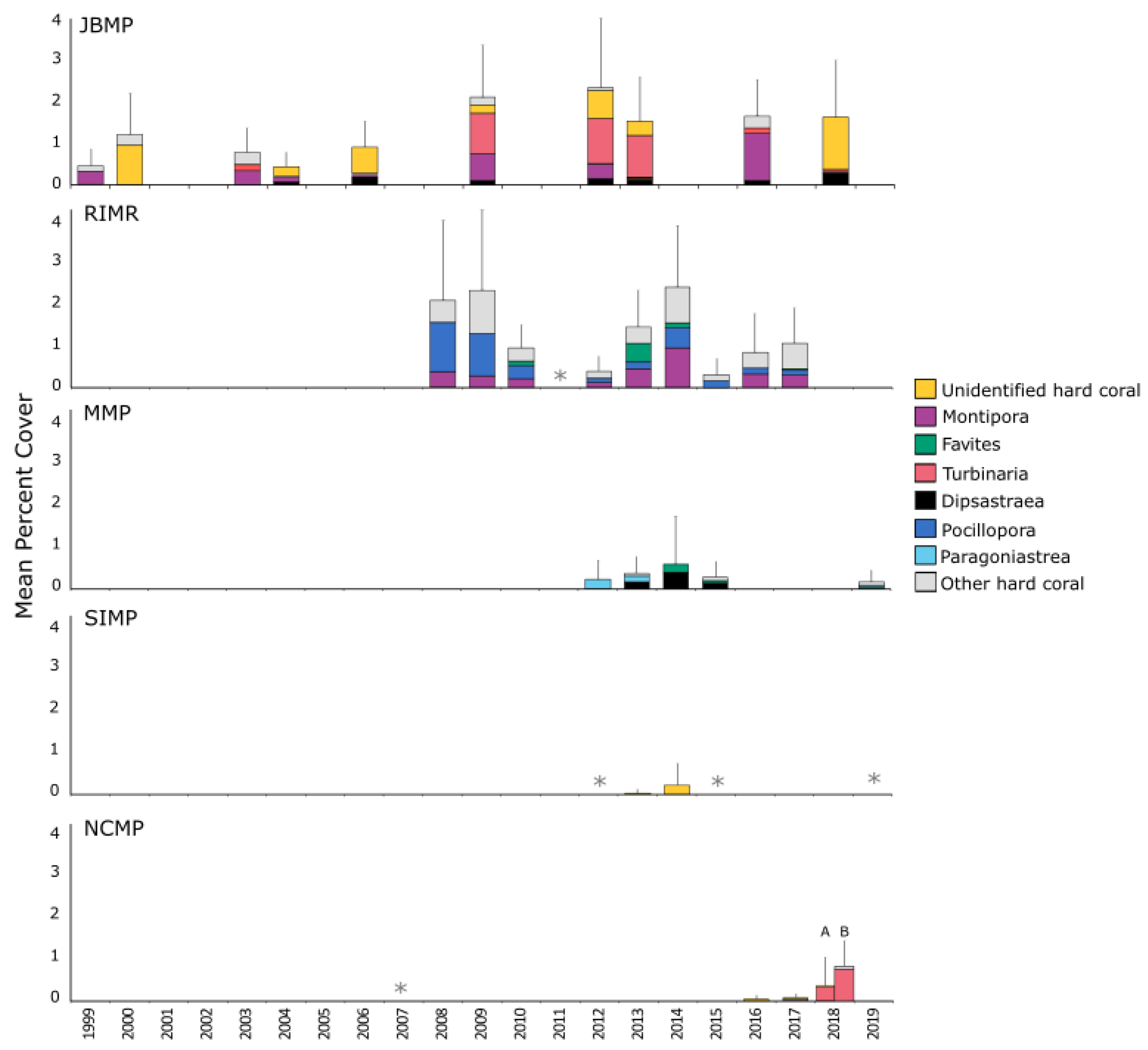
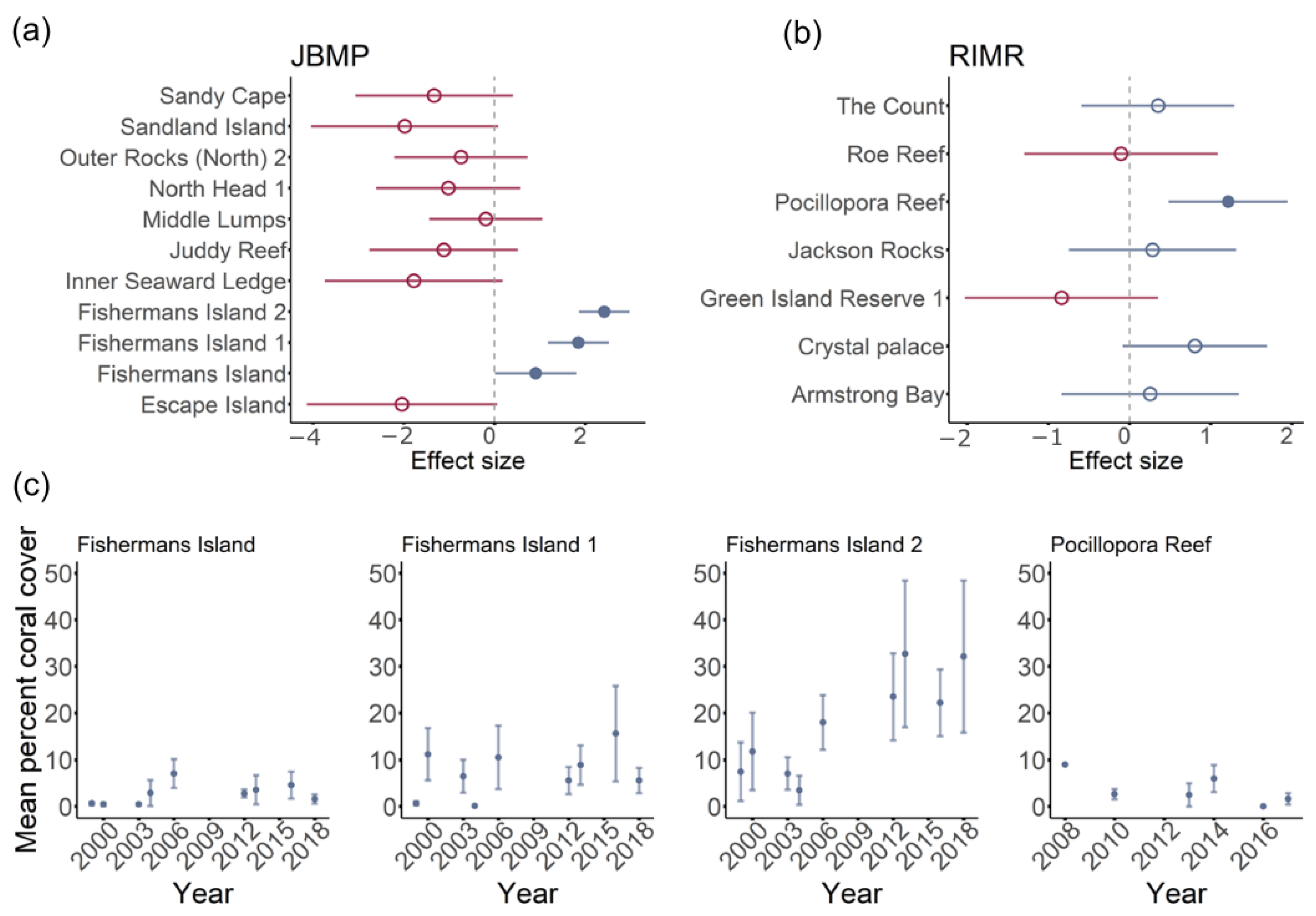
3.1. Total Hard-Coral Coverage
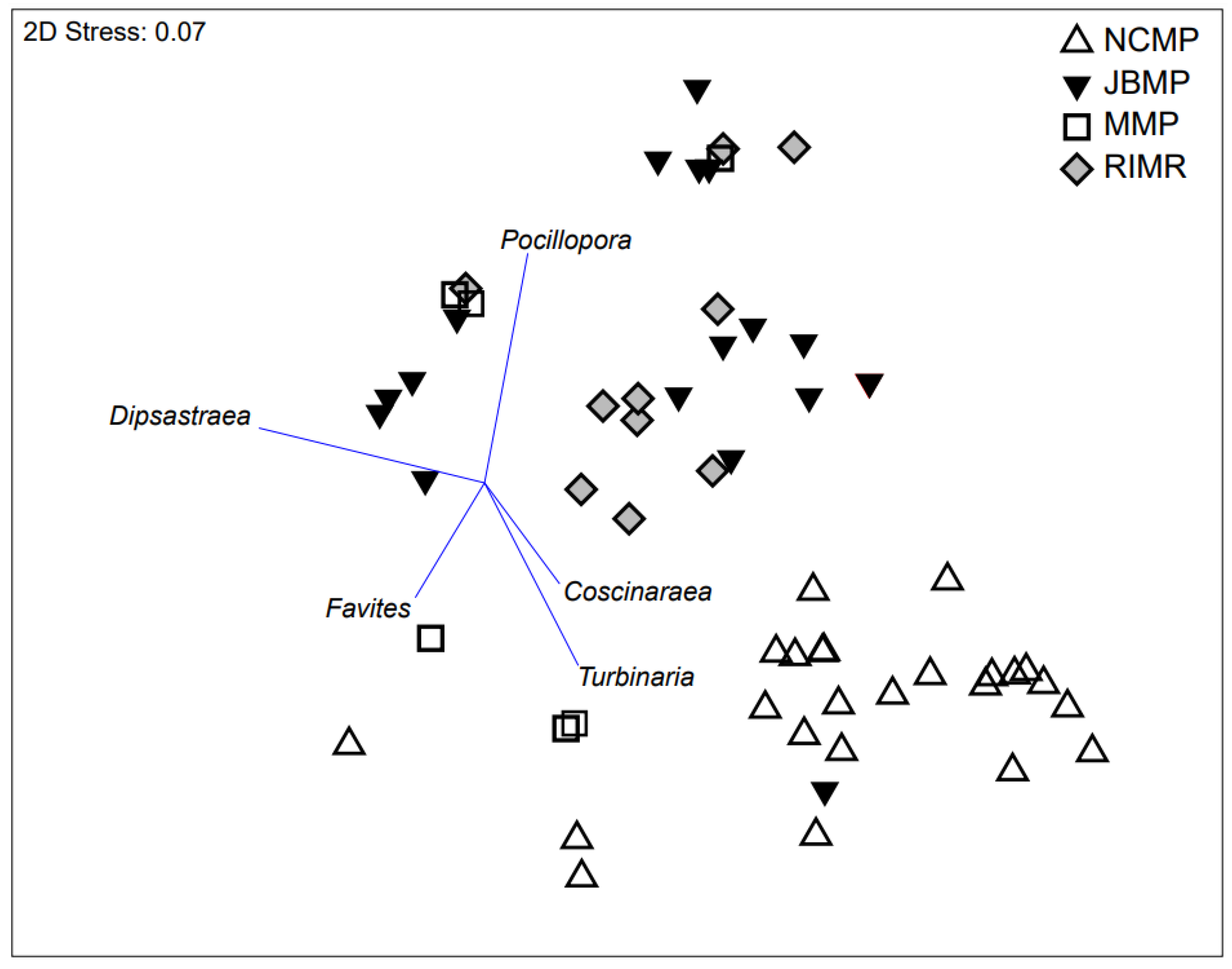
3.2. Analysis of Genera Assemblages
4. Discussion
4.1. Total Coral Cover
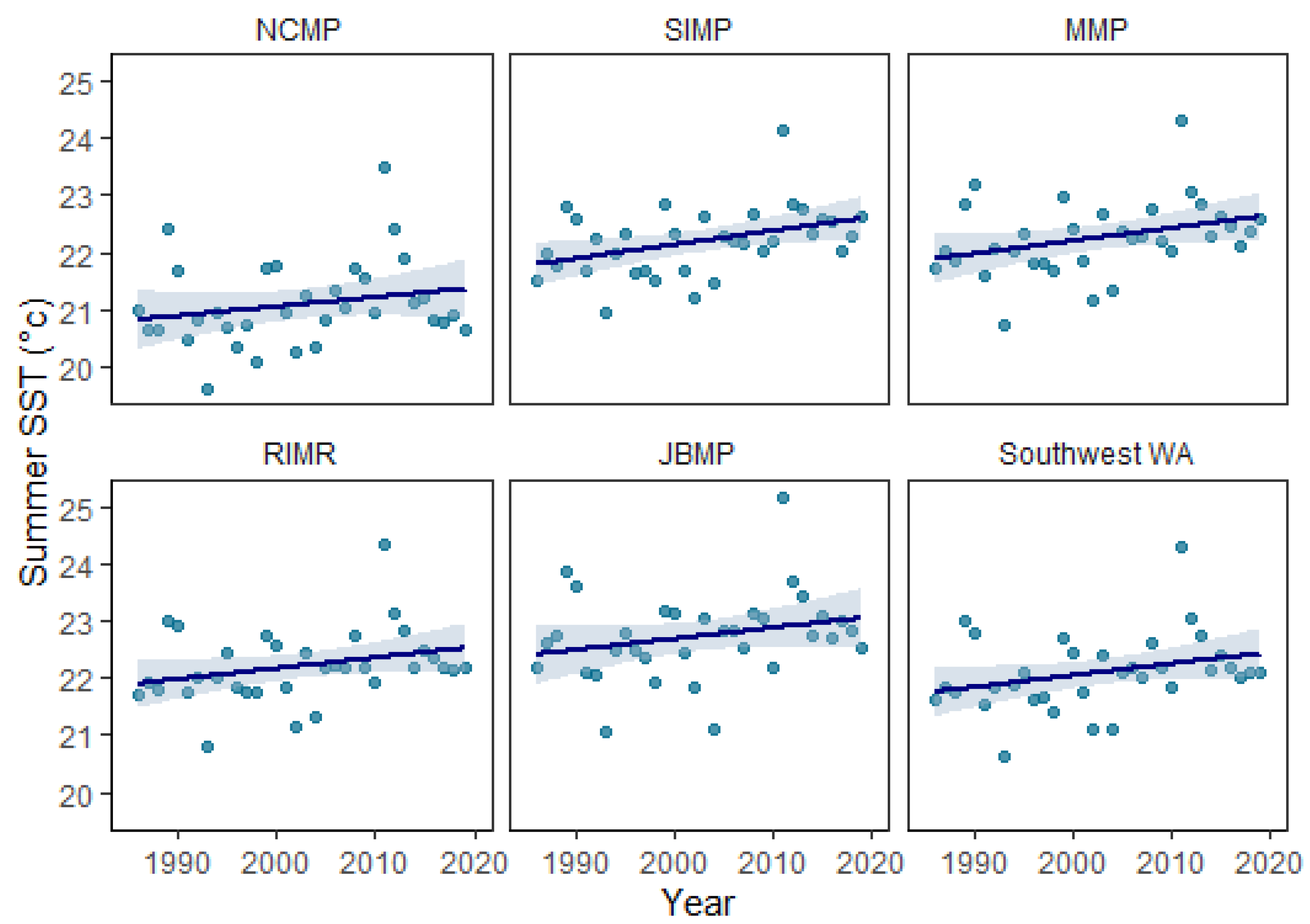
4.2. Temporal Trends in Coral Cover
4.3. Coral Genera Assemblages
5. Summary and Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wernberg, T.; Russell, B.D.; Thomsen, M.S.; Gurgel, C.F.D.; Bradshaw, C.J.A.; Poloczanska, E.S.; Connell, S.D. Seaweed communities in retreat from ocean warming. Curr. Biol. 2011, 21, 1828–1832. [Google Scholar] [CrossRef] [PubMed]
- Wernberg, T.; Bennett, S.; Babcock, R.C.; De Bettignies, T.; Cure, K.; Depczynski, M.; Dufois, F.; Fromont, J.; Fulton, C.J.; Hovey, R.K.; et al. Climate-driven regime shift of a temperate marine ecosystem. Science 2016, 353, 169–172. [Google Scholar] [CrossRef] [Green Version]
- Orth, R.J.; Carruthers, T.J.B.; Dennison, W.C.; Duarte, C.M.; Fourqurean, J.W.; Heck, K.L.; Hughes, A.R.; Kendrick, G.A.; Kenworthy, W.J.; Olyarnik, S.; et al. A global crisis for seagrass ecosystems. Bioscience 2006, 56, 987–996. [Google Scholar] [CrossRef] [Green Version]
- Strydom, S.; Murray, K.; Wilson, S.; Huntley, B.; Rule, M.; Heithaus, M.; Bessey, C.; Kendrick, G.A.; Burkholder, D.; Fraser, M.W.; et al. Too hot to handle: Unprecedented seagrass death driven by marine heatwave in a World Heritage Area. Glob. Chang. Biol. 2020, 26, 3525–3538. [Google Scholar] [CrossRef] [PubMed]
- Tuckett, C.A.; de Bettignies, T.; Fromont, J.; Wernberg, T. Expansion of corals on temperate reefs: Direct and indirect effects of marine heatwaves. Coral Reefs 2017, 36, 947–956. [Google Scholar] [CrossRef] [Green Version]
- Day, P.B.; Stuart-Smith, R.D.; Edgar, G.J.; Bates, A.E. Species’ thermal ranges predict changes in reef fish community structure during 8 years of extreme temperature variation. Divers. Distrib. 2018, 24, 1036–1046. [Google Scholar] [CrossRef] [Green Version]
- Mulders, Y.; Wernberg, T. Fifteen years in a global warming hotspot: Changes in subtidal mobile invertebrate communities. Mar. Ecol. Prog. Ser. 2020, 656, 227–238. [Google Scholar] [CrossRef]
- IPCC. IPCC, 2019: IPCC Special Report on the Ocean and Cryosphere in a Changing Climate; Pörtner, H.-O., Roberts, D.C., Masson-Delmotte, V., Zhai, P., Tignor, M., Poloczanska, E., Mintenbeck, K., Alegría, A., Nicolai, M., Okem, A., et al., Eds.; Intergovernmental Panel on Climate Change: Geneva, Switzerland, 2019.
- Cheung, W.W.L.; Meeuwig, J.J.; Feng, M.; Harvey, E.S.; Lam, V.W.Y.; Langlois, T.; Slawinski, D.; Sun, C.; Pauly, D. Climate-change induced tropicalisation of marine communities in Western Australia. Mar. Freshw. Res. 2012, 63, 415. [Google Scholar] [CrossRef] [Green Version]
- Vergés, A.; Steinberg, P.D.; Hay, M.E.; Poore, A.G.B.; Campbell, A.H.; Ballesteros, E.; Heck, K.L.; Booth, D.J.; Coleman, M.A.; Feary, D.A.; et al. The tropicalization of temperate marine ecosystems: Climate-mediated changes in herbivory and community phase shifts. Proc. R. Soc. B Biol. Sci. 2014, 281, 20140846. [Google Scholar] [CrossRef]
- Vergés, A.; McCosker, E.; Mayer-Pinto, M.; Coleman, M.A.; Wernberg, T.; Ainsworth, T.; Steinberg, P.D. Tropicalisation of temperate reefs: Implications for ecosystem functions and management actions. Funct. Ecol. 2019, 33, 1000–1013. [Google Scholar] [CrossRef] [Green Version]
- Figueira, W.F.; Booth, D.J. Increasing ocean temperatures allow tropical fishes to survive overwinter in temperate waters. Glob. Chang. Biol. 2010, 16, 506–516. [Google Scholar] [CrossRef]
- Cure, K.; Hobbs, J.A.; Harvey, E.S. High recruitment associated with increased sea temperatures towards the southern range edge of a Western Australian endemic reef fish Choerodon rubescens (family Labridae). Env. Biol. Fishes 2014, 98, 1059–1067. [Google Scholar] [CrossRef]
- Lenanton, R.C.J.; Dowling, C.E.; Smith, K.A.; Fairclough, D.V.; Jackson, G. Potential influence of a marine heatwave on range extensions of tropical fishes in the eastern Indian Ocean—Invaluable contributions from amateur observers. Reg. Stud. Mar. Sci. 2017, 13, 19–31. [Google Scholar] [CrossRef]
- Fowler, A.M.; Parkinson, K.; Booth, D.J. New poleward observations of 30 tropical reef fishes in temperate southeastern Australia. Mar. Biodivers. 2018, 48, 2249–2254. [Google Scholar] [CrossRef]
- Precht, W.; Aronson, R.B. Climate flickers and range shifts of reef corals. Front. Ecol. Environ. 2004, 2, 307–314. [Google Scholar] [CrossRef]
- Thomson, D. Range extension of the hard coral Goniopora norfolkensis (veron & pichon 1982) to the south-east Indian Ocean. J. R. Soc. West. Aust. 2010, 93, 115–117. [Google Scholar]
- Yamano, H.; Sugihara, K.; Nomura, K. Rapid poleward range expansion of tropical reef corals in response to rising sea surface temperatures. Geophys. Res. Lett. 2011, 38, 1–6. [Google Scholar] [CrossRef]
- Baird, A.H.; Sommer, B.; Madin, J.S. Pole-ward range expansion of Acropora spp. along the east coast of Australia. Coral Reefs 2012, 31, 1063. [Google Scholar] [CrossRef]
- Szabo, B. Uranium-series age of coral reef growth on Rottnest Island, Western Australia. Mar. Geol. 1979, 29, M11–M15. [Google Scholar] [CrossRef]
- Greenstein, B.; Pandolfi, J.M. Escaping the heat: Range shifts of reef coral taxa in coastal Western Australia. Glob. Chang. Biol. 2008, 14, 513–528. [Google Scholar] [CrossRef]
- Marsh, L.M. The occurrence and growth of Acropora in extra-tropical waters off Perth, Western Australia. In Proceedings of the Seventh International Coral Reef Symposium; University of Guam: Mangilao, GU, USA, 1992; Volume 2, pp. 1233–1238. [Google Scholar]
- Couce, E.; Ridgwell, A.; Hendy, E.J. Future habitat suitability for coral reef ecosystems under global warming and ocean acidification. Glob. Chang. Biol. 2013, 19, 3592–3606. [Google Scholar] [CrossRef] [Green Version]
- Madin, J.; Allen, A.; Baird, A.; Pandolfi, J.; Sommer, B. Scope for latitudinal extension of reef corals is species specific. Front. Biogeogr. 2016, 8, e29328. [Google Scholar] [CrossRef] [Green Version]
- Muir, P.R.; Wallace, C.C.; Done, T.; Aguirre, J.D. Limited scope for latitudinal extension of reef corals. Science 2015, 348, 1135–1138. [Google Scholar] [CrossRef] [PubMed]
- Pearse, V.; Muscatine, L. Role of symbiotic algae (zooxanthellae) in coral calcification. Biol. Bull. 1971, 141, 350–363. [Google Scholar] [CrossRef]
- Gattuso, J.-P.; Allemand, D.; Frankignoulle, M. Photosynthesis and calcification at cellular, organismal and community levels in coral reefs: A review on Interactions and control by carbonate chemistry. Am. Zool. 1999, 39, 160–183. [Google Scholar] [CrossRef] [Green Version]
- Grigg, R. Darwin Point: A threshold for atoll formation. Coral Reefs 1982, 1, 29–34. [Google Scholar] [CrossRef]
- Johannes, R.E.; Wiebe, W.J.; Crossland, C.J.; Rimmerl, D.W.; Smith, S.V. Latitudinal limits of coral reef growth. Mar. Ecol. Prog. Ser. 1983, 11, 105–111. [Google Scholar] [CrossRef]
- Kleypas, J.A.; McManus, J.; Menez, L. Environmental limits to coral reef development: Where do we draw the line? Am. Zool. 1999, 39, 146–159. [Google Scholar] [CrossRef]
- Cook, C.; Logan, A.; Ward, J.; Luckhurst, B.; Berg, C., Jr. Elevated temperatures and bleaching on a high latitude coral reef: The 1988 Bermuda event. Coral Reefs 1990, 9, 45–49. [Google Scholar] [CrossRef]
- Celliers, L.; Schleyer, M.H. Coral bleaching on high-latitude marginal reefs at Sodwana Bay, South Africa. Mar. Pollut. Bull. 2002, 44, 1380–1387. [Google Scholar] [CrossRef]
- Harrison, P.L.; Dalton, S.J.; Carroll, A.G. Extensive coral bleaching on the world’s southernmost coral reef at Lord Howe Island, Australia. Coral Reefs 2011, 30, 775. [Google Scholar] [CrossRef] [Green Version]
- Abdo, D.; Bellchambers, L.M.; Evans, S.N. Turning up the heat: Increasing temperature and coral bleaching at the high latitude coral reefs of the Houtman Abrolhos Islands. PLoS ONE 2012, 7, e43878. [Google Scholar] [CrossRef] [Green Version]
- Moore, J.A.Y.; Bellchambers, L.M.; Depczynski, M.R.; Evans, R.D.; Evans, S.N.; Field, S.N.; Friedman, K.J.; Gilmour, J.P.; Holmes, T.H.; Middlebrook, R.; et al. Unprecedented mass bleaching and loss of coral across 12° of latitude in Western Australia in 2010–11. PLoS ONE 2012, 7, e51807. [Google Scholar] [CrossRef] [Green Version]
- DeBose, J.L.; Nuttall, M.F.; Hickerson, E.L.; Schmahl, G.P. A high-latitude coral community with an uncertain future: Stetson Bank, northwestern Gulf of Mexico. Coral Reefs 2012, 32, 255–267. [Google Scholar] [CrossRef]
- Bridge, T.C.L.; Ferrari, R.; Bryson, M.; Hovey, R.; Figueira, W.F.; Williams, S.B.; Pizarro, O.; Harborne, A.R.; Byrne, M. Variable responses of benthic communities to anomalously warm sea temperatures on a high-latitude coral reef. PLoS ONE 2014, 9, 1–20. [Google Scholar] [CrossRef]
- Ross, C.L.; Schoepf, V.; DeCarlo, T.M.; McCulloch, M.T. Mechanisms and seasonal drivers of calcification in the temperate coral Turbinaria reniformis at its latitudinal limits. Proc. R. Soc. B 2018, 285, 20180215. [Google Scholar] [CrossRef] [PubMed]
- Tuckett, C.A.; Wernberg, T. High latitude corals tolerate severe cold spell. Front. Mar. Sci. 2018, 5, 1–6. [Google Scholar] [CrossRef] [Green Version]
- van Hooidonk, R.; Maynard, J.A.; Manzello, D.; Planes, S. Opposite latitudinal gradients in projected ocean acidification and bleaching impacts on coral reefs. Glob. Chang. Biol. 2014, 20, 103–112. [Google Scholar] [CrossRef] [PubMed]
- Veron, J.E.N. Corals of the World; Stafford-Smith, M., Ed.; Australian Institute of Marine Science: Townsville, Australia, 2000; Volume 3. [Google Scholar]
- Veron, J.; Marsh, L. Hermatypic corals of Western Australia. Records and annotated species list. Rec. West. Aust. Mus. 1988, 29, 1–136. [Google Scholar]
- Kendrick, A.; Wilson, S.; Friedman, K.; Waples, K.; Whiting, S.; Holmes, T.; Rule, M.; Halford, A.; Quartermaine, T.; Bobojcov, A.; et al. Strategic marine ecological research priorities for CALM act marine parks and reserves 2016–2021. Conserv. Sci. West. Aust. 2016, 10, 1–15. [Google Scholar]
- Smale, D.A.; Kendrick, G.; Waddington, K.I.; Van Niel, K.P.; Meeuwig, J.J.; Harvey, E. Benthic assemblage composition on subtidal reefs along a latitudinal gradient in Western Australia. Estuar. Coast. Shelf Sci. 2010, 86, 83–92. [Google Scholar] [CrossRef]
- Hutchins, J.; Pearce, A. Influence of the leeuwin current on recruitment of tropical reef fishes at Rottnest Island, Western Australia. Bull. Mar. Sci. 1994, 54, 245–255. [Google Scholar]
- Pearce, A.; Hutchins, B.; Hoschke, A.; Fearns, P. Record high damselfish recruitment at Rottnest Island, Western Australia, and the potential for climate-induced range extension. Reg. Stud. Mar. Sci. 2016, 8, 77–88. [Google Scholar] [CrossRef] [Green Version]
- Hatcher, B.G. Coral reefs in the Leeuwin Current—An ecological perspective. J. R. Soc. West. Aust. 1991, 74, 115–127. [Google Scholar]
- Gilmour, J.; Speed, C.W.; Babcock, R. Coral reproduction in Western Australia. PeerJ 2016, 4, e2010. [Google Scholar] [CrossRef] [Green Version]
- Wilson, S.K.; Kendrick, A.; Wilson, B. The North-Western Margin of Australia in World Seas: An Environmental Evaluation; Chapter 14; Academic Press: Cambridge, MA, USA, 2019. [Google Scholar]
- Thomson, D.P.; Frisch, A. Extraordinarily high coral cover on a nearshore, high-latitude reef in south-west Australia. Coral Reefs 2010, 29, 923–927. [Google Scholar] [CrossRef]
- Richards, Z.; Kirkendale, L.; Moore, G.; Hosie, A.; Huisman, J.; Bryce, M.; Marsh, L.; Bryce, C.; Hara, A.; Wilson, N.; et al. Marine biodiversity in temperate Western Australia: Multi-taxon surveys of Minden and Roe Reefs. Diversity 2016, 8, 7. [Google Scholar] [CrossRef] [Green Version]
- Ross, C.L.; Falter, J.L.; Schoepf, V.; McCulloch, M.T. Perennial growth of hermatypic corals at Rottnest Island, Western Australia (32° S). PeerJ 2015, 3, e781. [Google Scholar] [CrossRef]
- DEC. Ngari Capes Marine Park; Management Plan 74 2013–2023; Department of Environment and Conservation: Perth, Australia, 2013.
- Pearce, A.; Lenanton, R.; Jackson, G.; Moore, J.; Feng, M.; Gaughan, D. The “Marine Heat Wave” off Western Australia during the Summer of 2010/11 Fisheries Research Report No. 222; Western Australian Fisheries and Marine Research Laboratories: Hillarys, Australia, 2011. [Google Scholar]
- Smale, D.A.; Wernberg, T. Extreme climatic event drives range contraction of a habitat-forming species. Proc. R. Soc. B 2013, 280, 20122829. [Google Scholar] [CrossRef] [PubMed]
- Graham, N.A.J.; Wilson, S.K.; Jennings, S.; Polunin, N.V.C.; Bijoux, J.P.; Robinson, J. Dynamic fragility of oceanic coral reef ecosystems. Proc. Natl. Acad. Sci. USA 2006, 103, 8425–8429. [Google Scholar] [CrossRef] [Green Version]
- Jupiter, S.; Roff, G.; Marion, G.; Henderson, M.; Schrameyer, V.; McCulloch, M.; Hoegh-Guldberg, O. Linkages between coral assemblages and coral proxies of terrestrial exposure along a cross-shelf gradient on the southern Great Barrier Reef. Coral Reefs 2008, 27, 887–903. [Google Scholar] [CrossRef]
- Edgar, G.J.; Cooper, A.; Baker, S.C.; Barker, W.; Barrett, N.S.; Becerro, M.A.; Bates, A.E.; Brock, D.; Ceccarelli, D.M.; Clausius, E.; et al. Reef Life Survey: Establishing the ecological basis for conservation of shallow marine life. Biol. Conserv. 2020, 252, 108855. [Google Scholar] [CrossRef]
- Kohler, K.E.; Gill, S. Coral Point Count with Excel extensions (CPCe): A Visual Basic program for the determination of coral and substrate coverage using random point count methodology. Comput. Geosci. 2006, 32, 1259–1269. [Google Scholar] [CrossRef]
- Lin, X.; Zhang, D. Inference in generalized additive mixed models by using smoothing splines. J. R. Stat. Soc. Ser. B Stat. Methodol. 1999, 61, 381–400. [Google Scholar] [CrossRef] [Green Version]
- Tweedie, M.C.K. An index which distinguishes between some important exponential families. Statistics: Applications and New Directions. In Proceedings of the Indian Statistical Institute Golden Jubilee International Conference; Ghosh, J.K., Roy, J., Eds.; Indian Statistical Institute: Calcutta, Indian, 1984; pp. 579–604. [Google Scholar]
- Burnham, K.; Anderson, D.R. Model Selection and Multimodel Inference: A Practical Information-Theoretical Approach, 2nd ed.; Springer: New York, NY, USA, 2002. [Google Scholar]
- Wood, S. Fast stable restricted maximum likelihood and marginal likelihood estimation of semiparametric generalized linear models. J. R. Stat. Soc. B 2011, 73, 3–36. [Google Scholar] [CrossRef] [Green Version]
- Wood, S. Generalized Additive Models: An Introduction with R, 2nd ed.; Chapman and Hall/CRC: Boca Raton, FL, USA, 2017. [Google Scholar]
- Lenth, R. Emmeans: Estimated Marginal Means, Aka Least-Squares Means. 2021. Available online: https://CRAN.R-project.org/package=emmeans (accessed on 15 December 2020).
- Clarke, K.; Gorley, R.; Somerfield, P.; Warwick, R. Change in Marine Communities: An Apporach to Statistical Analysis and Interpretation, 3rd ed.; PRIMER-E Ltd.: Auckland, New Zealand, 2014. [Google Scholar]
- Anderson, M.; Gorley, R.; Clarke, K. PERMANOVA+ for PRIMER: Guide to Software and Statistical Methods. [PRIMER-E]; PRIMER-E Ltd.: Ivybridge, UK, 2008. [Google Scholar]
- Clarke, K.; Gorley, R. Getting Started with PRIMER v7 [PRIMER-E]; PRIMER-E Ltd.: Ivybridge, UK, 2015. [Google Scholar]
- Clarke, K.; Green, R. Statistical design and analysis for a’biological effects’ study. Mar. Ecol. Prog. Ser. 1988, 46, 213–226. [Google Scholar] [CrossRef]
- Miller, M.W. Growth of a temperate coral: Effects of temperature, light, depth, and heterotrophy. Mar. Ecol. Prog. Ser. 1995, 122, 217–225. [Google Scholar] [CrossRef] [Green Version]
- Goffredo, S.; Caroselli, E.; Mattioli, G.; Pignotti, E.; Dubinsky, Z.; Zaccanti, F. Inferred level of calcification decreases along an increasing temperature gradient in a Mediterranean endemic coral. Limnol. Oceanogr. 2009, 54, 930–937. [Google Scholar] [CrossRef] [Green Version]
- Mizerek, T.L.; Baird, A.H.; Beaumont, L.J.; Madin, J.S. Environmental tolerance governs the presence of reef corals at latitudes beyond reef growth. Glob. Ecol. Biogeogr. 2016, 25, 979–987. [Google Scholar] [CrossRef]
- Veron, J.E.N. Corals in Space and Time: The Biogeography and Evolution of the Scleractinia; Cornell University Press: Ithaca, NY, USA, 1995; pp. 1–321. [Google Scholar]
- Lough, J.M.; Barnes, D.J. Environmental controls on growth of the massive coral Porites. J. Exp. Mar. Biol. Ecol. 2000, 245, 225–243. [Google Scholar] [CrossRef]
- Coles, S.L.; Jokiel, P.L. Effects of temperature on photosynthesis and respiration in hermatypic corals. Mar. Biol. 1977, 43, 209–216. [Google Scholar] [CrossRef]
- Burton, E.A.; Walter, L.M. Relative precipitation rates of aragonite and Mg calcite from seawater: Temperature or carbonate ion control? Geology 1987, 15, 111. [Google Scholar] [CrossRef]
- Tribble, G.; Randall, R. A description of the high-latitude shallow water coral communities of Miyake-jima, Japan. Coral Reefs 1986, 4, 151–159. [Google Scholar] [CrossRef]
- Denis, V.; Mezaki, T.; Tanaka, K.; Kuo, C.; De Palmas, S.; Keshavmurthy, S.; Chen, C. Coverage, diversity, and functionality of a high-latitude coral community (Tatsukushi, Shikoku Island, Japan). PLoS ONE 2013, 8, e54330. [Google Scholar] [CrossRef]
- Harriott, V.; Banks, S. Latitudinal variation in coral communities in eastern Australia: A qualitative biophysical model of factors regulating coral reefs. Coral Reefs 2002, 21, 83–94. [Google Scholar] [CrossRef]
- Cresswell, G.; Golding, T.J. Observations of a south-flowing current in the southeastern Indian Ocean. Deep. Res. 1980, 27A, 449–466. [Google Scholar] [CrossRef]
- Adam, A.A.S.; Garcia, R.A.; Galaiduk, R.; Tomlinson, S.; Radford, B.; Thomas, L.; Richards, Z.T. Diminishing potential for tropical reefs to function as coral diversity strongholds under climate change conditions. Divers. Distrib. 2021, 27, 2245–2261. [Google Scholar] [CrossRef]
- Benthuysen, J.; Feng, M.; Zhong, L. Spatial patterns of warming off Western Australia during the 2011 Ningaloo Niño: Quantifying impacts of remote and local forcing. Cont. Shelf Res. 2014, 91, 232–246. [Google Scholar] [CrossRef]
- Caputi, N.; Fletcher, W.J.; Pearce, A.; Chubb, C.F. Effect of the Leeuwin Current on the recruitment of fish and invertebrates along the Western Australian coast. Mar. Freshw. Res. 1996, 47, 147–155. [Google Scholar] [CrossRef]
- Markey, K.L.; Abdo, D.A.; Evans, S.N.; Bosserelle, C. Keeping it local: Dispersal limitations of coral larvae to the high latitude coral reefs of the Houtman Abrolhos Islands. PLoS ONE 2016, 11, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Thomas, L.; Kennington, W.J.; Evans, R.D.; Kendrick, G.A.; Stat, M. Restricted gene flow and local adaptation highlight the vulnerability of high-latitude reefs to rapid environmental change. Glob. Chang. Biol. 2017, 23, 2197–2205. [Google Scholar] [CrossRef]
- Miller, M.W.; Hay, M.E. Coral-seaweed-grazer-nutrient interactions on temperate reefs. Ecol. Monogr. 1996, 66, 323–344. [Google Scholar] [CrossRef]
- Bennett, S.; Wernberg, T.; Harvey, E.S.; Santana-garcon, J.; Saunders, B.J. Tropical herbivores provide resilience to a climate-mediated phase shift on temperate reefs. Ecol. Lett. 2015, 18, 714–723. [Google Scholar] [CrossRef]
- Wyrwoll, K.; Greenstein, B.; Kendrick, G.; Chen, G. The palaeoceanography of the Leeuwin Current: Implications for a future world. J. R. Soc. West. Aust. 2009, 92, 37–51. [Google Scholar]
- Black, R.; Prince, J. Fauna associated with the coral Pocillopora damicornis at the Southern limit of its distribution in Western Australia. J. Biogeogr. 1983, 10, 135–152. [Google Scholar] [CrossRef]
- Ward, S. The effect of damage on the growth, reproduction and storage of lipids in the scleractinian coral Pocillopora damicornis (Linnaeus). J. Exp. Mar. Bio. Ecol. 1995, 187, 193–206. [Google Scholar] [CrossRef]
- Le Nohaïc, M.; Ross, C.L.; Cornwall, C.E.; Comeau, S.; Lowe, R.; McCulloch, M.T.; Schoepf, V. Marine heatwave causes unprecedented regional mass bleaching of thermally resistant corals in northwestern Australia. Sci. Rep. 2017, 7, 14999. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stirling, C.; Esat, T.; McCulloch, M.T.; Lambeck, K. High-precision U-series dating of corals from Western Australia and implications for the timing and duration of the Last Interglacial. Earth Planet. Sci. Lett. 1995, 135, 115–130. [Google Scholar] [CrossRef]
- Morgan, K.; Perry, C.; Smithers, S.; Johnson, J.; Daniell, J. Evidence of extensive reef development and high coral cover in nearshore environments: Implications for understanding coral adaptation in turbid settings. Sci. Rep. 2016, 6, 1–10. [Google Scholar]
- Veron, J. Corals of Australia and the Indo-Pacific; Angus and Robertson Publishers: North Ryde, Australia, 1986. [Google Scholar]
- Thomson, D.P.; Bearham, D.; Graham, F.; Eagle, J. High latitude, deeper water coral bleaching at Rottnest Island, Western Australia. Coral Reefs 2011, 30, 775. [Google Scholar] [CrossRef] [Green Version]
- Pante, E.; Dustan, P. Getting to the Point: Accuracy of Point Count in Monitoring Ecosystem Change. J. Mar. Biol. 2012, 2012, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Jokiel, P.L.; Rodgers, K.S.; Brown, E.K.; Kenyon, J.C.; Aeby, G.; Smith, W.R.; Farrell, F. Comparison of methods used to estimate coral cover in the Hawaiian Islands. PeerJ 2015, 3, e954. [Google Scholar] [CrossRef] [Green Version]
- Brock, R.; English, E.; Kenchington, E.; Tasker, M. The alphabet soup that protects cold-water corals in the North Atlantic. Mar. Ecol. Prog. Ser. 2009, 397, 355–360. [Google Scholar] [CrossRef]
- Huvenne, V.A.I.; Bett, B.J.; Masson, D.G.; Le Bas, T.P.; Wheeler, A. Effectiveness of a deep-sea cold-water coral Marine Protected Area, following eight years of fisheries closure. Biol. Conserv. 2016, 200, 60–69. [Google Scholar] [CrossRef] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Marine Park | Latitude (° S) | Longitude (° E) | Mean Annual SST (°C) | SST Increase (°C yr−1) | Max. DHW |
|---|---|---|---|---|---|
| JBMP | 30.125 | 114.925 | 21.0 | 0.0193 (r2 = 0.10, p = 0.15) | 28.0 |
| RIMR | 32.025 | 115.475 | 20.5 | 0.0206 (r2 = 0.10, p = 0.07) | 24.9 |
| MMP | 31.825 | 115.675 | 20.3 | 0.0224 (r2 = 0.12, p = 0.05) | 23.3 |
| SIMP | 32.325 | 115.675 | 20.1 | 0.0244 (r2 = 0.17, p = 0.02) | 22.3 |
| NCMP | 33.825 | 114.925 | 19.5 | 0.0164 (r2 = 0.05, p = 0.22) | 22.9 |
| South-west WA MPAs | - | - | 20.3 | 0.020 (r2 = 0.09, p = 0.08) | 28.0 |
| JBMP | EDF | ΔAICc | wAIC | Deviance Explained |
| Coral per cent cover ~ year + site | 14.7 | 0 | 0.99 | 49.7% |
| Coral per cent cover ~ 1 + site | 13.7 | 13.9 | 0.01 | 45.8% |
| RIMP | EDF | ΔAICc | wAIC | Deviance Explained |
| Coral per cent cover ~ year + site | 9.3 | 0 | 0.41 | 19.7% |
| Coral per cent cover ~ 1 + site | 8.2 | 0.7 | 0.59 | 18.8% |
| JBMP | RIMR | ||||||||
| df | MS | Pseudo-f | p-Value | df | MS | Pseudo-f | p-Value | ||
| PERMANOVA | Year | 9 | 8792.0 | 2.8 | <0.01 | 8 | 3609.5 | 1.0 | 0.5 |
| Site * | 8 | 4994.2 | 2.2 | <0.01 | 6 | 4885.8 | 1.9 | 0.01 | |
| Year × Site * | 27 | 3184.5 | 1.4 | 0.02 | 19 | 3654.9 | 1.4 | 0.05 | |
| Residual | 50 | 2302 | 16 | 2608 | |||||
| Total | 94 | 49 | |||||||
| ANOSIM | R-Stat | p-Value | R-Stat | p-Value | |||||
| Year (unordered) | 0.36 | 0.0001 | 0.25 | 0.035 | |||||
| Year (ordered) | 0.08 | >0.05 | 0.093 | >0.05 | |||||
| Site | 0.22 | 0.002 | 0.41 | 0.001 | |||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ross, C.L.; French, B.; Lester, E.K.; Wilson, S.K.; Day, P.B.; Taylor, M.D.; Barrett, N. Coral Communities on Marginal High-Latitude Reefs in West Australian Marine Parks. Diversity 2021, 13, 554. https://doi.org/10.3390/d13110554
Ross CL, French B, Lester EK, Wilson SK, Day PB, Taylor MD, Barrett N. Coral Communities on Marginal High-Latitude Reefs in West Australian Marine Parks. Diversity. 2021; 13(11):554. https://doi.org/10.3390/d13110554
Chicago/Turabian StyleRoss, Claire L., Ben French, Emily K. Lester, Shaun K. Wilson, Paul B. Day, Michael D. Taylor, and Neville Barrett. 2021. "Coral Communities on Marginal High-Latitude Reefs in West Australian Marine Parks" Diversity 13, no. 11: 554. https://doi.org/10.3390/d13110554