Changes in the Abundance of Danish Orchids over the Past 30 Years


Abstract
:
1. Introduction
2. Materials and Methods
2.1. Orchid Taxa in Denmark
2.2. Status, Biology, and Habitats of Danish Orchids
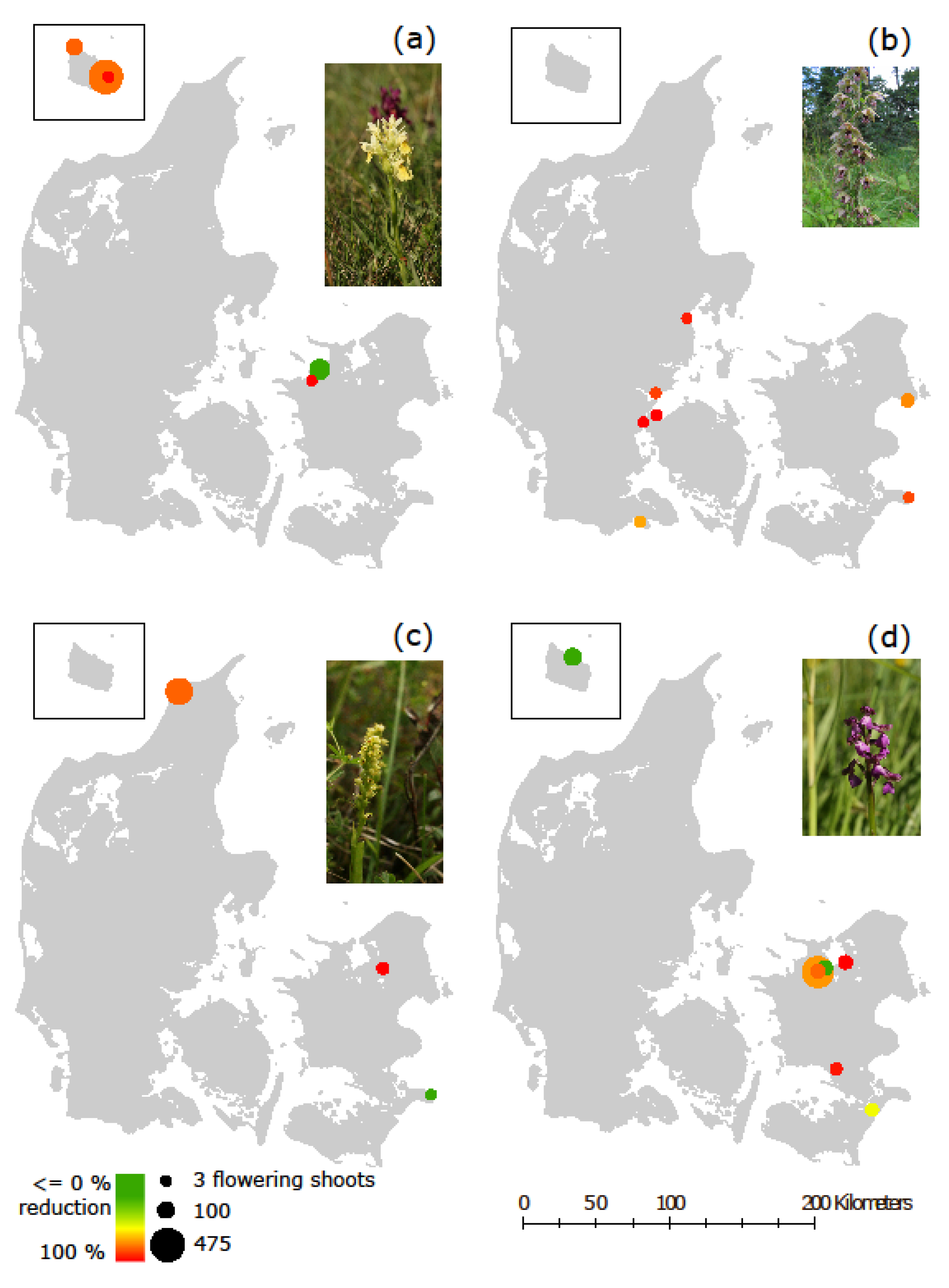
2.3. Orchid Censuses in Denmark
2.4. Orchid Abundance Data
2.5. Statistical Analysis
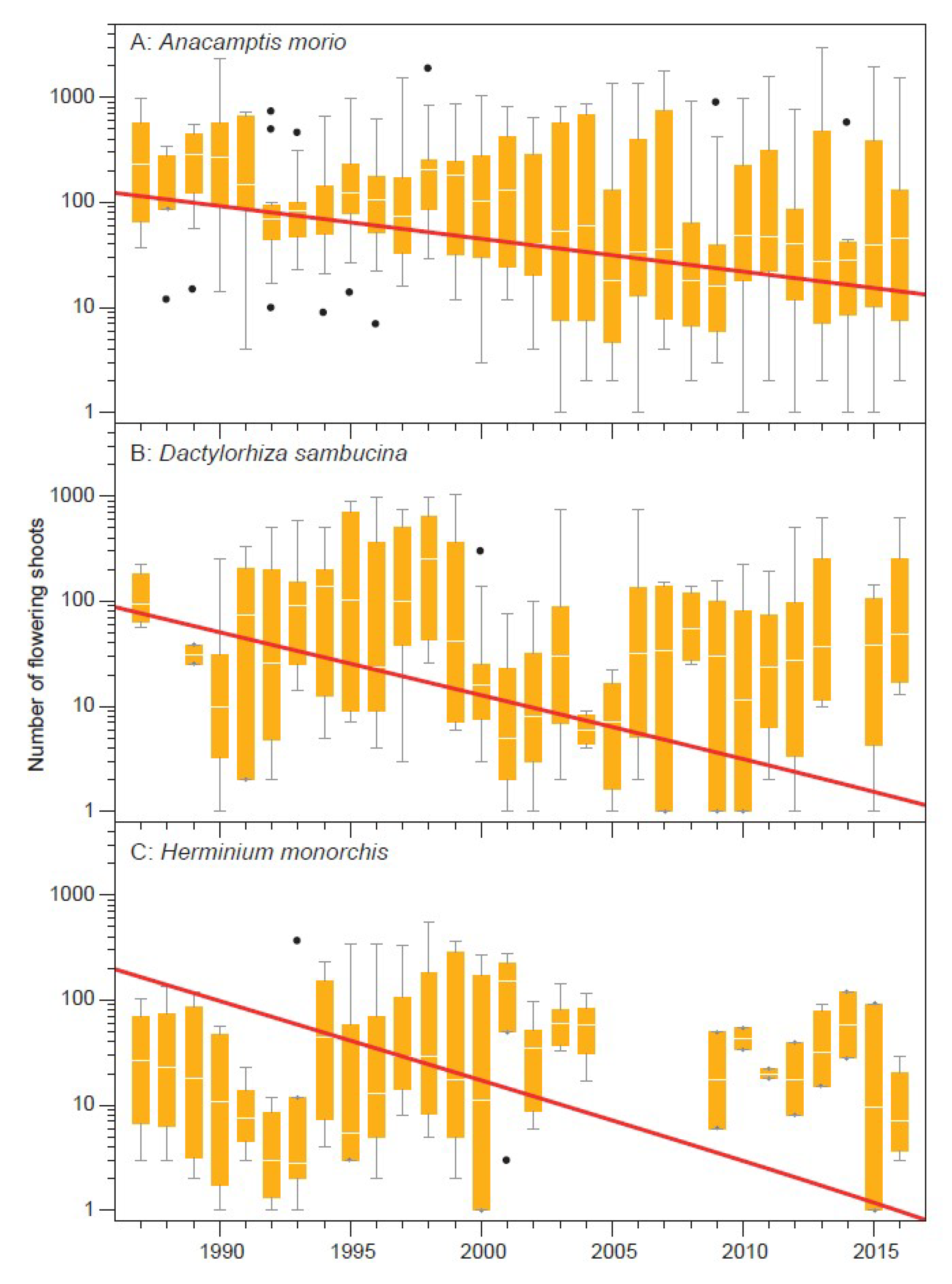
3. Results
4. Discussion
5. Conservation and Protection of Danish Orchids
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Kull, T.; Hutchings, M.J. A comparative analysis of decline in the distribution ranges of orchid species in Estonia and the United Kingdom. Biol. Conserv. 2006, 129, 31–39. [Google Scholar] [CrossRef]
- Tali, K.; Fay, M.F.; Bateman, R.M. Little genetic differentiation across Europe between early-flowering and late-flowering populations of the rapidly declining orchid Neotinea ustulata. Biol. J. Linn. Soc. 2006, 87, 13–25. [Google Scholar] [CrossRef]
- Vogt-Schilb, H.; Munoz, F.; Richard, F.; Schatz, B. Recent declines and range changes of orchids in Western Europe (France, Belgium and Luxembourg). Biol. Conserv. 2015, 190, 133–141. [Google Scholar] [CrossRef]
- Wind, P. Karplanter. In Den Danske Rødliste 2019; Moeslund, J.E., Ed.; DCE–Nationalt Center for Miljø og Energi, Aarhus Universitet: Rønde, Denmark, 2019; p. 31. [Google Scholar]
- Pedersen, H.A.; Faurholdt, N. Danmarks Vilde Orkidéer; Gyldendal: Copenhagen, Denmark, 2010. [Google Scholar]
- Djordjević, V.; Tsiftsis, S.; Lakušić, D.; Jovanovic, S.; Stevanović, V. Factors affecting the distribution and abundance of orchids in grasslands and herbaceous wetlands. Syst. Biodivers. 2016, 14, 1–16. [Google Scholar] [CrossRef]
- Hemrová, L.; Kotilínek, M.; Konečná, M.; Paulič, R.; Jersáková, J.; Těšitelová, T.; Knappová, J.; Münzbergová, Z. Identification of drivers of landscape distribution of forest orchids using germination experiment and species distribution models. Oecologia 2019, 190, 411–423. [Google Scholar] [CrossRef] [PubMed]
- McCormick, M.K.; Jacquemyn, H. What constrains the distribution of orchid populations? New Phytol. 2013, 202, 392–400. [Google Scholar] [CrossRef]
- Van der Heijden, M.G.A.; Martin, F.M.; Selosse, M.; Sanders, I.R. Mycorrhizal ecology and evolution: The past, the present, and the future. New Phytol. 2015, 205, 1406–1423. [Google Scholar] [CrossRef]
- Schiestl, F. On the success of a swindle: Pollination by deception in orchids. Naturwissenschaften 2005, 92, 255–264. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jacquemyn, H.; Brys, R.; Hermy, M.; Willems, J.H. Does nectar reward affect rarity and extinction probabilities of orchid species? An assessment using historical records from Belgium and the Netherlands. Biol. Conserv. 2005, 121, 257–263. [Google Scholar] [CrossRef]
- Mattiasson, G. Adam og Eva dör ut inom naturreseratet Kungsmarken. Svensk Bot. Tidskr. 1986, 80, 17–25. [Google Scholar]
- Schrautzer, J.; Fichtner, A.; Huckauf, A.; Rasran, L.; Jensen, K. Long-term population dynamics of Dactylorhiza incarnata (L.) Soó after abandonment and re-introduction of mowing. Flora Morphol. Distrib. Funct. Ecol. Plants 2011, 206, 622–630. [Google Scholar] [CrossRef]
- Ejrnæs, R.; Hansen, D.N.; Aude, E. Changing course of secondary succession in abandoned sandy fields. Biol. Conserv. 2003, 109, 343–350. [Google Scholar] [CrossRef]
- Catorci, A.; Cesaretti, S.; Gatti, R. Effect of long-term abandonment and spring grazing on floristic and functional composition of dry grasslands in a Central Apennine farmland. Polish J. Ecol. 2013, 61, 505–517. [Google Scholar]
- Whigham, D.F. Ecology of Woodland Herbs in Temperate Deciduous Forests. Annu. Rev. Ecol. Evol. Syst. 2004, 35, 583–621. [Google Scholar] [CrossRef] [Green Version]
- Miljøministeriet. Bekendtgørelse af 27 Jun 2016 om Fredning af Visse Dyre- Og Plantearter og Pleje af Tilskadekommet Vildt in Miljøministeriet; Miljø- og Fødevareministeriet: København, Danmark, 2016. [Google Scholar]
- Handley, J.; Heidel, B. Amerorchis Rotundifolia (Banks ex Pursh) Hultén (Roundleaf Orchid): A Technical Conservation Assessment; USDA Forest Service, Rocky Mountain Region: Houghton, MI, USA, 2005. [Google Scholar]
- Kull, T. Cypripedium calceolus L. J. Ecol. 1999, 87, 913–924. [Google Scholar] [CrossRef]
- Rocchio, J.; March, M.; Anderson, D.G. Epipactis gigantea Dougl. ex Hook. (stream orchid): A Technical Conservation Assessment; USDA Forest Service: Colorado State University: Fort Collins, CO, USA, 2006. [Google Scholar]
- Stewart, J. La Conservation des Orchidées Européennes T-PVS (89) 3; Conseil de l’Europe: Strasbourg, France, 1989. [Google Scholar]
- Stewart, J. Commercial trade. In Orchids–Status Survey and Conservation Action Plan; IUCN/SCC Orchid Specialist Group: Gland, Switzerland; Cambridge, UK, 1996; pp. 38–44. [Google Scholar]
- Wood, J.; Clements, M.; Muir, H. Plants in Peril, 2. Curtis’s Bot. Mag. 1984, 1, 139–142. [Google Scholar] [CrossRef]
- Phillips, R.D.; Barrett, M.D.; Dixon, K.; Hopper, S.D. Do mycorrhizal symbioses cause rarity in orchids? J. Ecol. 2011, 99, 858–869. [Google Scholar] [CrossRef]
- Dressler, R.L.; Rasmussen, H.N. Terrestrial Orchids: From Seed to Mycotrophic Plant. Syst. Bot. 1996, 21, 625. [Google Scholar] [CrossRef] [Green Version]
- Phillips, R.D.; Brown, A.P.; Dixon, K.; Hopper, S.D. Orchid biogeography and factors associated with rarity in a biodiversity hotspot, the Southwest Australian Floristic Region. J. Biogeogr. 2010, 38, 487–501. [Google Scholar] [CrossRef]
- Laroche, V.; Pellerin, S.; Brouillet, L. White Fringed Orchid as indicator of Sphagnum bog integrity. Ecol. Indic. 2012, 14, 50–55. [Google Scholar] [CrossRef]
- Newman, B. Orchids as Indicators of Ecosystem Health in Urban Bushland Fragments; Murdoch University: Murdoch, Australia, 2009. [Google Scholar]
- Sterk, A.A. Anacamptis pyramidalis bij Wijk aan Zee. Gorteria 1976, 8, 81–85. [Google Scholar]
- Tamm, C.O. Behaviour of some Orchid Population in a Changing Environment. In Observations on Permanent Plots, 1943–1990; Wells, T.C.E., Willems, J.H., Eds.; Population Ecology of Terrestrial Orchids: The Hague, The Netherlands, 1991; pp. 1–13. [Google Scholar]
- Morris, J.; Wells, T.C.E.; Willems, J.H. Population Ecology of Terrestrial Orchids. J. Ecol. 1993, 81, 202. [Google Scholar] [CrossRef]
- Vanhecke, L. Veranderingen in én kustpopulatie van de Rietorchis (Dactylorhiza praetermissa) tussen 1973 en 1985. Levende Nat. 1988, 2, 44–49. [Google Scholar]
- Vanhecke, L.E.M. Population dynamics of Dactylorhiza praetermissa in relation to topography and inundation. In Population Ecology of Terrestrial Orchids; Wells, T.C.E., Willems, J.H., Eds.; SPB Academic Publishing: The Hague, The Netherlands, 1991; pp. 15–32. [Google Scholar]
- Willems, J.H. Population dynamics of Spiranthes spiralis in South-Limburg, the Netherlands. Mém. Soc. R. Bot. Belgique 1989, 11, 115–121. [Google Scholar]
- Farrell, L. Population Changes and Management of Orchis Militaris at Two Sites in England; Wells, T.C.E., Willems, J.H., Eds.; SPB Academic Publishing: Amsterdam, The Netherlands, 1991; pp. 63–68. [Google Scholar]
- Waite, S.; Farrell, L. Population biology of the rare military orchid (Orchis militaris L.) at an established site in Suffolk, England. Bot. J. Linn. Soc. 1998, 126, 109–121. [Google Scholar] [CrossRef] [Green Version]
- Högström, S. Kärrnycklar, Orchis palustris, pa Gotland 1982–1990. Svensk Bot. Tidskr. 1991, 85, 355–376. [Google Scholar]
- Salkowski, H.-E. 20 Jahre Spiranthes spiralis (L.) Chevall. Im Rabengrund bei Wiesbaden. Ber. Arbeitskr. Heim. Orchid. 1990, 7, 73–76. [Google Scholar]
- Vogt-Schilb, H.; Geniez, P.; Pradel, R.; Richard, F.; Schatz, B. Inter-annual variability in flowering of orchids: Lessons learned from 8 years of monitoring in a Mediterranean region of France. Eur. J. Environ. Sci. 2013, 3, 129–137. [Google Scholar] [CrossRef]
- Kull, T. Population Dynamics of Nort Temperate Orchids. In Orchid Biology: Reviews and Perspectives; Kull, T., Arditti, J., Eds.; Kluwer Academic Publishers: Boston, MA, USA; London, UK, 2002. [Google Scholar]
- Løjtnant, B. Orchis militaris L. (Ridder-Gøgeurt)—Ny for Danmark. Flora Fauna 1981, 87, 59–62. [Google Scholar]
- Petersen, B.V.; Høyer-Nielsen, J. Biblomst, Ophrys apifera Huds—Nu i Danmark. URT 2015, 29, 16–19. [Google Scholar]
- Correll, N.H. Ridderen vender tilbage. URT 2016, 40, 42–45. [Google Scholar]
- Hartvig, P. Atlas Flora Danica; Gyldendal: Copenhagen, Denmark, 2015. [Google Scholar]
- Vogt-Schilb, H.; Pradel, R.; Geniez, P.; Hugot, L.; Delage, A.; Richard, F.; Schatz, B. Responses of orchids to habitat change in Corsica over 27 years. Ann. Bot. 2016, 118, 115–123. [Google Scholar] [CrossRef]
- Hagerup, O. Bestøvningen hos Liparis og Malaxis. Bot. Tidsskr. 1941, 45, 396–402. [Google Scholar]
- Swarts, N.; Dixon, K. Terrestrial orchid conservation in the age of extinction. Ann. Bot. 2009, 104, 543–556. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- European Union (EU). Council Directive 92/43/EEC of 21 May 1992 on the conservation of natural habitats and of wild fauna and flora in E. Commission. Off. J. Eur. Comm. 1992, 26, 7–50. [Google Scholar]
- Løjtnant, B. Overvagning af Orchideer 1987–1989. Flora Fauna 1991, 97, 63–121. [Google Scholar]
- Wind, P. Overvagning af Orkidéer 1998. Pavirkningsfaktorer; Arbejdsrapport; Danmarks Miljøundersøgleser, Miljø- og Energiministeriet: Rønde, Denmark, 1999. [Google Scholar]
- Rue, H.; Martino, S.; Chopin, N. Approximate Bayesian inference for latent Gaussian models using integrated nested Laplace approximations (with discussion). J. R. Stat. Soc. Ser. B 2009, 71, 319–392. [Google Scholar] [CrossRef]
- Blangiardo, M.; Cameletti, M.; Baio, G.; Rue, H. Spatial and spatio-temporal models with R-INLA. Spat. Spatio-Temporal Epidemiol. 2013, 4, 33–49. [Google Scholar] [CrossRef] [Green Version]
- Spiegelhalter, D.J.; Best, N.G.; Carlin, B.P.; van der Linde, A. Bayesian measures of model complexity and fit. J. R. Stat. Soc. Ser. B 2002, 64, 583–639. [Google Scholar] [CrossRef] [Green Version]
- Timmermann, A.; Damgaard, C.; Strandberg, M.; Svenning, J.-C. Pervasive early 21st-century vegetation changes across Danish semi-natural ecosystems: More losers than winners and a shift towards competitive, tall-growing species. J. Appl. Ecol. 2014, 52, 21–30. [Google Scholar] [CrossRef]
- Joffard, N.; Massol, F.; Grenié, M.; Montgelard, C.; Schatz, B. Effect of pollination strategy, phylogeny and distribution on pollination niches of Euro-Mediterranean orchids. J. Ecol. 2018, 107, 478–490. [Google Scholar] [CrossRef] [Green Version]
- Fay, M.F. Orchid conservation: How can we meet the challenges in the twenty-first century? Bot. Stud. 2018, 59, 16. [Google Scholar] [CrossRef] [Green Version]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| International Name | Taxon Status | Status 2020 | Monitoring Period |
|---|---|---|---|
| Anacamptis morio (L.) R.M. Bateman, Pridgeon & M.W. Chase | Species | NT | 1987–2016 |
| Anacamptis pyramidalis (L.) Rich. | Species | VU | 1987–2016 |
| Cephalanthera damasonium (Mill.) Druce | Species | VU | 1987–2016 |
| Cephalanthera longifolia (L.) Fritsch | Species | EN | 1987–2016 |
| Cephalanthera rubra (L.) Rich. | Species | CR | 1987–2016 |
| Coeloglossum viride (L.) Hartm. | Species | RE | |
| Corallorhiza trifida Châtel | Species | VU | 1987–2016 |
| Cypripedium calceolus L. | Species | VU | 1987–2016 |
| Dactylorhiza incarnata (L.) Soó subsp. incarnata | Subspecies | LC | 1987–2016 |
| Dactylorhiza incarnata (L.) Soo subsp. lobelii (Verm.) H.A. Pedersen | Subspecies | VU | |
| Dactylorhiza incarnata (L.) Soó subsp. incarnata var. ochroleuca (Boll) Hyl. | Variety | EN | 1987–2016 |
| Dactylorhiza maculata (L.) Soó subsp. fuchsii (Druce) Hyl. | Subspecies | LC | 1987–2016 |
| Dactylorhiza maculata (L.) Soó subsp. maculata | Subspecies | LC | 1987–2016 |
| Dactylorhiza majalis (Rchb.) P.F. Hunt & Summerh. subsp. calcifugiens H.A. Pedersen | Subspecies | NE | |
| Dactylorhiza majalis (Rchb.) P.F. Hunt & Summerh. subsp. integrata (E.G. Camus) H.A. Pedersen var. integrata | Variety | LC | 1987–2001, 2014–2016 |
| Dactylorhiza majalis (Rchb.) P.F. Hunt & Summerh. subsp. integrata (E.G. Camus) H.A. Pedersen var. junialis (Verm.) H.A. Pedersen | Variety | VU | 1997–2000, 2007–2008 |
| Dactylorhiza majalis (Rchb.) P.F. Hunt & Summerh. subsp. lapponica (Hartm.) H. Sund. | Subspecies | RE | |
| Dactylorhiza majalis (Rchb.) P.F. Hunt & Summerh. subsp. majalis | Subspecies | LC | 1987–2016 |
| Dactylorhiza majalis (Rchb.) P.F. Hunt & Summerh. subsp. purpurella (T. & T.A. Stephenson) D.M. Moore & Soó var. cambrensis (R.H. Roberts) H.A. Pedersen | Variety | LC | 1987–2003 |
| Dactylorhiza majalis (Rchb.) P.F. Hunt & Summerh. subsp. purpurella (T. & T.A. Stephenson) D.M. Moore & Soó var. purpurella | Variety | LC | 1987–2016 |
| Dactylorhiza majalis (Rchb.) P.F. Hunt & Summerh. subsp. sphagnicola (Höppner) H.A. Pedersen & Hedrén | Subspecies | CR | |
| Dactylorhiza sambucina (L.) Soó | Species | EN | 1987–2016 |
| Epipactis atrorubens (Hoffm. ex Bernh.) Besser | Species | NT | 1989–2010 |
| Epipactis helleborine (L.) Crantz subsp. helleborine | Subspecies | LC | 1987–2016 |
| Epipactis helleborine (L.) Crantz subsp. neerlandica (Verm.) Buttler var. neerlandica | Variety | LC | 1990–1993, 2007–2016 |
| Epipactis helleborine (L.) Crantz subsp. neerlandica (Verm.) Buttler var. renzii (Rob.) J.Claess., Kleynen & Wielinga | Variety | NT | |
| Epipactis leptochila (Godfery) Godfery | Species | NT | 1987–2012 |
| Epipactis palustris (L.) Crantz | Species | NT | 1987–2016 |
| Epipactis phyllanthes G.E. Sm. | Species | LC | 1989–2016 |
| Epipactis purpurata Sm. | Species | LC | 1987–2016 |
| Epipogium aphyllum Sw. | Species | CR | 1987–2012 |
| Goodyera repens (L.) R. Br. | Species | LC | 1987–2000, 2010–2016 |
| Gymnadenia conopsea (L.) R. Br. subsp. conopsea | Subspecies | CR | 1987–2016 |
| Gymnadenia conopsea (L.) R. Br. subsp. densiflora (Wahlenb.) K Richt. | Subspecies | CR | 1998–2011, 2015–2016 |
| Hammarbya paludosa (L.) Kuntze | Species | EN | 1987–2016 |
| Herminium monorchis (L.) R. Br. | Species | EN | 1987–2016 |
| Liparis loeselii (L.) Rich. | Species | EN | 1987–2016 |
| Neotinea ustulata (L.) R.M. Bateman, Pridgeon & M.W. Chase | Species | CR | 1987–1991, 1994–2003, 2008–2016 |
| Neottia cordata (L.) Rich. | Species | LC | 1989–2011 |
| Neottia nidus–avis (L.) Rich. | Species | LC | 1987–2016 |
| Neottia ovata (L.) Bluff & Fingerh. | Species | LC | 1987–2016 |
| Ophrys apifera Huds. | Species | VU | 2004–2016 |
| Ophrys insectifera L. | Species | CR | 1987–1995, 2005–2009 |
| Orchis mascula (L.) L. | Species | LC | 1987–2016 |
| Orchis militaris L. | Species | NA | 1987–1994 |
| Orchis purpurea Huds. | Species | NT | 1987–2016 |
| Platanthera bifolia (L.) Rich. subsp. bifolia | Subspecies | NT | 1987–2016 |
| Platanthera bifolia (L.) Rich. subsp. latiflora (Drejer) Løjtnant | Subspecies | EN | 1987–2016 |
| Platanthera chlorantha (Custer) Rchb. | Species | NT | 1987–2016 |
| Pseudorchis albida (L.) À & D. Löve | Species | CR | 1987–2016 |
| Spiranthes spiralis (L.) Chevall. | Species | RE | 1987–1991, 1996–1999 |
| Orchid Name | No. Grid-Squares | No. of Sites | Underground Part | Vegetative Propagation | Fertilisation | Production of Nectar | Pimary Habitat(s) |
|---|---|---|---|---|---|---|---|
| Ana mor | 26 | 34 | Tuber | Formation of more tubers | Cross-fertilization | No | Grassland |
| Ana pyr | (2) 3 | 4 | Tuber | Formation of more tubers | Cross-fertilization | No | Calcareous grassland |
| Cep dam | 20 | 21 | Rhizome | Division of rhizome | Self-fertilization | No | Deciduous woodland on chalk |
| Cep lon | 13 | 13 | Rhizome | Division of rhizome | Cross-fertilization | No | Deciduous woodland on chalk |
| Cep rub | 3 | 5 | Rhizome | Division of rhizome | Cross-fertilization | No | Deciduous woodland on chalk |
| Coe vir | 0 | 0 | Tuber | Formation of more tubers | Cross-fertilization | Yes | Grassland |
| Cor tri | 20 | 21 | Rhizome | Division of rhizome | Cross- and self-fertilization | No | Deciduous woodland on chalk, wooded poor fen |
| Cyp cal | 2 | 2 | Rhizome | Division of rhizome | Cross-fertilization | No | Deciduous woodland on chalk, calcareous grassland |
| Dac inc inc | 338 | 384 | Tuber | Formation of more tubers | Cross-fertilization | No | Rich fen |
| Dac inc lob | 4 | 7 | Tuber | Formation of more tubers | Cross-fertilization | No | Rich fen |
| Dac inc inc och | (4) 3 | 3 | Tuber | Formation of more tubers | Cross-fertilization | No | Rich fen |
| Dac mac fuc | 71 | 130 | Tuber | Formation of more tubers | Cross-fertilization | No | Deciduous woodland |
| Dac mac mac | 268 | 424 | Tuber | Formation of more tubers | Cross-fertilization | No | Grassland, heathland, fen |
| Dac maj cal | 7 | 10 | Tuber | Formation of more tubers | Cross-fertilization | No | Dune slack, rich fen |
| Dac maj int int | 9 | 11 | Tuber | Formation of more tubers | Cross-fertilization | No | Rich fen |
| Dac maj int jun | 4 | 7 | Tuber | Formation of more tubers | Cross-fertilization | No | Rich fen |
| Dac maj lap | 0 | 0 | Tuber | Formation of more tubers | Cross-fertilization | No | Fen |
| Dac maj maj | 556 | 830 | Tuber | Formation of more tubers | Cross-fertilization | No | Rich fen |
| Dac maj pur cam | 12 | 30 | Tuber | Formation of more tubers | Cross-fertilization | No | Dune slack, rich fen |
| Dac ma pur pur | 73 | 60 | Tuber | Formation of more tubers | Cross-fertilization | No | Dune slack, rich fen |
| Dac maj sph | 3 | 3 | Tuber | Formation of more tubers | Cross-fertilization | No | Poor fen |
| Dac sam | 7 | 13 | Tuber | Formation of more tubers | Cross-fertilization | No | Grassland, heathland |
| Epi atr | 2 | 3 | Rhizome | Division of rhizome | Cross-fertilization | Yes | Conifer and deciduous woodland on chalk, grassland |
| Epi hel hel | 677 | 701 | Rhizome | Division of rhizome | Cross-fertilization | Yes | Deciduous woodland, grassland |
| Epi hel nee nee | 19 | 30 | Rhizome | Division of rhizome | Cross-fertilization | Yes | Calcareous dune |
| Epi hel nee ren | 3 | 4 | Rhizome | Division of rhizome | Self-fertilization | No | Calcareous dune |
| Epi lep | 7 | 11 | Rhizome | Division of rhizome | Self-fertilization | No | Deciduous woodland |
| Epi pal | 123 | 231 | Rhizome | Division of rhizome | Cross-fertilization | Yes | Dune slack, rich fen |
| Epi phy | 70 | 119 | Rhizome | Division of rhizome | Self-fertilization | No | Deciduous woodland |
| Epi pur | 23 | 52 | Rhizome | Division of rhizome | Cross-fertilization | Yes | Deciduous woodland |
| Epi aph | 1 | 1 | Rhizome | Division of rhizome | Cross-fertilization | Yes | Deciduous woodland |
| Goo rep | 17 | 18 | Rhizome | Division of rhizome | Cross-fertilization | Yes | Conifer forest |
| Gym con con | (4) 1 | 1 | Tuber | Formation of more tubers | Cross-fertilization | Yes | Rich fen |
| Gym con den | (1) 2 | 2 | Tuber | Formation of more tubers | Cross-fertilization | Yes | Rich fen |
| Ham pal | 18 | 21 | Pseudobulb | Formation of more pseudobulbs and bulbils at leaftips | Cross-fertilization | Yes | Poor fen, blanket bog |
| Her mon | 10 | 18 | Tuber | Formation of more tubers | Cross-fertilization | No | Dune slack, rich fen |
| Lip loe | 15 | 20 | Pseudobulb | Formation of more pseudobulbs | Cross- and self-fertilization | No | Dune slack, rich fen |
| Neoti ust | 2 | 3 | Tuber | Formation of more tubers | Cross-fertilization | No | Calcareous grassland |
| Neo cor | 18 | 21 | Rhizome | Division of rhizome | Cross-fertilization | Yes | Conifer forest, wooded poor fen |
| Neo nid | 119 | 149 | Rhizome | Division of rhizome | Cross- and self-fertilization | No | Deciduous woodland |
| Neo ova | 306 | 495 | Rhizome | Division of rhizome | Cross-fertilization | Yes | Deciduous woodland, grassland, rich fen |
| Oph api | 2 | 5 | Tuber | Formation of more tubers | Self-fertilization | No | Grassland |
| Oph ins | 2 | 1 | Tuber | Formation of more tubers | Cross-fertilization | No | Calcareous grassland |
| Orc mas | 301 | 576 | Tuber | Formation of more tubers | Cross-fertilization | No | Deciduous woodland, grassland |
| Orc mil | (0) 1 | 1 | Tuber | Formation of more tubers | Cross-fertilization | No | Calcareous grassland |
| Orc pur | 9 | 10 | Tuber | Formation of more tubers | Cross-fertilization | No | Deciduous woodland on chalk, calcareous grassland |
| Pla bif bif | 90 | 123 | Tuber | Formation of more tubers | Cross-fertilization | Yes | Grassland, heathland |
| Pla bif lat | 3 | 3 | Tuber | Formation of more tubers | Cross-fertilization | Yes | Deciduous woodland on chalk |
| Pla chl | 231 | 374 | Tuber | Formation of more tubers | Cross-fertilization | Yes | Deciduous woodland, grassland |
| Pse alb | (6) 3 | 3 | Tuber | Formation of more tubers | Cross- and self-fertilization | No | Grassland |
| Spi spi | 0 | 0 | Tuberous root | Division of tuberous roots | Cross-fertilization | Yes | Grassland |
| Species | N | 2.5% | 50% | 97.5% | Model |
|---|---|---|---|---|---|
| Anacamptis morio | 433 | −0.1378 | −0.0715 | −0.007 | 2 |
| Anacamptis pyramidalis | 100 | −0.009 | 0.1233 | 0.242 | 2 |
| Cephalanthera damasonium | 137 | −0.1002 | −0.0102 | 0.0662 | 2 |
| Cephalanthera longifolia | 182 | −0.066 | −0.0265 | 0.0094 | 2 |
| Cephalanthera rubra | 60 | −0.2268 | −0.017 | 0.1976 | 2 |
| Corallorhiza trifida | 189 | −0.4227 | −0.2633 | −0.1144 | 2 |
| Cypripedium calceolus | 83 | −0.1162 | −0.0157 | 0.0746 | 2 |
| Dactylorhiza incarnata subsp. incarnata | 641 | −0.2437 | −0.1155 | 0.0086 | 2 |
| Dactylorhiza maculata subsp. fuchsii | 182 | −0.0851 | −0.0491 | −0.0176 | 2 |
| Dactylorhiza maculata subsp. maculata | 391 | −0.1085 | 0.0153 | 0.1329 | 2 |
| Dactylorhiza majalis subsp. majalis | 928 | −0.0439 | 0.012 | 0.0666 | 2 |
| Dactylorhiza majalis subsp. integrata | 33 | −0.1851 | 0.3126 | 0.8105 | 2 |
| Dactylorhiza majalis subsp. purpurella | 158 | −0.1351 | −0.0363 | 0.058 | 2 |
| Dactylorhiza sambucina | 231 | −0.2581 | −0.1358 | −0.0245 | 2 |
| Epipactis atrorubens | 38 | −0.0823 | −0.0599 | −0.038 | 1 |
| Epipactis helleborine subsp. helleborine | 652 | −0.1575 | −0.1173 | −0.0781 | 2 |
| Epipactis helleborine subsp. neerlandica | 20 | 0.0058 | 0.0228 | 0.0404 | 1 |
| Epipactis leptochila | 217 | −0.1834 | −0.1208 | −0.0608 | 2 |
| Epipactis palustris | 310 | −0.3607 | −0.1382 | 0.0771 | 2 |
| Epipactis phyllanthes | 232 | −0.2395 | −0.1625 | −0.0967 | 2 |
| Epipactis purpurata | 247 | −0.0742 | −0.0352 | 0.0014 | 2 |
| Epipogium aphyllum | 26 | −0.1018 | 0.0223 | 0.1545 | 2 |
| Goodyera repens | 45 | −1.3174 | −0.5082 | 0.2319 | 2 |
| Gymnadenia conopsea subsp. conopsea | 63 | −0.2497 | −0.1098 | −0.0264 | 2 |
| Gymnadenia conopsea subsp. densiflora | 20 | −0.099 | 0.0926 | 0.287 | 2 |
| Hammarbya paludosa | 122 | −0.4299 | −0.2721 | −0.1274 | 2 |
| Herminium monorchis | 131 | −0.3406 | −0.1741 | −0.004 | 2 |
| Liparis loeselii | 408 | −0.124 | −0.0418 | 0.0391 | 2 |
| Neotinea ustulata | 32 | −0.1095 | 0.0433 | 0.1952 | 2 |
| Neottia cordata | 31 | −0.6676 | −0.1139 | 0.4011 | 2 |
| Neottia nidus-avis | 163 | −0.116 | −0.0576 | −0.002 | 2 |
| Neottia ovata | 447 | −0.08 | −0.0411 | −0.0054 | 2 |
| Ophrys apifera | 21 | −0.1546 | 0.7153 | 1.5831 | 2 |
| Ophrys insectifera | 18 | 0 | 0.0344 | 0.0689 | 2 |
| Orchis mascula | 795 | −0.0678 | −0.0361 | −0.005 | 2 |
| Orchis militaris | 6 | −1.9705 | −1.085 | −0.4714 | 2 |
| Orchis purpurea | 140 | −0.0725 | −0.0272 | 0.0208 | 2 |
| Platanthera bifolia subsp. bifolia | 281 | −0.3781 | −0.2153 | −0.074 | 2 |
| Platanthera bifolia subsp. latiflora | 82 | −0.1708 | −0.1162 | −0.0683 | 2 |
| Platanthera chlorantha | 315 | −0.1442 | 0.059 | 0.2744 | 2 |
| Pseudorchis albida | 181 | −0.0725 | −0.0606 | −0.049 | 1 |
| Effect | 2.5% | 50% | 97.5% |
|---|---|---|---|
| Intercept | −0.061 | 0.023 | 0.108 |
| Intensity of livestock grazing | −0.009 | 0.016 | 0.040 |
| Intensity of forest management | −0.031 | 0.006 | 0.043 |
| Overgrowth with tall-growing herbs and shrubs | −0.052 | −0.029 | −0.006 |
| Public disturbance | −0.038 | −0.004 | 0.030 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Damgaard, C.; Moeslund, J.E.; Wind, P. Changes in the Abundance of Danish Orchids over the Past 30 Years. Diversity 2020, 12, 244. https://doi.org/10.3390/d12060244
Damgaard C, Moeslund JE, Wind P. Changes in the Abundance of Danish Orchids over the Past 30 Years. Diversity. 2020; 12(6):244. https://doi.org/10.3390/d12060244
Chicago/Turabian StyleDamgaard, Christian, Jesper Erenskjold Moeslund, and Peter Wind. 2020. "Changes in the Abundance of Danish Orchids over the Past 30 Years" Diversity 12, no. 6: 244. https://doi.org/10.3390/d12060244