
The Influence of Copper and Zinc on Photosynthesis and Phenolic Levels in Basil (Ocimum basilicum L.), Borage (Borago officinalis L.), Common Nettle (Urtica dioica L.) and Peppermint (Mentha piperita L.)
Abstract
:1. Introduction
2. Results
2.1. Soil Analysis
2.2. Plants Analysis
2.2.1. Heavy Metals Uptake by Herbs
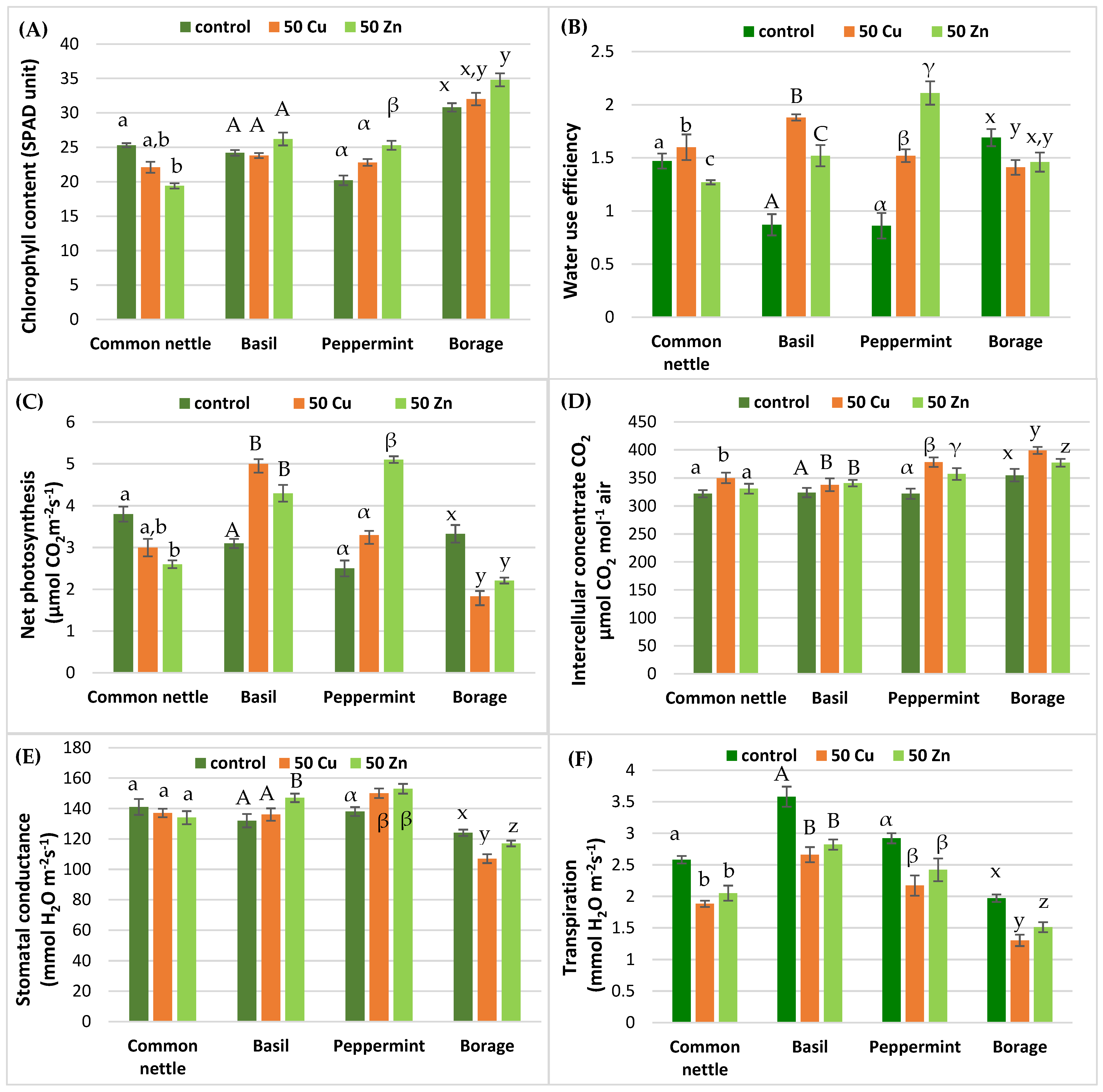
2.2.2. Photosynthesis Parameters
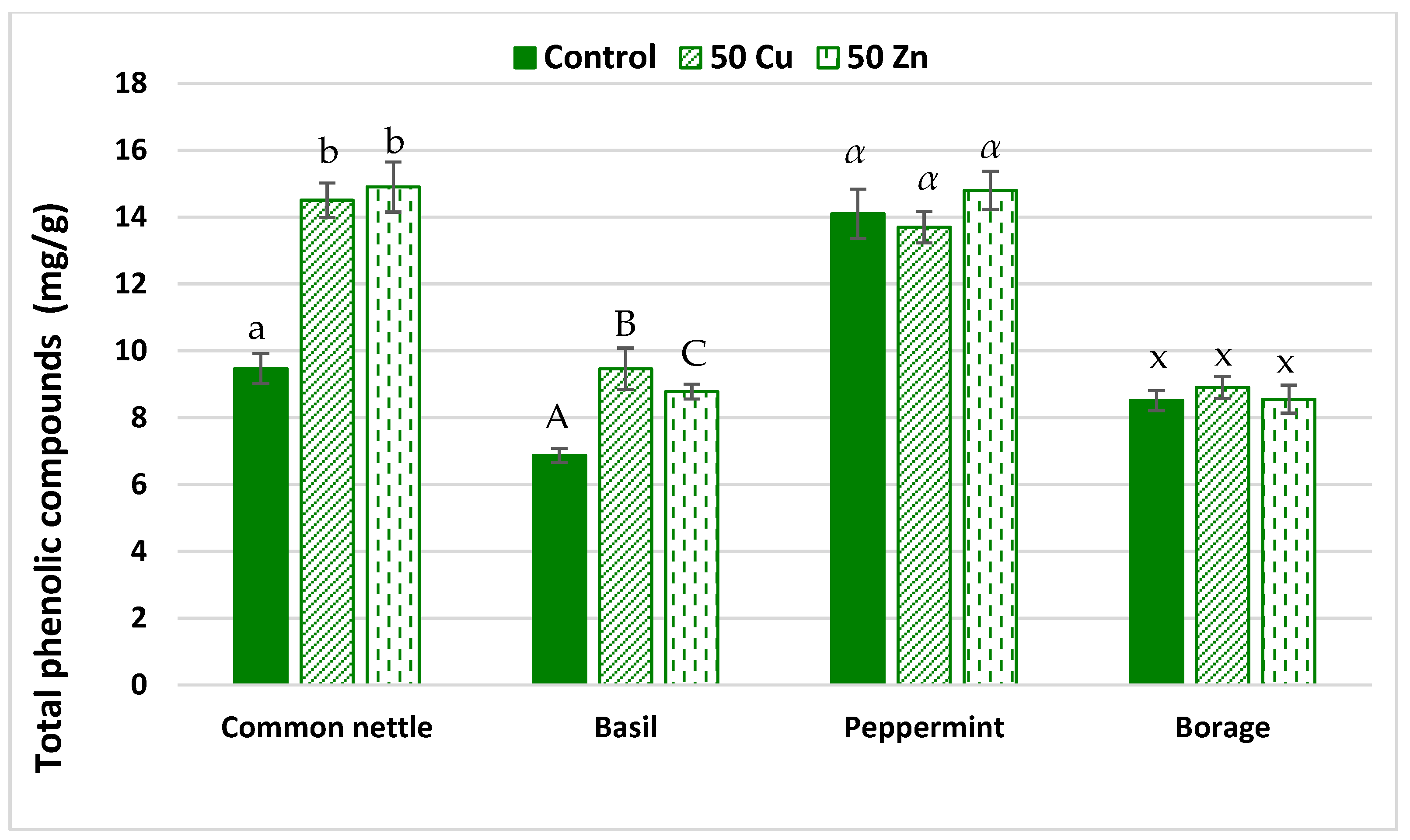
2.2.3. Total Phenolic Compounds Content
3. Discussion
4. Materials and Methods
4.1. Soil Analysis and Preparation for Cultivation
4.2. Plant Material Preparation
4.3. Photosynthesis Parameters
4.4. Determination of Total Phenolic Compounds in Herbs by Folin–Ciocalteu Method
4.5. Determination of Heavy Metals in Herbs
4.6. Data Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Arif, N.; Yadav, V.; Singh, S.; Singh, S.; Ahmad, P.; Mishra, R.K.; Sharma, S.; Tripathi, D.K.; Dubey, N.K.; Chauhan, D.K. Influence of high and low levels of plant-beneficial heavy metal ions on plant growth and development. Front. Environ. Sci. 2016, 4, 69. [Google Scholar] [CrossRef]
- He, H.Y.; Li, Y.; He, L.-F. The central role of hydrogen sulfide in plant responses to toxic metal stress. Ecotoxicol. Environ. Saf. 2018, 157, 403–408. [Google Scholar] [CrossRef]
- Seeda Abou, M.A.; Abou El-Nour, E.A.A.; Yassen, A.A.; Gad, M.M.; Sahar, M.Z. Interaction of copper, zinc, and their importance in plant physiology: Review, Acquisition and Transport. Middle East J. Appl. Sci. 2020, 10, 407–434. [Google Scholar]
- Rizwan, M.; Ali, S.; Maqbool, A. A critical review on the effects of zinc at toxic levels of cadmium in plants. Environ. Sci. Pollut. Res. 2019, 26, 6279–6289. [Google Scholar] [CrossRef]
- Han, G.; Qiao, Z.; Li, Y.; Wang, C.; Wang, B. The roles of CCCH zinc-finger proteins in plant abiotic stress tolerance. Int. J. Mol. Sci. 2021, 22, 8327. [Google Scholar] [CrossRef] [PubMed]
- Alloway, B.J. Soil factors associated with zinc deficiency in crops and humans. Environ. Geochem. Health 2009, 3, 537–548. [Google Scholar] [CrossRef]
- Pietrini, F.; Carnevale, M.; Beni, C.; Zacchini, M.; Gallucci, F.; Santangelo, E. Effect of different copper levels on growth and morpho-physiological parameters in Giant Reed (Arundo donax L.) in semi-hydroponic mesocosm experiment. Water 2019, 11, 1837. [Google Scholar] [CrossRef]
- Badiaa, O.; Yssaad, H.A.R.; Topcuoglu, B. Effect of heavy metals (copper and zinc) on proline, polyphenols and flavonoids content of tomato (Lycopersicon esculentum mill.). Plant Arch. 2020, 20, 2125–2137. [Google Scholar]
- Ninh, A.P.; Xing, G.; Miller, C.J.; Waite, T.D. Fenton-like copper redox chemistry revisited: Hydrogen peroxide and superoxide mediation of copper-catalyzed oxidant production. J. Catal. 2013, 301, 54–64. [Google Scholar]
- Bernal, M.; Ramiro, M.V.; Cases, R.; Picorel, R.; Yruela, I. Excess copper effect on growth, chloroplast ultrastructure, oxygen-evolution activity and chlorophyll fluorescence in Glycine max cell suspensions. Physiol. Plant. 2006, 127, 312–325. [Google Scholar] [CrossRef]
- Shams, M.; Ekinci, M.; Turan, M.; Dursun, A.; Kul, R.; Yildirim, E. Growth, nutrient uptake and enzyme activity response of Lettuce (Lactucasativa L.) to excess copper. Environ. Sustain. 2019, 2, 67–73. [Google Scholar] [CrossRef]
- Giannakoula, A.; Therios, I.; Chatzissavvidis, C. Effect of lead and copper on photosynthetic apparatus in citrus (Citrus aurantium L.) plants. The role of antioxidants in oxidative damage as a response to heavy metal stress. Plants 2021, 10, 155. [Google Scholar] [CrossRef]
- Sahito, S.R.; Memon, M.A.; Kazi, T.G.; Kazi, G.H. Evaluation of mineral contents in medicinal plant Azadirachta indica (Neem). J. Chem. Soc. Pak. 2003, 25, 139–143. [Google Scholar]
- Biezanowska-Kopeć, R.; Piatkowska, E. Total polyphenols and antioxidant properties of selected fresh and dried herbs and spices. Appl. Sci. 2022, 12, 4876. [Google Scholar] [CrossRef]
- Pateiro, M.; Gomez-Salazar, J.A.; Jaime-Patlan, M.; Sosa-Morales, M.E.; Lorenzo, J.M. Plant extracts obtained with green solvents as natural antioxidants in fresh meat products. Antioxidants 2021, 10, 181. [Google Scholar] [CrossRef]
- Adamczyk-Szabela, D.; Lisowska, K.; Romanowska-Duda, Z.; Wolf, W.M. Associated effects of cadmium and copper alter the heavy metals uptake by Melissa Officinalis. Molecules 2019, 24, 2458. [Google Scholar] [CrossRef]
- Adamczyk-Szabela, D.; Lisowska, K.; Romanowska-Duda, Z.; Wolf, W.M. Combined cadmium-zinc interactions alter manganese, lead, copper uptake by Melissa officinalis. Sci. Rep. 2020, 10, 1675. [Google Scholar] [CrossRef]
- Mostafavi, S.; Asadi-Gharneh, H.A.; Miransari, M. The phytochemical variability of fatty acids in basil seeds (Ocimum Basilicum L.) affected by genotype and geographical differences. Food Chem. 2019, 276, 700–706. [Google Scholar] [CrossRef] [PubMed]
- Shahrajabian, M.H.; Wenli, S.; Qi, C. Chemical components and pharmacological benefits of basil (Ocimum basilicum): A review. Int. J. Food Prop. 2020, 23, 1961–1970. [Google Scholar] [CrossRef]
- Javanmardi, J.; Khalighi, A.; Kashi, A.; Bais, H.P.; Vivanco, J.M. Chemical characterization of basil (Ocimum basilicum L.) found in local accessions and used in traditional medicines in Iran. J. Agric. Food Chem. 2002, 50, 5878–5888. [Google Scholar] [CrossRef] [PubMed]
- Mousavi, L.; Salleha, M.R.; Murugaiyah, V. Phytochemical and bioactive compounds identification of Ocimum tenuiflorum leaves of methanol extract and its fraction with an antidiabetic potential. Int. J. Food Prop. 2018, 21, 2390–2399. [Google Scholar] [CrossRef]
- Ahmed, A.F.; Attia, F.A.K.; Liu, Z.; Li, C.; Wei, J.; Kang, W. Antioxidant activity and total phenolic content of essential oils and extracts of sweet basil (Ocimum basilicum L.) plants. Food Sci. Hum. Wellness 2019, 8, 299–305. [Google Scholar] [CrossRef]
- Bulgari, R.; Morgutti, S.; Cocetta, G.; Negrini, N.; Farris, S.; Calcante, A.; Spinardi, A.; Ferrari, E.; Mignani, I.; Oberti, R.; et al. Evaluation of borage extracts as potential biostimulant using a phenomic, agronomic, physiological, and biochemical approach. Front. Plant Sci. 2017, 8, 935. [Google Scholar] [CrossRef]
- Montaner, C.; Zufiaurre, R.; Movila, M.; Mallor, C. Evaluation of borage (Borago officinalis L.) genotypes for nutraceutical value based on leaves fatty acids composition. Foods 2022, 11, 16. [Google Scholar] [CrossRef]
- Gupta, M.; Singh, S. Borago officinalis L.: An important medicinal plant of Mediterranean region: A review. Int. J. Pharm. Sci. Rev. Res. 2010, 5, 27–34. [Google Scholar]
- Sheikhzadeh, P.; Zare, N.; Mahmoudi, F. The synergistic effects of hydro and hormone priming on seed germination, antioxidant activity and cadmium tolerance in borage. Acta Bot. Croat. 2021, 80, 18–28. [Google Scholar] [CrossRef]
- Genc, Z.; Yarat, A.; Tunali-Akbay, T.; Sener, G. The effect of stinging nettle (Urtica dioica) seed oil on experimental colitis in rats. J. Med. Food 2011, 14, 1554–1561. [Google Scholar] [CrossRef]
- Repajic, M.; Cegledi, E.; Zoric, Z.; Pedisic, S.; Elez Garofulic, I.; Radman, S.; Palcic, I.; Dragovic-Uzelac, V. Bioactive compounds in Wild Nettle (Urtica dioica L.) leaves and stalks: Polyphenols and pigments upon seasonal and habitat variations. Foods 2021, 10, 190. [Google Scholar] [CrossRef]
- Beigi, M.; Torki-Harchegani, M.; Pirbalouti, A.G. Quantity and chemical composition of essential oil of Peppermint (Mentha piperita L.) leaves under different drying methods. Int. J. Food Prop. 2018, 1, 267–276. [Google Scholar] [CrossRef]
- Bensabah, F.; Houbairi, S.; Essahli, M.; Lamiri, A.; Naja, J. Chemical composition and inhibitory effect of the essential oil from mentha spicata irrigated by wastewater on the corrosion of aluminum in 1 molar hydrochloric acid. Port. Elect. Chim. Acta 2013, 31, 195–206. [Google Scholar] [CrossRef]
- Zeljkovic, S.C.; Šišková, J.; Komzáková, K.; De Diego, N.; Kaffková, K.; Tarkowski, P. Phenolic compounds and biological activity of selected Mentha Specie. Plants 2021, 10, 550. [Google Scholar] [CrossRef]
- Adamczyk-Szabela, D.; Chrzescijanska, E.; Zielenkiewicz, P.; Wolf, W. Antioxidant activity and photosynthesis efficiency in Melissa officinalis subjected to heavy metals stress. Molecules 2023, 28, 2642. [Google Scholar] [CrossRef]
- He, Z.; Shentu, J.; Yang, X.; Baligar, V.C.; Zhang, T.; Stoffella, P.J. Heavy metal contamination of soils: Sources, indicators, and assessment. J. Environ. Ind. 2015, 9, 17–18. [Google Scholar]
- Regulation of the Minister of Environment. Item 1395, 1 August 2016. Available online: https://isap.sejm.gov.pl/isap.nsf/DocDetails.xsp?id=wdu20160001395 (accessed on 19 March 2024).
- Page, V.; Feller, U. Heavy metals in crop plants: Transport and redistribution processes on the whole plant level. Agronomy 2015, 5, 447–463. [Google Scholar] [CrossRef]
- Skiba, E.; Adamczyk-Szabela, D.; Wolf, W.M. Metal-Based Nanoparticles’ Interactions with Plants, Nanotechnology in the Life Sciences; Springer Nature: Cham, Switzerland, 2021; pp. 145–169. [Google Scholar]
- Behtash, F.; Abedini, F.; Ahmadi, H.; Mosavi, S.B.; Aghaee, A.; Morshedloo, M.R.; Lorenzo, J.M. Zinc application mitigates copper toxicity by regulating cu uptake, activity of antioxidant enzymes, and improving physiological characteristics in summer squash. Antioxidants 2022, 11, 1688. [Google Scholar] [CrossRef]
- Le, T.T.Y.; Vijver, M.G.; Kinraide, T.B.; Peijnenburg, W.J.G.M.; Hendriks, A.J. Modelling metal–metal interactions and metal toxicity to lettuce Lactuca sativa following mixture exposure (Cu2+–Zn2+ and Cu2+–Ag+). Environ. Pollut. 2013, 176, 185–192. [Google Scholar] [CrossRef]
- Andresen, E.; Peiter, E.; Küpper, H. Trace metal metabolism in plants. J. Exp. Bot. 2018, 69, 909–954. [Google Scholar] [CrossRef]
- Walker, E.L.; Connolly, E.L. Time to pump iron: Iron-defciency signaling mechanisms of higher plants. Curr. Opin. Plant Biol. 2008, 11, 530–535. [Google Scholar] [CrossRef]
- Mleczek, M.; Budka, A.; Gąsecka, M.; Budzyńska, S.; Drzewiecka, K.; Magdziak, Z.; Rutkowski, P.; Goliński, P.; Niedzielski, P. Copper, lead and zinc interactions during phytoextraction using Acer platanoides L.: A pot trial. Environ. Sci. Pollut. Res. 2023, 30, 27191–27207. [Google Scholar] [CrossRef]
- Kabata Pendias, A.; Pendias, H. Trace Elements in Soils and Plants, 3rd ed.; CRC Press: Boca Raton, FL, USA; London, UK; New York, NY, USA; Washington, DC, USA, 2001. [Google Scholar]
- Kobayashi, T.; Nozoye, T.; Naoko, K.; Nishizawa, I. Iron transport and its regulation in plants. Free Radic. Biol. Med. 2019, 133, 11–20. [Google Scholar] [CrossRef]
- Ning, X.; Lin, M.; Huang, G.; Mao, J.; Gao, Z.; Wang, X. Research progress on iron absorption, transport, and molecular regulation strategy in plants. Front. Plant Sci. 2023, 14, 1190768. [Google Scholar] [CrossRef]
- Pande, P.; Anwar, M.; Chand, S.; Yadav, V.K.; Patra, D. Optimal level of iron and zinc in relation to its influence on herb yield and production of essential oil in menthol mint. Commun. Soil Sci. Plant Anal. 2007, 38, 561–578. [Google Scholar] [CrossRef]
- Krzesłowska, M.; Timmers, A.C.J.; Mleczek, M.; Niedzielski, P.; Rabęda, I.; Woźny, A.; Goliński, P. Alterations of root architecture and cell wall modifcations in tilia cordata miller (Linden) growing on mining sludge. Environ. Pollut. 2019, 248, 247–259. [Google Scholar] [CrossRef]
- Nasim, S.A.; Dhir, B. Heavy metals alter the potency of medicinal plants. Rev. Environ. Contam. Toxicol. 2010, 203, 139–149. [Google Scholar]
- Dresler, S.; Rutkowska, E.; Bednarek, W.; Stanislawski, G.; Kubrak, T.; Bogucka-Kocka, A.; Wójcik, M. Selected secondary metabolites in Echium vulgare L. populations from nonmetalliferous and metalliferous areas. Phytochemistry 2017, 133, 4–14. [Google Scholar] [CrossRef]
- Clemens, S. Molecular mechanisms of plant metal tolerance and homeostasis. Planta 2001, 212, 475–486. [Google Scholar] [CrossRef]
- Guo, Z.; Gao, Y.; Yuan, X.; Yuan, M.; Huang, L.; Wang, S.; Liu, C.; Duan, C. Effects of heavy metals on stomata in plants: A Review. Int. J. Mol. Sci. 2023, 24, 9302. [Google Scholar] [CrossRef]
- Alkhatib, R.; Mheidat, M.; Abdo, N.; Tadros, M.; Al-Eitan, L.; Al-Hadid, K. Effect of lead on the physiological, biochemical and ultrastructural properties of Leucaena leucocephala. Plant Biol. 2019, 21, 1132–1139. [Google Scholar] [CrossRef]
- Khudsar, T.; Arshi, A.; Siddiqi, T.O.; Mahmooduzzafar; Iqbal, M. Zinc-Induced Changes in Growth Characters, Foliar Properties and Zn-Accumulation Capacity of Pigeon Pea at Different Stages of Plant Growth. J. Plant Nutr. 2008, 31, 281–306. [Google Scholar] [CrossRef]
- Vasilachi, I.C.; Stoleru, V.; Gavrilescu, M. Analysis of heavy metal impacts on cereal crop growth and development in contaminated soils. Agriculture 2023, 13, 1983. [Google Scholar] [CrossRef]
- Xiong, T.; Zhang, T.; Dumat, C.; Sobanska, S.; Dappe, V.; Shahid, M.; Xian, Y.; Li, X.; Li, S. Airborne foliar transfer of particular metals in Lactuca sativa L.: Translocation, phytotoxicity, and bioaccessibility. Environ. Sci. Pollut. Res. Int. 2019, 26, 20064–20078. [Google Scholar] [CrossRef]
- Liao, J.Q.; Li, Z.L.; Wang, J.S.; Tian, D.S.; Tian, D.; Niu, S.L. Nitrogen use efficiency of terrestrial plants in China: Geographic patterns, evolution, and determinants. Ecol. Process. 2021, 10, 69. [Google Scholar] [CrossRef]
- Zeng, X. Water and nutrient use efficiencies of Stipa purpurea Griseb. along a precipitation gradient of the Tibetan plateau. Plant Soil Environ. 2023, 69, 230–237. [Google Scholar] [CrossRef]
- Pandey, A.K.; Zoric, L.; Sun, T.; Karanovic, D.; Fang, P.; Borišev, M.; Wu, X.; Lukovic, J.; Xu, P. The anatomical basis of heavy metal responses in legumes and their impact on plant–rhizosphere interactions. Plants 2022, 11, 2554. [Google Scholar] [CrossRef]
- Rucinska-Sobkowiak, R. Water relations in plants subjected to heavy metal stresses. Acta Physiol. Plant 2016, 38, 257. [Google Scholar] [CrossRef]
- AbdElgawad, H.; Zinta, G.; Hamed, B.A.; Selim, S.; Beemster, G.; Hozzein, W.N.; Wadaan, M.A.M.; Asard, H.; Abuelsoud, W. Maize roots and shoots show distinct profiles of oxidative stress and antioxidant defense under heavy metal toxicity. Environ. Pollut. 2020, 258, 113705. [Google Scholar] [CrossRef]
- Chen, L.; Zhang, D.; Yang, W.; Liu, Y.; Zhang, L.; Gao, S. Sex-specific responses of Populus deltoides to Glomus in traradices colonization and Cd pollution. Chemosphere 2016, 155, 196–206. [Google Scholar] [CrossRef]
- Afzal, S.; Abdul Manap, A.S.; Attiq, A.; Albokhadaim, I.; Kandeel, M.; Alhojaily, S.M. From imbalance to impairment: The central role of reactive oxygen species in oxidative stressinduced disorders and therapeutic exploration. Front. Pharmacol. 2023, 14, 1269581. [Google Scholar] [CrossRef]
- Bhaduri, A.M.; Fulekar, M. Antioxidant enzyme responses of plants to heavy metal stress. Rev. Environ. Sci. Bio/Technol. 2012, 11, 55–69. [Google Scholar] [CrossRef]
- Feng, D.; Wang, R.; Sun, X.; Liu, L.; Liu, P.; Tang, J.; Zhang, C.; Liu, H. Heavy metal stress in plants: Ways to alleviate with exogenous substances. Sci. Total Environ. 2023, 897, 165397. [Google Scholar] [CrossRef]
- Martins, L.L.; Mourato, M.P.; Cardoso, A.I.; Pinto, A.P.; Mota, A.M.; Gonçalves, M.; Plantarum, A. Oxidative stress induced by-cadmium in Nicotiana tabacum L.: Effects on growth parameters, oxidative damage and antioxidant responses in different plant parts. Acta Physiol. Plant. 2011, 33, 1375–1383. [Google Scholar] [CrossRef]
- Chaki, M.; Begara-Morales, J.C.; Barroso, J.B. Oxidative stress in plants. Antioxidants 2020, 9, 481. [Google Scholar] [CrossRef]
- Bertelli, A.; Biagi, M.; Corsini, M.; Baini, G.; Cappellucci, G.; Miraldi, E. Polyphenols: From theory to practice. Foods 2021, 10, 2595. [Google Scholar] [CrossRef]
- Ullah, R.; Hadi, F.; Ahmad, S.; Ullah, A.J.; Rongliang, Q. Phytoremediation of lead and chromium contaminated soil improves with the endogenous phenolics and proline production in Parthenium, Cannabis, Euphorbia, and Rumex Species. Water Air Soil Pollut. 2019, 230, 40. [Google Scholar] [CrossRef]
- Azzazy, M.F. Plant bioindicators of pollution in Sadat City, Western Nile Delta. Egypt PLoS ONE 2020, 15, e0226315. [Google Scholar] [CrossRef]
- PN-ISO 10381-4; Soil Quality—Sampling—Part 4: Rules for Procedure during the Research Areas of Natural, Semi-Natural and Cultivated. International Organization for Standardization: Geneva, Switzerland, 2007.
- PN-ISO 10390; Agricultural Chemical Analysis of the Soil. Determination of pH. International Organization for Standardization: Geneva, Switzerland, 1997. Available online: http://sklep.pkn.pl/pn-iso-10390-1997p.html (accessed on 20 April 2019).
- Schumacher, B.A. Methods for the Determination of Total Organic Carbon (TOC) in Soils and Sediments; Environmental Protection Agency: Washington, DC, USA, 2002. [Google Scholar]
- Ndhlala, A.R.; Kasiyamhuru, A.; Mupure, C.; Chitindingu, K.; Benhura, M.A.; Muchuweti, M. Phenolic composition of Flacourtia indica, Opuntia megacantha and Sclerocarya birrea. Food Chem. 2007, 103, 82–87. [Google Scholar] [CrossRef]
- Goodson, D.Z. Mathematical Methods for Physical and Analytical Chemistry; Wiley: Hoboken, NJ, USA, 2011. [Google Scholar]
- Razali, N.M.; Wah, Y.B. Power comparisons of Shapiro-Wilk, Kolmogorov-Smirnov, Lilliefors and Anderson-Darling tests. J. Stat. Model. Anal. 2011, 2, 21–33. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
| Metals | Bioavailable Forms (µg/g) | Pseudo-Total Forms (µg/g) | Limit Values (µg/g) * |
|---|---|---|---|
| Mn | 75.7 ± 1.0 | 108 ± 0.6 | not applicable |
| Fe | 1323 ± 46 | 2035 ± 190 | not applicable |
| Cu | 9.46 ± 0.06 | 16.7 ± 0.1 | 150 |
| Zn | 61.8 ± 0.8 | 93.3 ± 0.39 | 300 |
| Treatments | Metal Contents in Above-Ground Parts (µg/g) | |||
|---|---|---|---|---|
| Mn | Fe | Cu | Zn | |
| Basil | ||||
| Control | 59.7 ± 3.1 (aa) | 148 ± 11 (ba) | 10.8 ± 1.8 (ca) | 73 ± 5 (da) |
| 50Cu | 66.3 ± 4.7 (aa) | 121 ± 6 (bb) | 47.2 ± 3.1 (cb) | 52 ± 7 (db) |
| 50Zn | 45.1 ± 3.9 (ab) | 104 ± 7 (bc) | 7.61 ± 1.22 (ca) | 191 ± 10 (dc) |
| Borage | ||||
| Control | 52.8 ± 2.5 (ea) | 80.4 ± 6.2 (fa) | 10.1 ± 0.8 (ga) | 56.7 ± 4.8 (ha) |
| 50Cu | 29.8 ± 2.1 (eb) | 71.2 ± 5.1 (fb) | 29.4 ± 1.9 (gb) | 60.2 ± 5.0 (hb) |
| 50Zn | 30.6 ± 2.1 (eb) | 65.4 ± 6.8 (fc) | 8.53 ± 0.88 (gc) | 138 ± 8 (hc) |
| Common nettle | ||||
| Control | 69.7 ± 6.7 (ia) | 103 ± 6 (ja) | 7.42 ± 0.44 (ka) | 29.1 ± 2.8 (la) |
| 50Cu | 58.2 ± 7.1 (ib) | 95 ± 7 (jb) | 28.5 ± 1.9 (kb) | 33.8 ± 2.8 (la) |
| 50Zn | 47.5 ± 6.3 (ic) | 101 ± 7 (ja,jb) | 8.12 ± 0.73 (ka) | 94.6 ± 9.1 (lb) |
| Peppermint | ||||
| Control | 63.3 ± 3.1 (ma) | 115 ± 11 (na) | 7.35 ± 0.72 (oa) | 48.6 ± 3.7 (pa) |
| 50Cu | 49.6 ± 3.7 (mb) | 108 ± 9 (na,nb) | 31.8 ± 5.8 (ob) | 34.2 ± 3.3 (pb) |
| 50Zn | 33.7 ± 2.9 (mc) | 92.6 ± 7 (nb) | 6.39 ± 2.52 (oa) | 105 ± 5 (pc) |
| Treatments | Metal Content in Roots (µg/g) | |||
|---|---|---|---|---|
| Mn | Fe | Cu | Zn | |
| Basil | ||||
| Control | 31.2 ± 2.1 (aa) | 198 ± 6 (ba) | 15.9 ± 1.9 (ca) | 109 ± 6 (da) |
| 50Cu | 70.5 ± 4.3 (ab) | 181 ± 5 (bb) | 113 ± 5 (cb) | 92.1 ± 7.2 (db) |
| 50Zn | 17.1 ± 3.4 (ac) | 138 ± 7 (bc) | 13.2 ± 2.0 (cd) | 287 ± 12 (dc) |
| Borage | ||||
| Control | 40.8 ± 2.5 (ea) | 114 ± 6 (fa) | 11.5 ± 0.8 (ga) | 62.7 ± 4.8 (ha) |
| 50Cu | 32.4 ± 2.1 (eb) | 85.8 ± 5.1 (fb) | 37.4 ± 1.9 (gb) | 70.2 ± 5.0 (hb) |
| 50Zn | 40.6 ± 1.9 (ea) | 75.4 ± 6.8 (fc) | 10.1 ± 0.9 (gc) | 154 ± 8 (hc) |
| Common nettle | ||||
| Control | 49.7 ± 6.7 (ia) | 172 ± 6 (ja) | 9.42 ± 0.44 (ka) | 31.1 ± 2.8 (la) |
| 50Cu | 68.2 ± 7.1 (ib) | 125 ± 7 (jb) | 28.5 ± 1.9 (kb) | 23.8 ± 2.8 (lb) |
| 50Zn | 57.5 ± 6.3 (ic) | 137 ± 7 (jc) | 7.12 ± 0.73 (kc) | 114 ± 9 (lc) |
| Peppermint | ||||
| Control | 71.3 ± 7.1 (ma) | 175 ± 9 (na) | 8.34 ± 0.78 (oa) | 63.6 ± 7.7 (pa) |
| 50Cu | 91.6 ± 8.7 (mb) | 128 ± 7 (nb) | 43.8 ± 5.8 (ob) | 44.2 ± 3.3 (pb) |
| 50Zn | 83.7 ± 2.9 (mc) | 142 ± 8 (nc) | 9.32 ± 2.56 (oa) | 115 ± 6 (pc) |
| Basil | Borage | Common Nettle | Peppermint | |
|---|---|---|---|---|
| Above-Ground Parts | ||||
| Mn | p = 3.85 × 10−6 F = 41.9357 | p = 1.47 × 10−8 F = 115.2513 | p = 4.95 × 10−8 F = 93.0262 | p = 1.70 × 10−9 F = 167.6868 |
| Fe | p = 1.21 × 10−8 F = 119.1513 | p = 2.48 × 10−7 F = 69.7012 | p = 2.02 × 10−2 F = 5.4899 | p = 2.12 × 10−3 F = 10.7391 |
| Cu | p = 6.45 × 10−11 F = 387.6501 | p = 1.52 × 10−9 F = 204.5946 | p = 5.19 × 10−10 F = 254.4199 | p = 5.04 × 10−11 F = 407.4172 |
| Zn | p = 1.17 × 10−11 F = 546.103 | p = 3.21 × 10−9 F = 175.8112 | p = 1.60 × 10−10 F = 322.7082 | p = 1.11 × 10−9 F = 218.2207 |
| Basil | Borage | Common Nettle | Peppermint | |
|---|---|---|---|---|
| Roots | ||||
| Mn | p = 8.23 × 10−13 F = 613.7908 | p = 2.92 × 10−4 F = 17.3007 | p = 1.31 × 10−7 F = 78.1844 | p = 5.46 × 10−7 F = 60.3707 |
| Fe | p = 7.51 × 10−9 F = 129.5797 | p = 2.69 × 10−8 F = 103.6419 | p = 1.14 × 10−9 F = 179.6911 | p = 8.17 × 10−8 F = 85.0893 |
| Cu | p = 5.49 × 10−13 F = 1010.294 | p = 3.28 × 10−8 F = 109.3599 | p = 4.32 × 10−10 F = 264.0011 | p = 1.04 × 10−7 F = 86.1670 |
| Zn | p = 5.72 × 10−11 F = 397.1455 | p = 4.88 × 10−11 F = 409.9500 | p = 7.27 × 10−9 F = 148.8850 | p = 1.17 × 10−7 F = 84.1723 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Adamczyk-Szabela, D.; Wolf, W.M. The Influence of Copper and Zinc on Photosynthesis and Phenolic Levels in Basil (Ocimum basilicum L.), Borage (Borago officinalis L.), Common Nettle (Urtica dioica L.) and Peppermint (Mentha piperita L.). Int. J. Mol. Sci. 2024, 25, 3612. https://doi.org/10.3390/ijms25073612
Adamczyk-Szabela D, Wolf WM. The Influence of Copper and Zinc on Photosynthesis and Phenolic Levels in Basil (Ocimum basilicum L.), Borage (Borago officinalis L.), Common Nettle (Urtica dioica L.) and Peppermint (Mentha piperita L.). International Journal of Molecular Sciences. 2024; 25(7):3612. https://doi.org/10.3390/ijms25073612
Chicago/Turabian StyleAdamczyk-Szabela, Dorota, and Wojciech M. Wolf. 2024. "The Influence of Copper and Zinc on Photosynthesis and Phenolic Levels in Basil (Ocimum basilicum L.), Borage (Borago officinalis L.), Common Nettle (Urtica dioica L.) and Peppermint (Mentha piperita L.)" International Journal of Molecular Sciences 25, no. 7: 3612. https://doi.org/10.3390/ijms25073612