
Early Increase in Blood–Brain Barrier Permeability in a Murine Model Exposed to Fifteen Days of Intermittent Hypoxia
 ,
,
Abstract
:1. Introduction
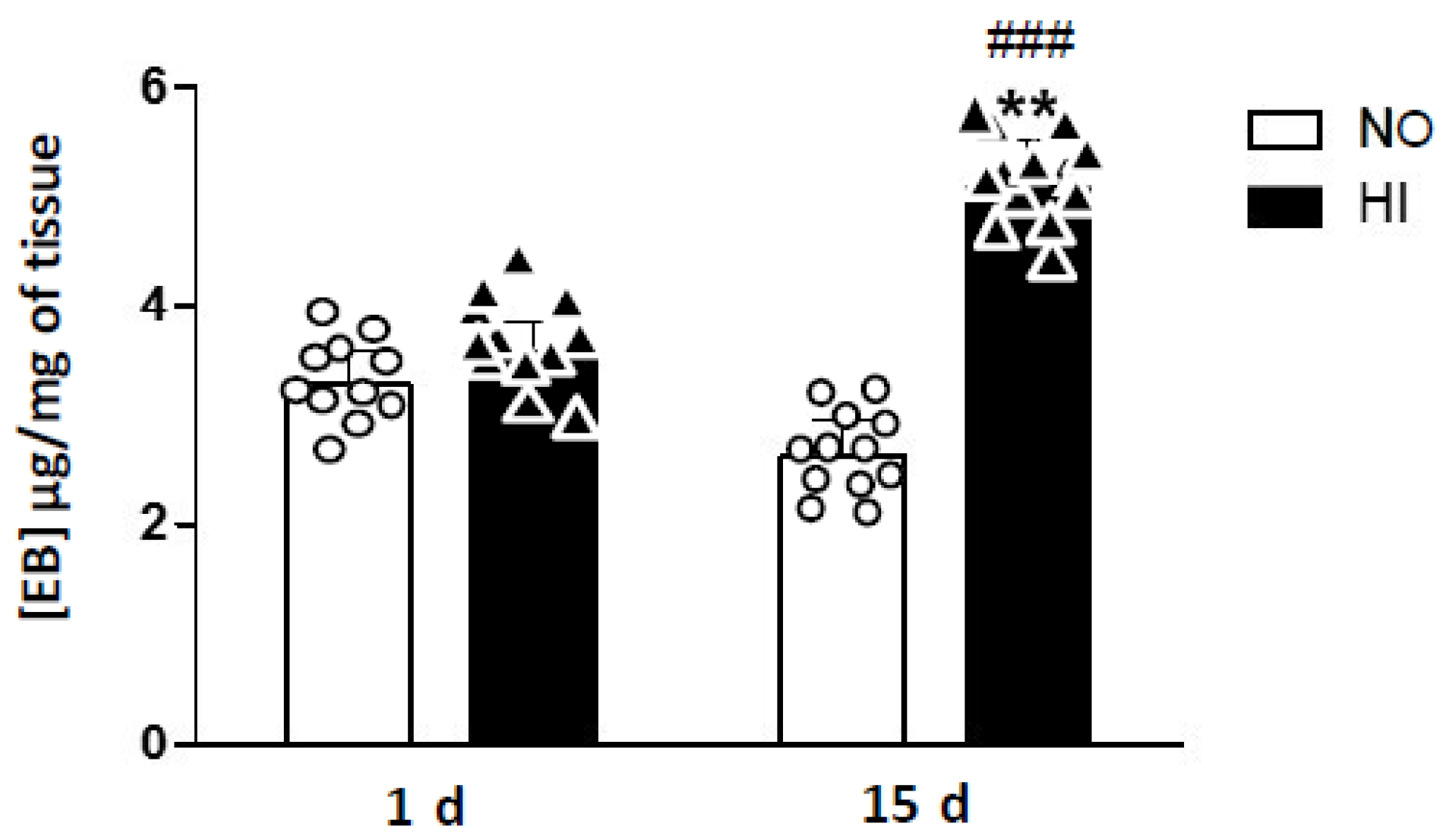
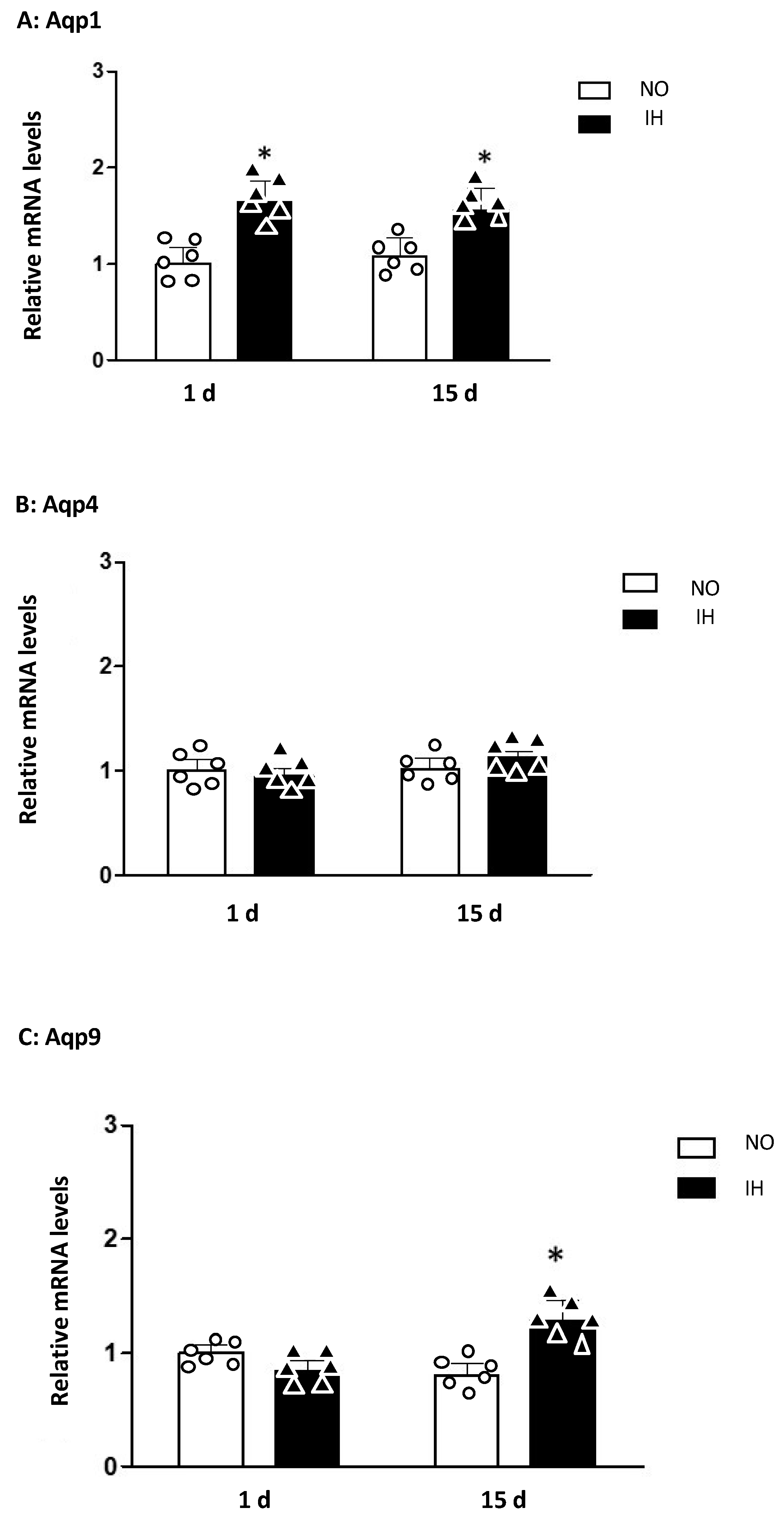
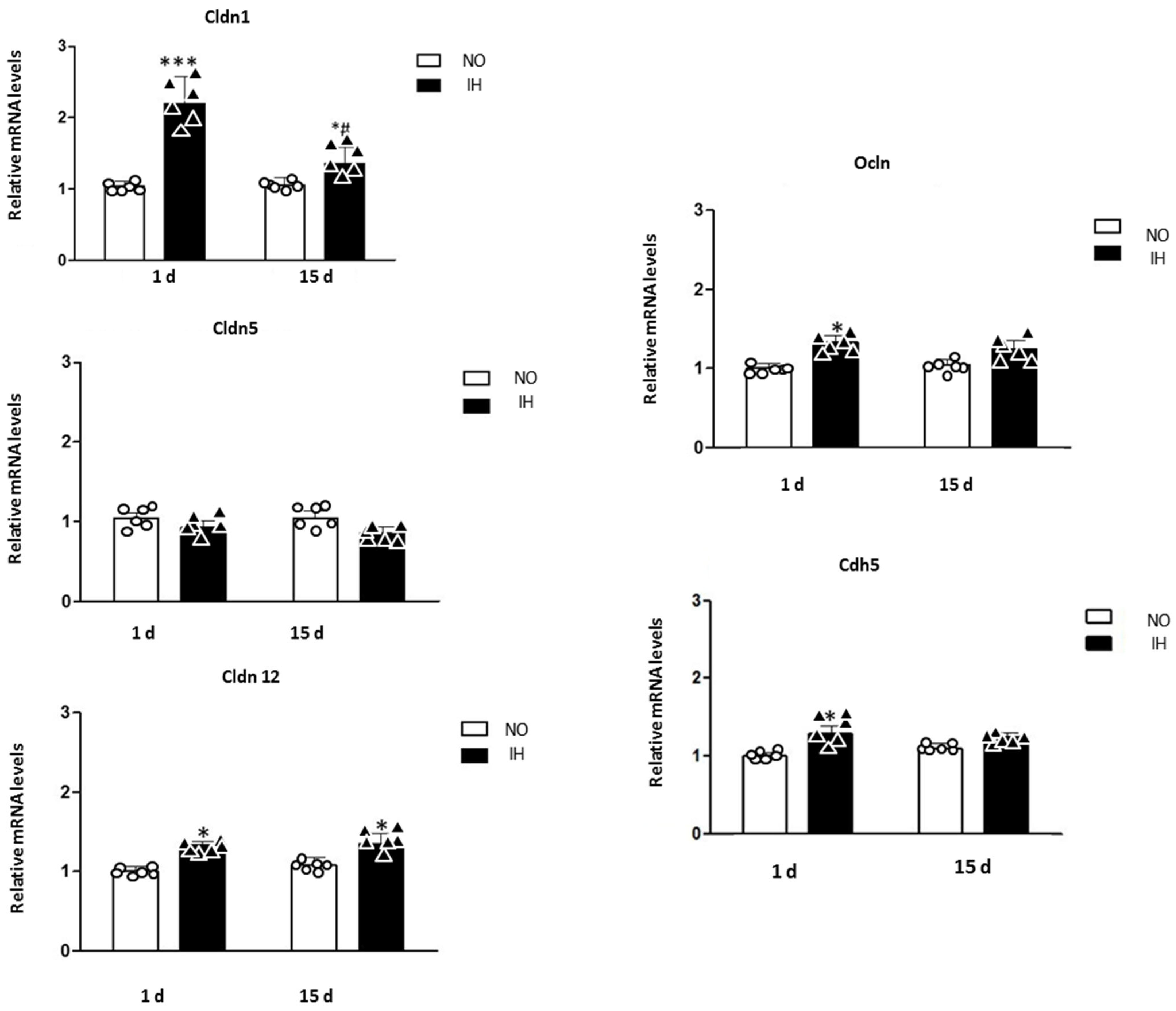
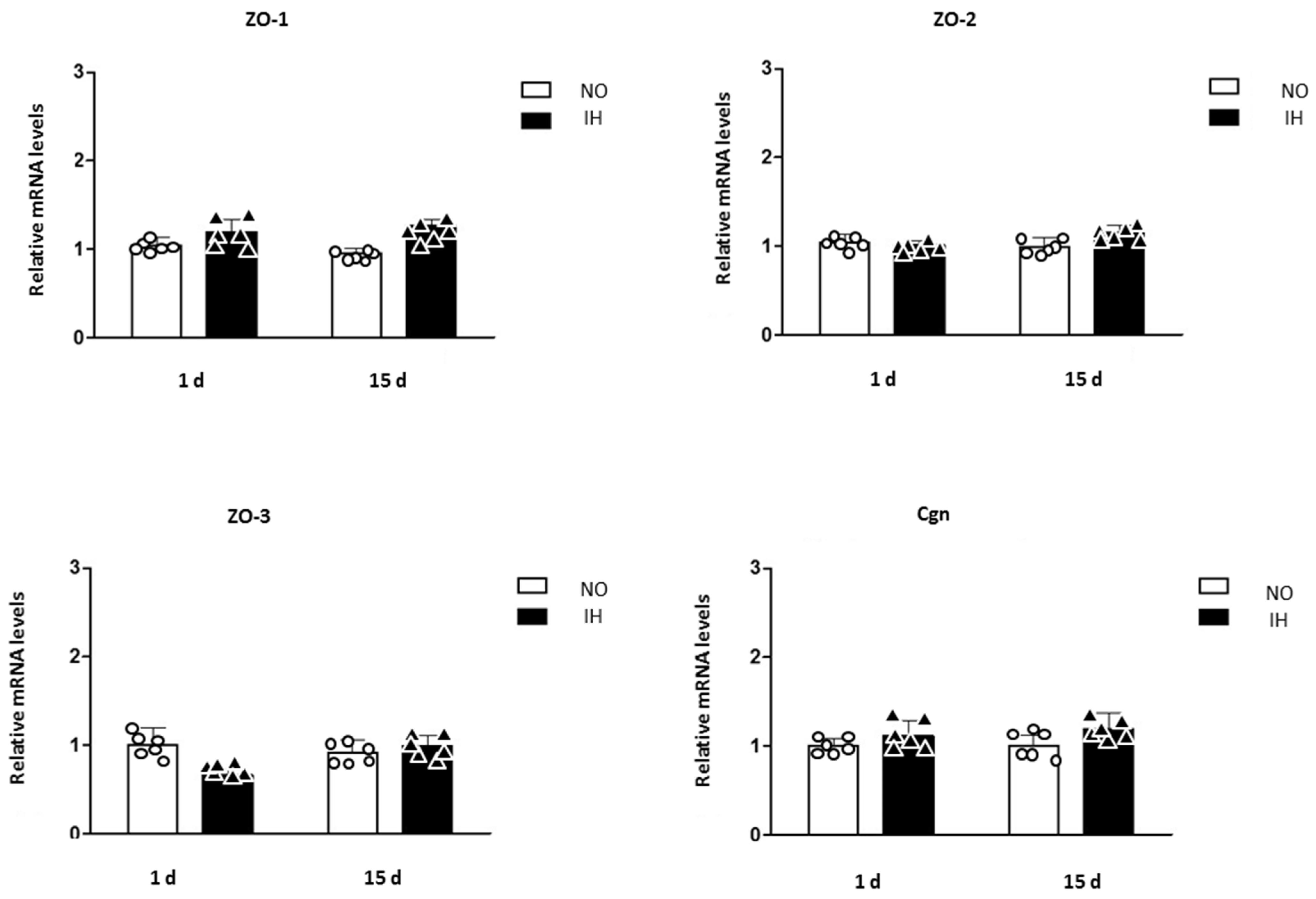
2. Results
3. Discussion
4. Materials and Methods
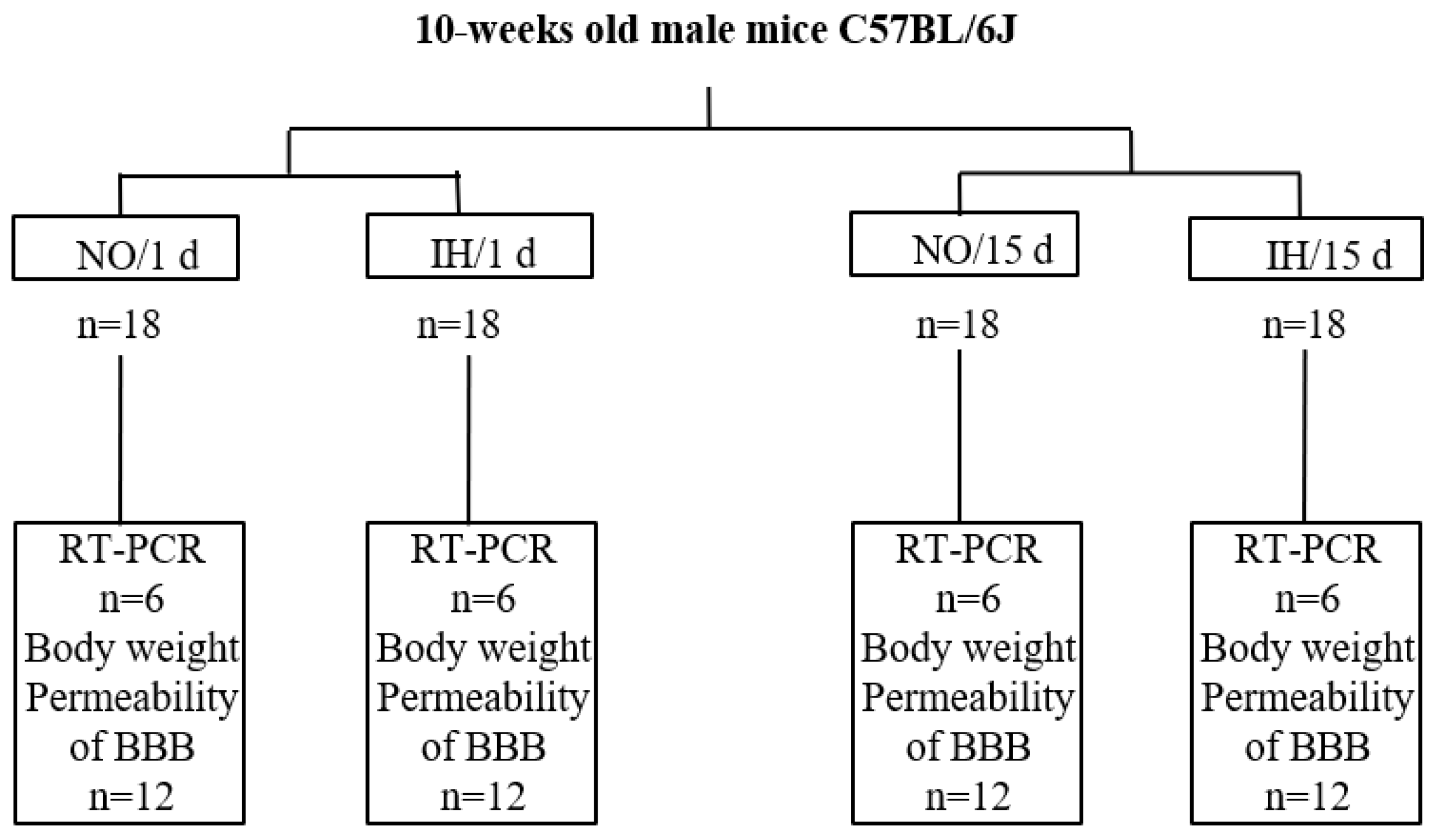
4.1. Murine Model of IH
4.2. Evaluation of BBB Permeability In Vivo
4.3. Relative Measurement of mRNA Expression of BBB Actors by Quantitative RT-PCR
4.4. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| Aqp | Aquaporin |
| BBB | Blood–Brain Barrier |
| Cdh5 | VE-Cadherin |
| Cgn | Cingulin |
| Cldn | Claudin |
| CNS | Central Nervous System |
| IH | Intermittent Hypoxia |
| MRI | Magnetic Resonance Imaging |
| NO | Normoxia |
| Ocln | Occludin |
| RT-PCR | Real-Time Polymerase Chain Reaction |
| ZO | Zonula Occludens |
References
- Al Lawati, N.M.; Patel, S.R.; Ayas, N.T. Epidemiology, Risk Factors, and Consequences of Obstructive Sleep Apnea and Short Sleep Duration. Prog. Cardiovasc. Dis. 2009, 51, 285–293. [Google Scholar] [CrossRef]
- Mehra, R.; Chung, M.K.; Olshansky, B.; Dobrev, D.; Jackson, C.L.; Kundel, V.; Linz, D.; Redeker, N.S.; Redline, S.; Sanders, P.; et al. Sleep-Disordered Breathing and Cardiac Arrhythmias in Adults: Mechanistic Insights and Clinical Implications: A Scientific Statement From the American Heart Association. Circulation 2022, 146, e119–e136. [Google Scholar] [CrossRef]
- Mazzotti, D.R.; Keenan, B.T.; Lim, D.C.; Gottlieb, D.J.; Kim, J.; Pack, A.I. Symptom Subtypes of Obstructive Sleep Apnea Predict Incidence of Cardiovascular Outcomes. Am. J. Respir. Crit. Care Med. 2019, 200, 493–506. [Google Scholar] [CrossRef] [PubMed]
- Kielb, S.A.; Ancoli-Israel, S.; Rebok, G.W.; Spira, A.P. Cognition in Obstructive Sleep Apnea-Hypopnea Syndrome (OSAS): Current Clinical Knowledge and the Impact of Treatment. NeuroMolecular Med. 2012, 14, 180–193. [Google Scholar] [CrossRef] [PubMed]
- Sforza, E.; Haba-Rubio, J.; De Bilbao, F.; Rochat, T.; Ibanez, V. Performance vigilance task and sleepiness in patients with sleep-disordered breathing. Eur. Respir. J. 2004, 24, 279–285. [Google Scholar] [CrossRef] [PubMed]
- Pahari, P.; Korkalainen, H.; Karhu, T.; Rissanen, M.; Arnardottir, E.S.; Hrubos-Strøm, H.; Duce, B.; Töyräs, J.; Leppänen, T.; Nikkonen, S. Obstructive sleep apnea-related intermittent hypoxaemia is associated with impaired vigilance. J. Sleep Res. 2023, 32, e13803. [Google Scholar] [CrossRef] [PubMed]
- Diekelmann, S.; Born, J. The memory function of sleep. Nat. Rev. Neurosci. 2010, 11, 114–126. [Google Scholar] [CrossRef] [PubMed]
- Lin, W.-C.; Huang, C.-C.; Chen, H.-L.; Chou, K.-H.; Chen, P.-C.; Tsai, N.-W.; Chen, M.-H.; Friedman, M.; Lin, H.-C.; Lu, C.-H. Longitudinal Brain Structural Alterations and Systemic Inflammation in Obstructive Sleep Apnea before and after Surgical Treatment. J. Transl. Med. 2016, 14, 139. [Google Scholar] [CrossRef] [PubMed]
- Row, B.W. Intermittent Hypoxia and Cognitive Function: Implications from Chronic Animal Models. Adv. Exp. Med. Biol. 2007, 618, 51–67. [Google Scholar] [CrossRef]
- Lim, D.C.; Pack, A.I. Obstructive Sleep Apnea and Cognitive Impairment: Addressing the Blood-Brain Barrier. Sleep Med. Rev. 2014, 18, 35–48. [Google Scholar] [CrossRef]
- Lim, D.C.; Pack, A.I. Obstructive Sleep Apnea: Update and Future. Annu. Rev. Med. 2017, 68, 99–112. [Google Scholar] [CrossRef]
- Morita, K.; Furuse, M.; Fujimoto, K.; Tsukita, S. Claudin Multigene Family Encoding Four-Transmembrane Domain Protein Components of Tight Junction Strands. Proc. Natl. Acad. Sci. USA 1999, 96, 511–516. [Google Scholar] [CrossRef]
- Greene, C.; Hanley, N.; Campbell, M. Claudin-5: Gatekeeper of Neurological Function. Fluids Barriers CNS 2019, 16, 3. [Google Scholar] [CrossRef]
- Nitta, T.; Hata, M.; Gotoh, S.; Seo, Y.; Sasaki, H.; Hashimoto, N.; Furuse, M.; Tsukita, S. Size-Selective Loosening of the Blood-Brain Barrier in Claudin-5-Deficient Mice. J. Cell Biol. 2003, 161, 653–660. [Google Scholar] [CrossRef]
- Sladojevic, N.; Stamatovic, S.M.; Johnson, A.M.; Choi, J.; Hu, A.; Dithmer, S.; Blasig, I.E.; Keep, R.F.; Andjelkovic, A.V. Claudin-1-Dependent Destabilization of the Blood-Brain Barrier in Chronic Stroke. J. Neurosci. Off. J. Soc. Neurosci. 2019, 39, 743–757. [Google Scholar] [CrossRef] [PubMed]
- Papadopoulos, M.C.; Verkman, A.S. Aquaporin Water Channels in the Nervous System. Nat. Rev. Neurosci. 2013, 14, 265–277. [Google Scholar] [CrossRef]
- Agre, P.; Brown, D.; Nielsen, S. Aquaporin Water Channels: Unanswered Questions and Unresolved Controversies. Curr. Opin. Cell Biol. 1995, 7, 472–483. [Google Scholar] [CrossRef]
- Sugiyama, S.; Sasaki, T.; Tanaka, H.; Yan, H.; Ikegami, T.; Kanki, H.; Nishiyama, K.; Beck, G.; Gon, Y.; Okazaki, S.; et al. The Tight Junction Protein Occludin Modulates Blood-Brain Barrier Integrity and Neurological Function after Ischemic Stroke in Mice. Sci. Rep. 2023, 13, 2892. [Google Scholar] [CrossRef] [PubMed]
- Günzel, D.; Yu, A.S.L. Claudins and the Modulation of Tight Junction Permeability. Physiol. Rev. 2013, 93, 525–569. [Google Scholar] [CrossRef]
- Itoh, M.; Furuse, M.; Morita, K.; Kubota, K.; Saitou, M.; Tsukita, S. Direct Binding of Three Tight Junction-Associated MAGUKs, ZO-1, ZO-2, and ZO-3, with the COOH Termini of Claudins. J. Cell Biol. 1999, 147, 1351–1363. [Google Scholar] [CrossRef] [PubMed]
- Payen, J.F.; Fauvage, B.; Falcon, D.; Lavagne, P. [Brain oedema following blood-brain barrier disruption: Mechanisms and diagnosis]. Ann. Fr. Anesth. Reanim. 2003, 22, 220–225. [Google Scholar] [CrossRef]
- Tran, N.D.; Kim, S.; Vincent, H.K.; Rodriguez, A.; Hinton, D.R.; Bullock, M.R.; Young, H.F. Aquaporin-1-Mediated Cerebral Edema Following Traumatic Brain Injury: Effects of Acidosis and Corticosteroid Administration. J. Neurosurg. 2010, 112, 1095–1104. [Google Scholar] [CrossRef]
- Baronio, D.; Martinez, D.; Fiori, C.Z.; Bambini-Junior, V.; Forgiarini, L.F.; Pase da Rosa, D.; Kim, L.J.; Cerski, M.R. Altered Aquaporins in the Brains of Mice Submitted to Intermittent Hypoxia Model of Sleep Apnea. Respir. Physiol. Neurobiol. 2013, 185, 217–221. [Google Scholar] [CrossRef]
- Sapin, E.; Peyron, C.; Roche, F.; Gay, N.; Carcenac, C.; Savasta, M.; Levy, P.; Dematteis, M. Chronic Intermittent Hypoxia Induces Chronic Low-Grade Neuroinflammation in the Dorsal Hippocampus of Mice. Sleep 2015, 38, 1537–1546. [Google Scholar] [CrossRef]
- Sweeney, M.D.; Zhao, Z.; Montagne, A.; Nelson, A.R.; Zlokovic, B.V. Blood-Brain Barrier: From Physiology to Disease and Back. Physiol. Rev. 2019, 99, 21–78. [Google Scholar] [CrossRef]
- Kim, L.J.; Martinez, D.; Fiori, C.Z.; Baronio, D.; Kretzmann, N.A.; Barros, H.M.T. Hypomyelination, Memory Impairment, and Blood-Brain Barrier Permeability in a Model of Sleep Apnea. Brain Res. 2015, 1597, 28–36. [Google Scholar] [CrossRef]
- Pfeiffer, F.; Schäfer, J.; Lyck, R.; Makrides, V.; Brunner, S.; Schaeren-Wiemers, N.; Deutsch, U.; Engelhardt, B. Claudin-1 induced sealing of blood-brain barrier tight junctions ameliorates chronic experimental autoimmune encephalomyelitis. Acta Neuropathol. 2011, 122, 601–614. [Google Scholar] [CrossRef]
- Agre, P.; Kozono, D. Aquaporin Water Channels: Molecular Mechanisms for Human Diseases. FEBS Lett. 2003, 555, 72–78. [Google Scholar] [CrossRef] [PubMed]
- Saadoun, S.; Papadopoulos, M.C.; Davies, D.C.; Bell, B.A.; Krishna, S. Increased Aquaporin 1 Water Channel Expression in Human Brain Tumours. Br. J. Cancer 2002, 87, 621–623. [Google Scholar] [CrossRef] [PubMed]
- Mobasheri, A.; Marples, D. Expression of the AQP-1 Water Channel in Normal Human Tissues: A Semiquantitative Study Using Tissue Microarray Technology. Am. J. Physiol. Cell Physiol. 2004, 286, C529–C537. [Google Scholar] [CrossRef] [PubMed]
- Blixt, J.; Svensson, M.; Gunnarson, E.; Wanecek, M. Aquaporins and Blood-Brain Barrier Permeability in Early Edema Development after Traumatic Brain Injury. Brain Res. 2015, 1611, 18–28. [Google Scholar] [CrossRef] [PubMed]
- Katada, R.; Akdemir, G.; Asavapanumas, N.; Ratelade, J.; Zhang, H.; Verkman, A.S. Greatly Improved Survival and Neuroprotection in Aquaporin-4-Knockout Mice Following Global Cerebral Ischemia. FASEB J. Off. Publ. Fed. Am. Soc. Exp. Biol. 2014, 28, 705–714. [Google Scholar] [CrossRef] [PubMed]
- Castro Dias, M.; Coisne, C.; Baden, P.; Enzmann, G.; Garrett, L.; Becker, L.; Hölter, S.M.; Hrabě de Angelis, M.; Deutsch, U.; Engelhardt, B. Claudin-12 Is Not Required for Blood-Brain Barrier Tight Junction Function. Fluids Barriers CNS 2019, 16, 30. [Google Scholar] [CrossRef] [PubMed]
- Chatard, M.; Puech, C.; Roche, F.; Perek, N. Hypoxic Stress Induced by Hydralazine Leads to a Loss of Blood-Brain Barrier Integrity and an Increase in Efflux Transporter Activity. PLoS ONE 2016, 11, e0158010. [Google Scholar] [CrossRef] [PubMed]
- Voirin, A.C.; Chatard, M.; Briançon-Marjollet, A.; Pepin, J.L.; Perek, N.; Roche, F. Loss of Blood-Brain Barrier Integrity in an In Vitro Model Subjected to Intermittent Hypoxia: Is Reversion Possible with a HIF-1α Pathway Inhibitor? Int. J. Mol. Sci. 2023, 24, 5062. [Google Scholar] [CrossRef] [PubMed]
- Chatard, M.; Puech, C.; Perek, N.; Roche, F. Hydralazine Is a Suitable Mimetic Agent of Hypoxia to Study the Impact of Hypoxic Stress on In Vitro Blood-Brain Barrier Model. Cell. Physiol. Biochem. Int. J. Exp. Cell. Physiol. Biochem. Pharmacol. 2017, 42, 1592–1602. [Google Scholar] [CrossRef]
- Zolotoff, C.; Voirin, A.-C.; Puech, C.; Roche, F.; Perek, N. Intermittent Hypoxia and Its Impact on Nrf2/HIF-1α Expression and ABC Transporters: An in Vitro Human Blood-Brain Barrier Model Study. Cell. Physiol. Biochem. Int. J. Exp. Cell. Physiol. Biochem. Pharmacol. 2020, 54, 1231–1248. [Google Scholar] [CrossRef]
- Voirin, A.-C.; Celle, S.; Perek, N.; Roche, F. Sera of Elderly Obstructive Sleep Apnea Patients Alter Blood-Brain Barrier Integrity in Vitro: A Pilot Study. Sci. Rep. 2020, 10, 11309. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Sequence 5′-3′ | |
|---|---|---|
| Aqp 1 | Forward | CTGCTGGCGATTGACTACACTGG |
| Reverse | CCCCACCCAGAAAATCCAGTG | |
| Aqp 4 | Forward | GGTGGGGTAAGTGTGGACATTCC |
| Reverse | AGCGTGGCCAGAAATTCTGCT | |
| Aqp 9 | Forward | CATCACGGGAGAAAATGGAACG |
| Reverse | GAGGAACATGGTAGACACCACTTGG | |
| Cldn 1 | Forward | GTTTGCAGAGACCCCATCAC |
| Reverse | AGAAGCCAGGATGAAACCCA | |
| Cldn 5 | Forward | TCTGCTGGTTCGCCAACAT |
| Reverse | CGGCACCGTCGGATCA | |
| Cldn 12 | Forward | ATCACATTCAACAGAAACGAG |
| Reverse | GGTCGTACATCAGGCAGTCA | |
| ZO-1 | Forward | TTAAGGACAAATTCCGCAGGCTAC |
| Reverse | TCTCAGCCTTCTTGATCAGTGCG | |
| ZO-2 | Forward | GATAGCAGCCATCGTGGTCAAGA |
| Reverse | GCGGAAACTTCTGCCATCAAATT | |
| ZO-3 | Forward | CCAGTTTTAAGCGCCCCGTG |
| Reverse | AGTCCGGGACATGCTTTCTGC | |
| Cingulin | Forward | GCAGTCCACCAATCGCAAACTAG |
| Reverse | GCCTTCACCCTCAGGGTTAGCT | |
| Cadherin 5 | Forward | GTACAGCATCATGCAGGGCGA |
| Reverse | CTTCATAGTCCAGGGACTTCGTGG |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Roche, F.; Briançon-Marjollet, A.; Dematteis, M.; Baldazza, M.; Gonthier, B.; Bertholon, F.; Perek, N.; Pépin, J.-L. Early Increase in Blood–Brain Barrier Permeability in a Murine Model Exposed to Fifteen Days of Intermittent Hypoxia. Int. J. Mol. Sci. 2024, 25, 3065. https://doi.org/10.3390/ijms25053065
Roche F, Briançon-Marjollet A, Dematteis M, Baldazza M, Gonthier B, Bertholon F, Perek N, Pépin J-L. Early Increase in Blood–Brain Barrier Permeability in a Murine Model Exposed to Fifteen Days of Intermittent Hypoxia. International Journal of Molecular Sciences. 2024; 25(5):3065. https://doi.org/10.3390/ijms25053065
Chicago/Turabian StyleRoche, Frederic, Anne Briançon-Marjollet, Maurice Dematteis, Marie Baldazza, Brigitte Gonthier, Frederique Bertholon, Nathalie Perek, and Jean-Louis Pépin. 2024. "Early Increase in Blood–Brain Barrier Permeability in a Murine Model Exposed to Fifteen Days of Intermittent Hypoxia" International Journal of Molecular Sciences 25, no. 5: 3065. https://doi.org/10.3390/ijms25053065