Simultaneous Metabarcoding of Eukaryotes and Prokaryotes to Elucidate the Community Structures within Tardigrade Microhabitats


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
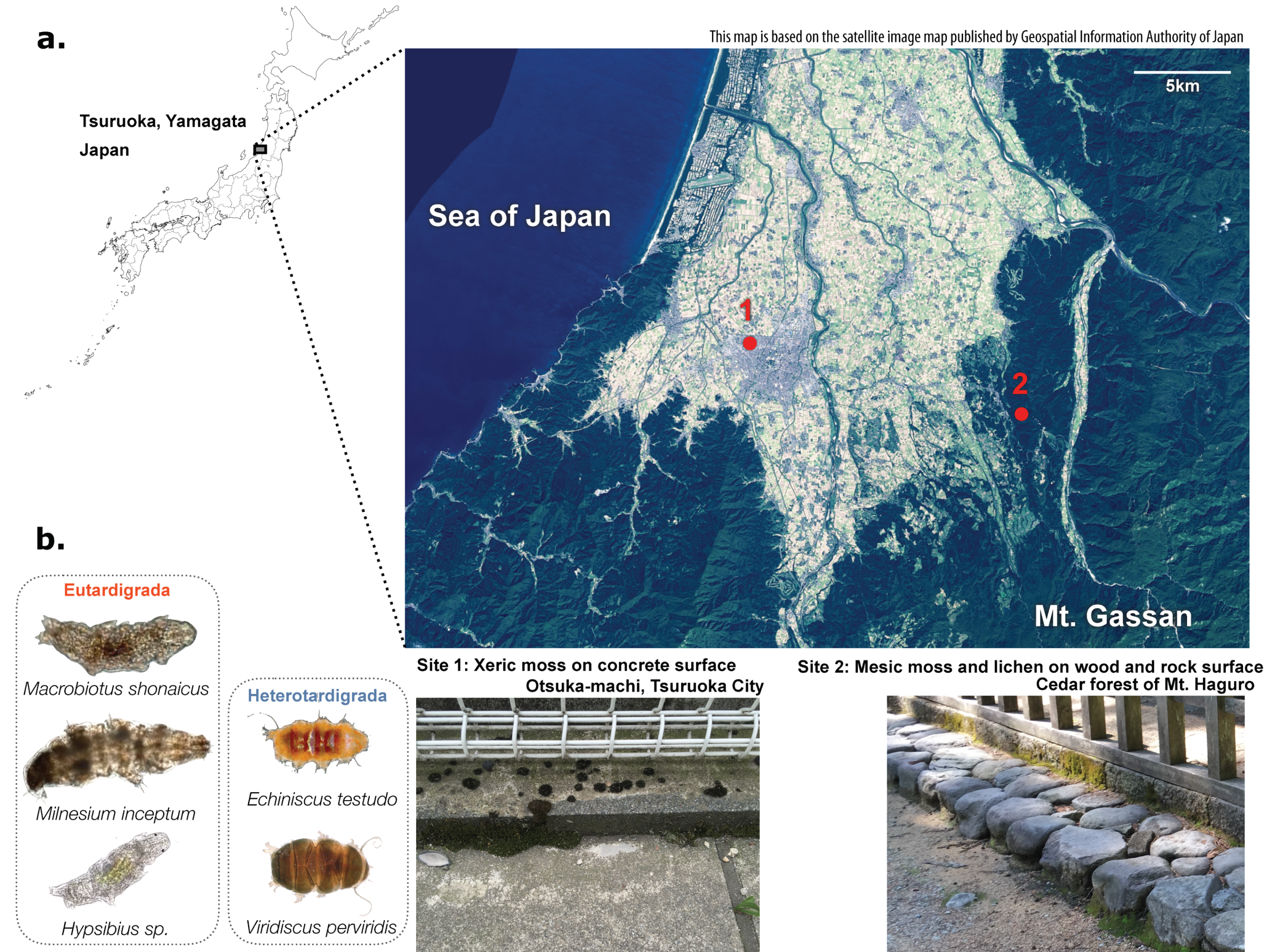
2.1. Sample Collection
2.2. DNA Extraction and PCR Amplification
2.3. Sequencing and Analysis
2.4. Statistics
2.5. Data Availability
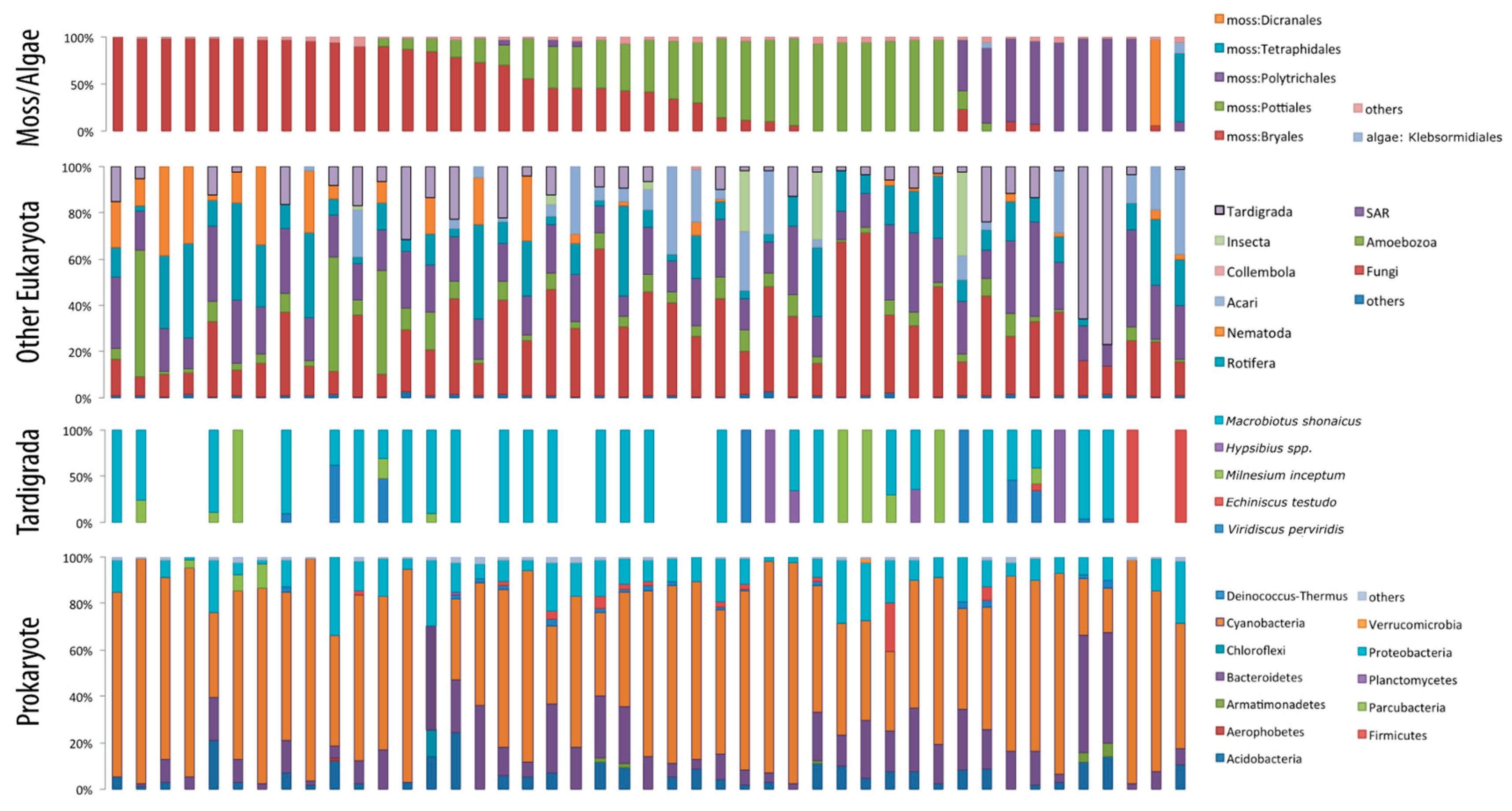
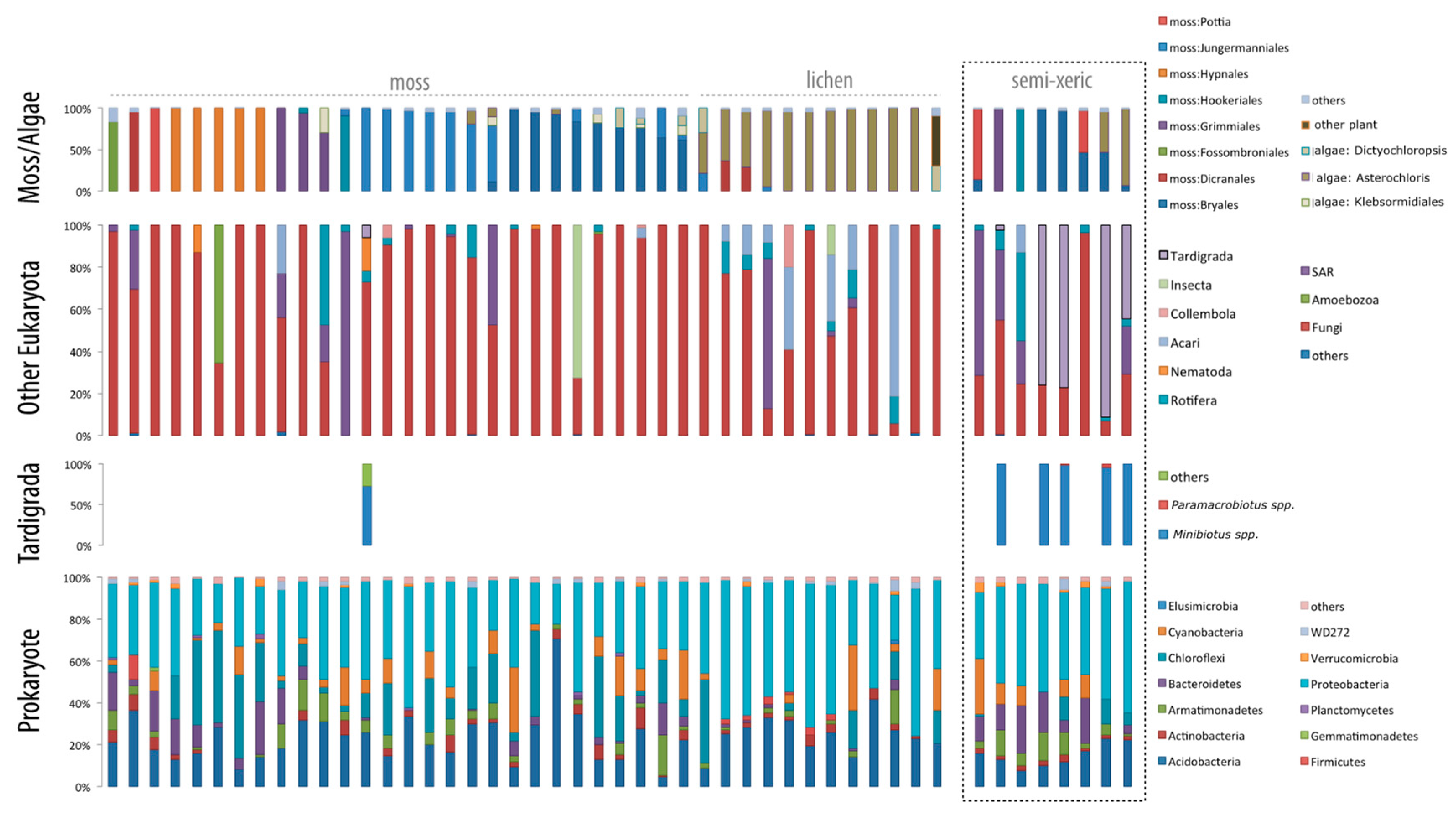
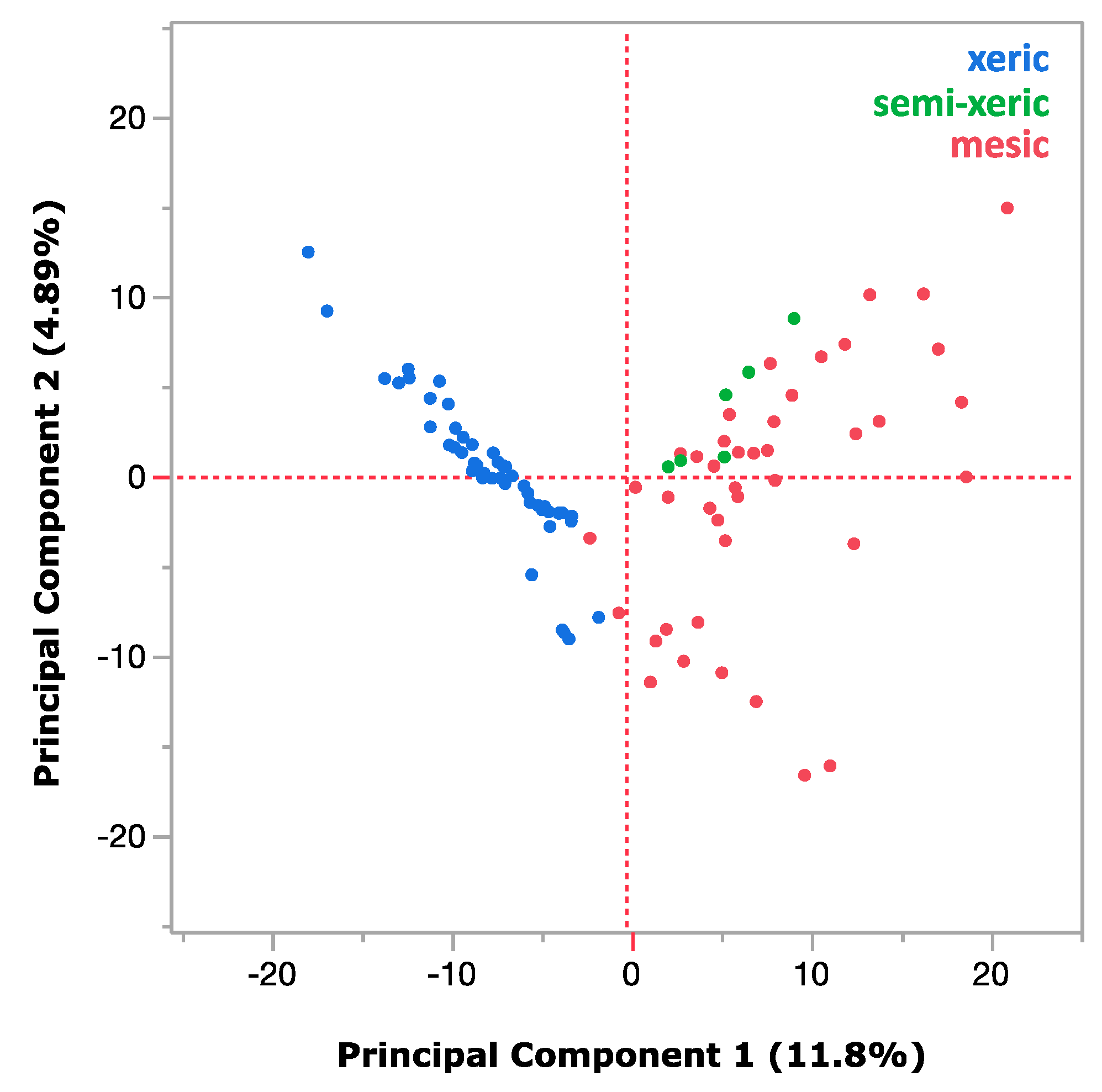
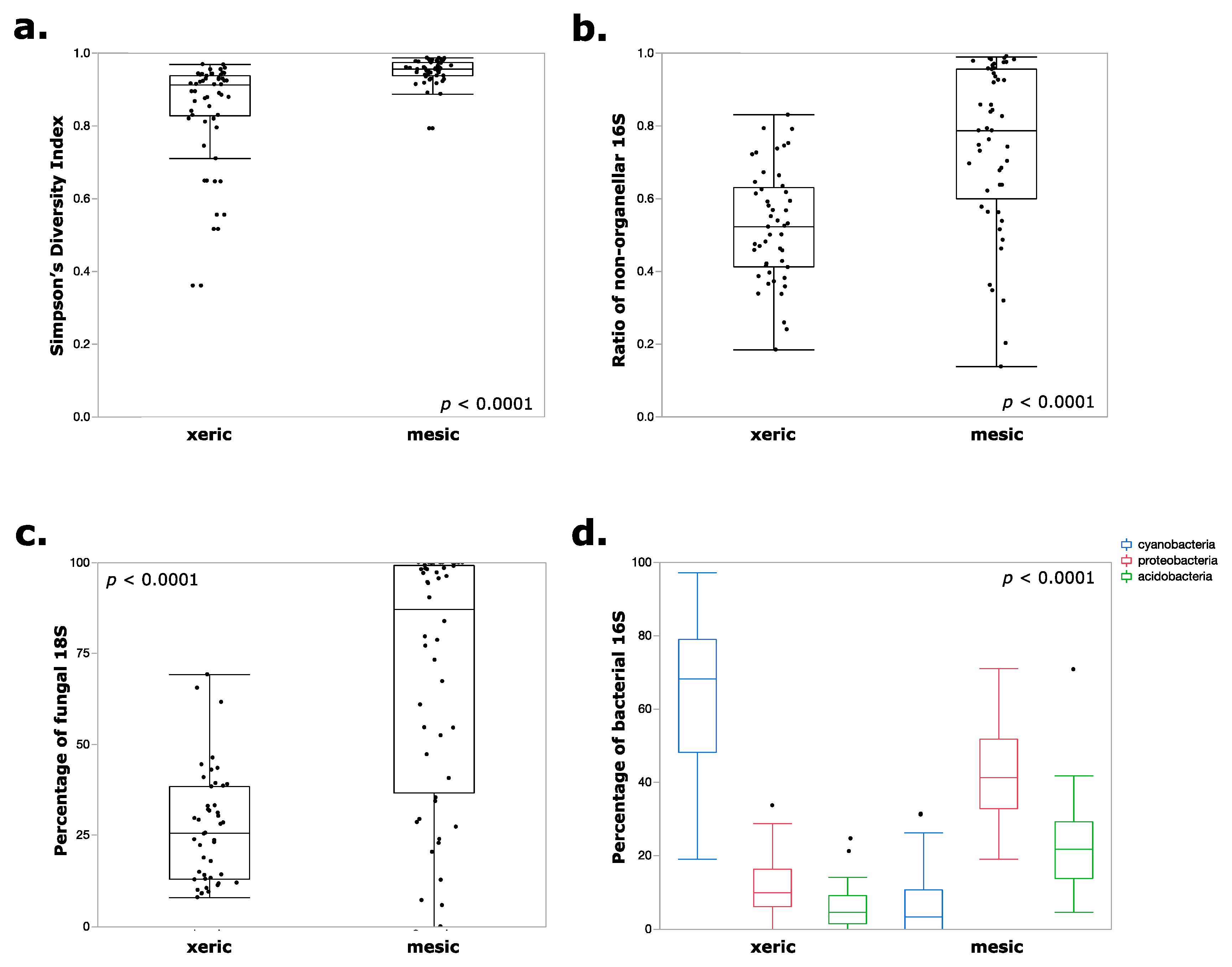
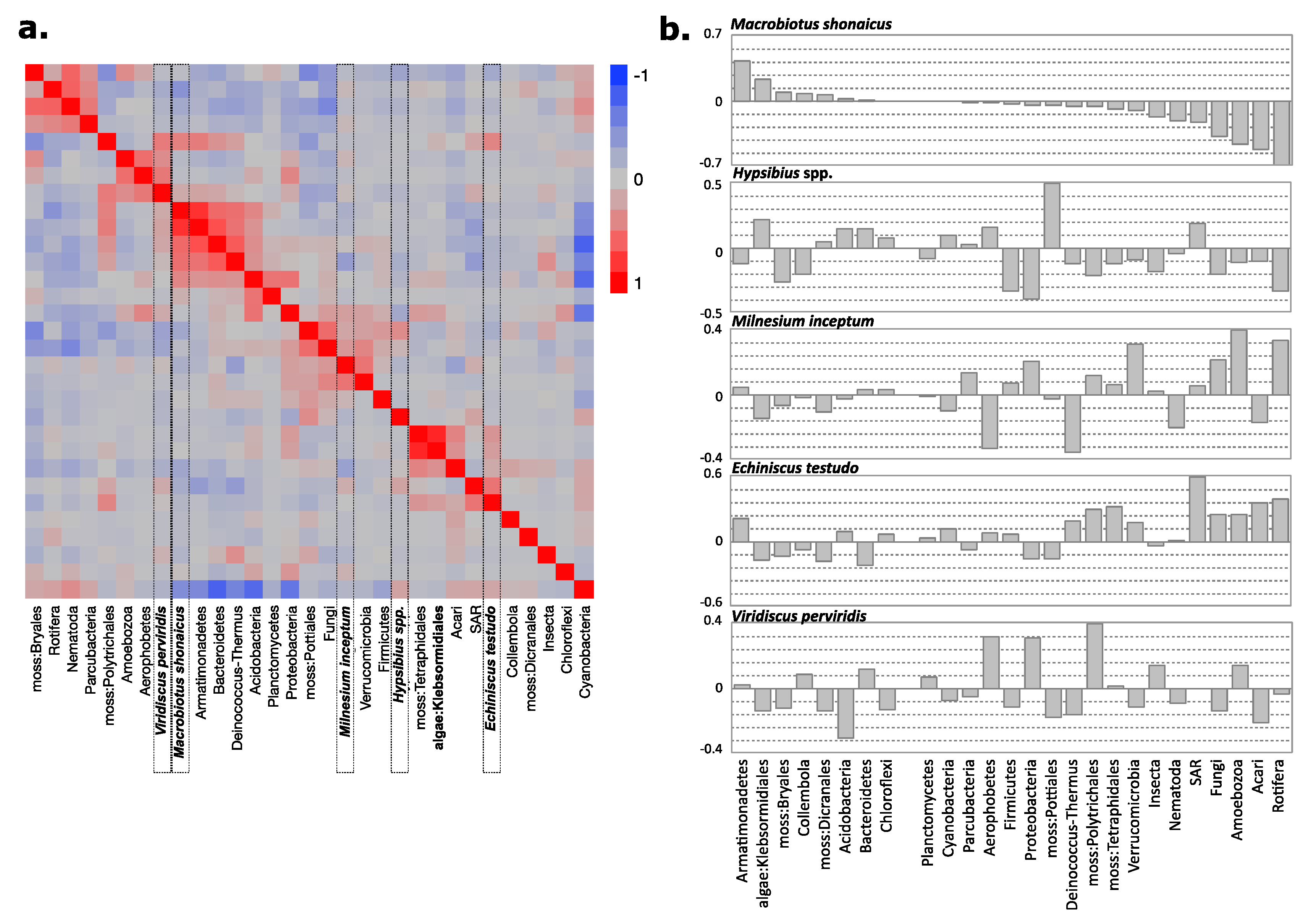
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Funding
Acknowledgments
Conflicts of Interest
References
- Greven, H.; Schuttler, L. How to crawl and dehydrate on moss. Zool. Anz. 2001, 240, 341–344. [Google Scholar] [CrossRef]
- Nelson, D.R.; Adkins, R.G. Distribution of tardigrades within a moss cushion: Do tardigrades migrate in response to changing moisture conditions? Zool. Anz. 2001, 240, 493–500. [Google Scholar] [CrossRef]
- Guidetti, R.; Altiero, T.; Rebecchi, L. On dormancy strategies in tardigrades. J. Insect. Physiol. 2011, 57, 567–576. [Google Scholar] [CrossRef] [PubMed]
- Keilin, D. The Leeuwenhoek Lecture, The problem of anabiosis or latent life: History and current concept. Proc. Royal. Soc. London Series B 1959, 150, 149–191. [Google Scholar]
- Bartels, P.J.; Nelson, D.R. An evaluation of species richness estimators for tardigrades of the Great Smoky Mountains National Park, TN and NC, USA. J. Limnol. 2007, 66, 104–110. [Google Scholar] [CrossRef] [Green Version]
- McInnes, S.J.; Pugh, P.J.A. Tardigrade biogeography. In Water Bears: The Biology of Tardigrades; Schill, R.O., Ed.; Springer: Basel, Switzerland, 2019; pp. 115–129. [Google Scholar]
- Nelson, D.R.; Bartels, P.J.; Fegley, S.R. Environmental correlates of tardigrade community structure in mosses and lichens in the Great Smoky Mountains National Park (Tennessee and North Carolina, USA). Zool. J. Linn. Soc. 2019. [Google Scholar] [CrossRef]
- Kathman, D.; Cross, S. Ecological distribution of moss-dwelling tardigrades on Vancouver Island, British Columbia, Canada. Can. J. Zool. 2011, 69, 122–129. [Google Scholar] [CrossRef]
- Guil, N.; Hortal, J. Effects of macro and micro-environmental factors on the species richness of terrestrial tardigrade assemblages in an Iberian mountain environment. Landsc. Ecol. 2009, 24, 375–390. [Google Scholar] [CrossRef]
- Kaczmarek, Ł.; Goldyn, B.; Welnic, W.; Michalczyk, Ł. Ecological factors determining Tardigrada distribution in Costa Rica. J. Zool. Syst. Evol. Res. 2011, 49, 78–83. [Google Scholar] [CrossRef]
- Schuster, R.; Greven, H. Reproductive traits of Macrobiotus hufelandi during a long-term field study with notes on Paramacrobiotus richtersi and Diphascon pingue (Eutardigrada). J. Limnol. 2013, 72, 166–174. [Google Scholar] [CrossRef] [Green Version]
- Meyer, H. Interspecific association and substrate specificity in tardigrades from Florida, Southeastern United States. Hydrobiologia 2006, 558, 129–132. [Google Scholar] [CrossRef]
- Wright, J.C. The significance of four xeric parameters in the ecology of terrestrial Tardigrada. J. Zool. 1991, 224, 59–77. [Google Scholar] [CrossRef]
- Jönsson, K. Long-term experimental manipulation of moisture conditions and its impact on moss-living tardigrades. J. Limnol. 2007, 66, 119–125. [Google Scholar] [CrossRef] [Green Version]
- Sanchez-Moreno, S.; Ferris, H.; Guil, N. Role of tardigrades in the suppressive service of a soil food web. Agric. Ecosyst. Environ. 2008, 124, 187–192. [Google Scholar] [CrossRef]
- Hyvönen, R.; Persson, T. Effects of fungivorous and predatory arthropods on nematodes and tardigrades in microcosms with coniferous forest soil. Biol. Fertil. Soils 1996, 21, 121–127. [Google Scholar] [CrossRef]
- Altiero, T.; Rebecchi, L. Rearing tardigrades: Results and problems. Zool. Anz. 2001, 240, 217–221. [Google Scholar] [CrossRef]
- Morgan, C.I. Population dynamics of two species of Tardigrada, Macrobiotus hufelandii (Schultze) and Echiniscus testudo (Doyère), in roof moss from Swansea. J. Anim. Ecol. 1977, 46, 263–279. [Google Scholar] [CrossRef]
- Vecchi, M.; Vicente, F.; Guidetti, R.; Bertolani, R.; Rebecchi, L.; Cesari, M. Interspecific relationships of tardigrades with bacteria, fungi and protozoans, with a focus on the phylogenetic position of Pyxidium tardigradum (Ciliophora). Zool. J. Linn. Soc. 2016, 178, 846–855. [Google Scholar] [CrossRef]
- Robeson, M.S., II; Costello, E.K.; Freeman, K.R.; Whiting, J.; Adams, B.; Martin, A.P.; Schmidt, S.K. Environmental DNA sequencing primers for eutardigrades and bdelloid rotifers. BMC Ecol. 2009, 9, 25. [Google Scholar] [CrossRef] [Green Version]
- Vecchi, M.; Newton, I.L.G.; Cesari, M.; Rebecchi, L.; Guidetti, R. The microbial community of tardigrades: Environmental influence and species specificity of microbiome structure and composition. Microb. Ecol. 2018, 76, 467–481. [Google Scholar] [CrossRef]
- Kaczmarek, L.; Roszkowska, M.; Poprawa, I.; Janelt, K.; Kmita, H.; Gawlak, M.; Fialkowska, E.; Mioduchowska, M. Integrative description of bisexual Paramacrobiotus experimentalis sp. nov. (Macrobiotidae) from republic of Madagascar (Africa) with microbiome analysis. Mol. Phylogenet. Evol. 2020, 145, 106730. [Google Scholar] [CrossRef] [PubMed]
- Greenwood, J.L.; Stark, L.R.; Chiquoine, L.P. Effects of rate of drying, life history phase, and ecotype on the ability of the moss Bryum argenteum to survive desiccation events and the influence on conservation and selection of material for restoration. Front. Ecol. Evol. 2019, 7. [Google Scholar] [CrossRef] [Green Version]
- Stec, D.; Arakawa, K.; Michalczyk, L. An integrative description of Macrobiotus shonaicus sp. nov. (Tardigrada: Macrobiotidae) from Japan with notes on its phylogenetic position within the hufelandi group. PLoS ONE 2018, 13, e0192210. [Google Scholar] [CrossRef] [Green Version]
- Gąsiorek, P.; Morek, W.; Stec, D.; Michalczyk, Ł. Untangling the Echiniscus Gordian knot: Paraphyly of the “arctomys group” (Heterotardigrada: Echiniscidae). Cladistics 2019, 35, 633–653. [Google Scholar] [CrossRef]
- Morek, W.; Suzuki, A.C.; Schill, R.O.; Georgiev, D.; Yankova, M.; Marley, N.J.; Michalczyk, Ł. Reinstituting Milnesium alpigenum Ehrenberg, 1853 (Tardigrada: Apochela) after decades of oblivion, with a description of the laboratory model species Milnesium inceptum sp. nov. Zootaxa 2018, 4586, 35–64. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thompson, L.R.; Sanders, J.G.; McDonald, D.; Amir, A.; Ladau, J.; Locey, K.J.; Prill, R.J.; Tripathi, A.; Gibbons, S.M.; Ackermann, G.; et al. A communal catalogue reveals Earth’s multiscale microbial diversity. Nature 2017, 551, 457–463. [Google Scholar] [CrossRef] [Green Version]
- Lundberg, D.S.; Yourstone, S.; Mieczkowski, P.; Jones, C.D.; Dangl, J.L. Practical innovations for high-throughput amplicon sequencing. Nat. Methods 2013, 10, 999–1002. [Google Scholar] [CrossRef]
- Edgar, R.C. UPARSE: Highly accurate OTU sequences from microbial amplicon reads. Nat. Methods 2013, 10, 996–998. [Google Scholar] [CrossRef]
- Yilmaz, P.; Parfrey, L.W.; Yarza, P.; Gerken, J.; Pruesse, E.; Quast, C.; Schweer, T.; Peplies, J.; Ludwig, W.; Glockner, F.O. The SILVA and "All-species Living Tree Project (LTP)" taxonomic frameworks. Nucleic. Acids Res. 2014, 42, D643–D648. [Google Scholar] [CrossRef] [Green Version]
- Leray, M.; Knowlton, N. Censusing marine eukaryotic diversity in the twenty-first century. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2016, 371. [Google Scholar] [CrossRef]
- Gilbert, J.A.; Jansson, J.K.; Knight, R. The Earth Microbiome project: Successes and aspirations. BMC Biol. 2014, 12, 69. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, S.; Xiong, J.; Yu, Y. Taxonomic resolutions based on 18S rRNA genes: A case study of subclass Copepoda. PLoS ONE 2015, 10, e0131498. [Google Scholar] [CrossRef] [PubMed]
- De Vargas, C.; Audic, S.; Henry, N.; Decelle, J.; Mahe, F.; Logares, R.; Lara, E.; Berney, C.; Le Bescot, N.; Probert, I.; et al. Ocean plankton. Eukaryotic plankton diversity in the sunlit ocean. Science 2015, 348, 1261605. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Degma, P.; Katina, S.; Sabatovičová, L. Horizontal distribution of moisture and Tardigrada in a single moss cushion. J. Zool. Syst. Evol. Res. 2011, 49, 71–77. [Google Scholar] [CrossRef]
- Rao, S.; Chan, Y.; Lacap, D.C.; Hyde, K.D.; Pointing, S.B.; Farrell, R.L. Low-diversity fungal assemblage in an Antarctic Dry Valleys soil. Polar Biol. 2012, 35, 567–574. [Google Scholar] [CrossRef] [Green Version]
- Meyer, H.A. Small-scale spatial distribution variability in terrestrial tardigrade populations. Hydrobiologia 2006, 558, 133–139. [Google Scholar] [CrossRef]
- Janssen, P.H. Identifying the dominant soil bacterial taxa in libraries of 16S rRNA and 16S rRNA genes. Appl. Environ. Microbiol. 2006, 72, 1719–1728. [Google Scholar] [CrossRef] [Green Version]
- Rousk, K.; Jones, D.L.; Deluca, T.H. Moss-cyanobacteria associations as biogenic sources of nitrogen in boreal forest ecosystems. Front. Microbiol. 2013, 4, 150. [Google Scholar] [CrossRef] [Green Version]
- Billi, D.; Potts, M. Life and death of dried prokaryotes. Res. Microbiol. 2002, 153, 7–12. [Google Scholar] [CrossRef]
- Singh, H. Desiccation and radiation stress tolerance in cyanobacteria. J. Basic Microbiol. 2018, 58, 813–826. [Google Scholar] [CrossRef]
- Suzuki, A.C. Life history of Milnesium tardigradum Doyère (tardigrada) under a rearing environment. Zool. Sci. 2003, 20, 49–57. [Google Scholar] [CrossRef] [PubMed]






© 2020 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Arakawa, K. Simultaneous Metabarcoding of Eukaryotes and Prokaryotes to Elucidate the Community Structures within Tardigrade Microhabitats. Diversity 2020, 12, 110. https://doi.org/10.3390/d12030110
Arakawa K. Simultaneous Metabarcoding of Eukaryotes and Prokaryotes to Elucidate the Community Structures within Tardigrade Microhabitats. Diversity. 2020; 12(3):110. https://doi.org/10.3390/d12030110
Chicago/Turabian StyleArakawa, Kazuharu. 2020. "Simultaneous Metabarcoding of Eukaryotes and Prokaryotes to Elucidate the Community Structures within Tardigrade Microhabitats" Diversity 12, no. 3: 110. https://doi.org/10.3390/d12030110