
Deciphering the Interrelationship of arnT Involved in Lipid-A Alteration with the Virulence of Salmonella Typhimurium


Abstract
:1. Introduction
2. Results
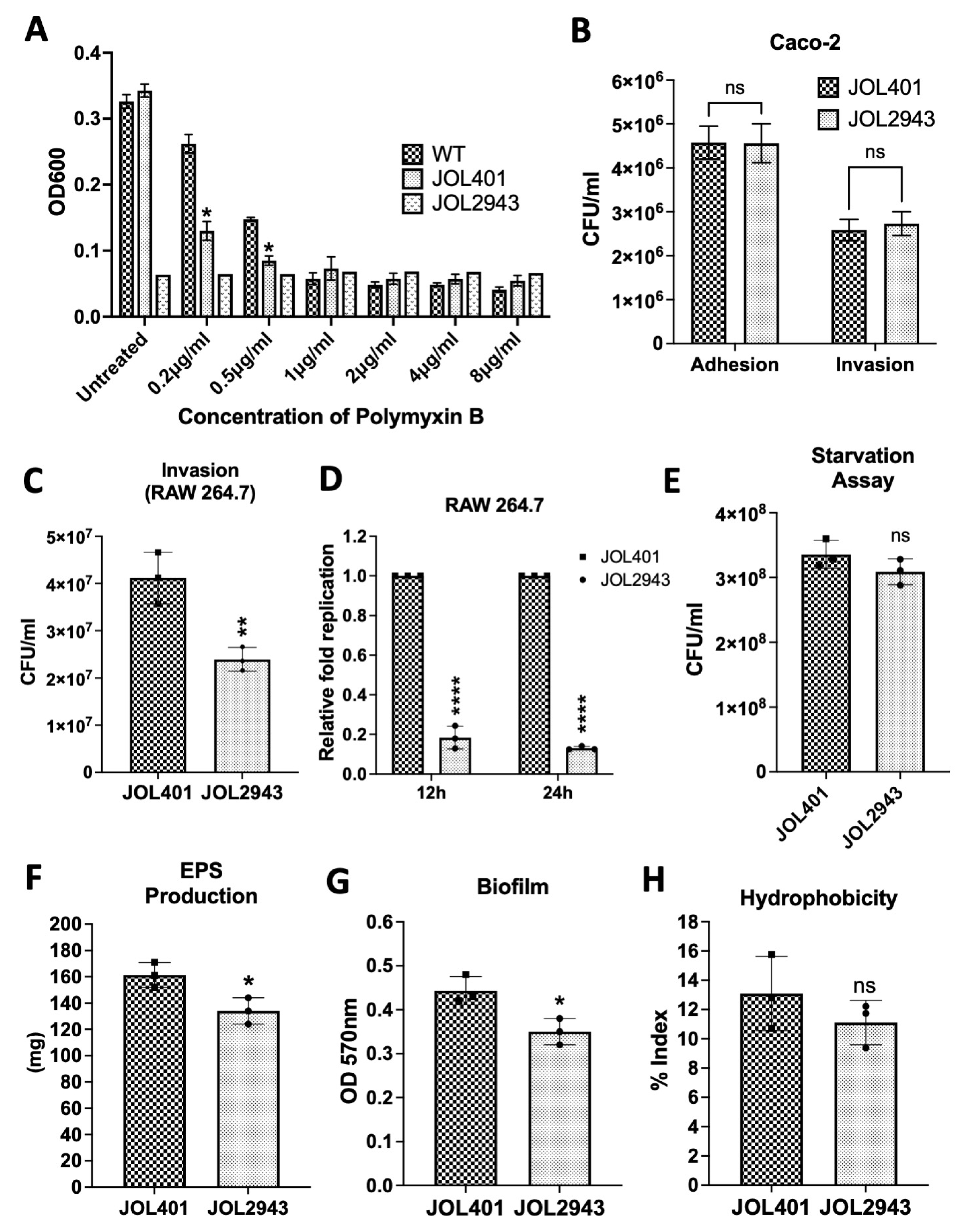
2.1. Effects of arnT Deletion on Polymyxin B Sensitivity, Adhesion, Invasion, Intracellular Survival, and Starvation
2.2. Effects of arnT Deletion on Biofilm, EPS Production, and Swarming
2.3. Assessment of Endotoxicity and Host Cytokine Responses
2.4. Serum Susceptibility Assay
2.5. In Vivo Host Colonization
3. Discussion
4. Materials and Methods
4.1. Animal Ethics Statement
4.2. Bacterial Strains and Growth Conditions, Plasmids, Primers
4.3. Gene Deletion
4.4. Polymyxin B Sensitivity
4.5. Adhesion, Invasion, and Intracellular Survival Assay
4.6. Starvation Assay
4.7. EPS Production Assay
4.8. Biofilm Assay
4.9. SEM and TEM Microscopic Visualization
4.10. MATH Assay
4.11. Swarming Assay
4.12. RT-qPCR Analysis of Cytokine Expression In Vitro and In Vivo
4.13. Serum Susceptibility Assay
4.14. In Vivo Host Colonization
4.15. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Jajere, S.M. A review of Salmonella enterica with particular focus on the pathogenicity and virulence factors, host specificity and antimicrobial resistance including multidrug resistance. Vet. World 2019, 12, 504–521. [Google Scholar] [CrossRef]
- Rabsch, W.; Andrews, H.L.; Kingsley, R.A.; Prager, R.; Tschäpe, H.; Adams, L.G.; Bäumler, A.J. Salmonella enterica Serotype Typhimurium and Its Host-Adapted Variants. Infect. Immun. 2002, 70, 2249–2255. [Google Scholar] [CrossRef]
- Becerra-Báez, E.I.; Meza-Toledo, S.E.; Muñoz-López, P.; Flores-Martínez, L.F.; Fraga-Pérez, K.; Magaño-Bocanegra, K.J.; Juárez-Hernández, U.; Mateos-Chávez, A.A.; Luria-Pérez, R. Recombinant Attenuated Salmonella enterica as a Delivery System of Heterologous Molecules in Cancer Therapy. Cancers 2022, 14, 4224. [Google Scholar] [CrossRef] [PubMed]
- Hegazy, W.A.H.; Hensel, M. Salmonella enterica as a Vaccine Carrier. Future Microbiol. 2012, 7, 111–127. [Google Scholar] [CrossRef]
- Moreno, M.; Kramer, M.G.; Yim, L.; Chabalgoity, J.A. Salmonella as Live Trojan Horse for Vaccine Development and Cancer Gene Therapy. Curr. Gene Ther. 2010, 10, 56–76. [Google Scholar] [CrossRef]
- Chorobik, P.; Marcinkiewicz, J. Therapeutic vaccines based on genetically modified Salmonella: A novel strategy in cancer immunotherapy. Pol. Arch. Med. Wewn. 2011, 121, 461–467. [Google Scholar] [CrossRef]
- Broadway, K.M.; Scharf, B.E. Salmonella Typhimurium as an Anticancer Therapy: Recent Advances and Perspectives. Curr. Clin. Microbiol. Rep. 2019, 6, 225–239. [Google Scholar] [CrossRef]
- Kawasaki, K. Complexity of lipopolysaccharide modifications in Salmonella enterica: Its effects on endotoxin activity, membrane permeability, and resistance to antimicrobial peptides. Food Res. Int. 2012, 45, 493–501. [Google Scholar] [CrossRef]
- Kumar, V. Toll-like receptors in sepsis-associated cytokine storm and their endogenous negative regulators as future immunomodulatory targets. Int. Immunopharmacol. 2020, 89, 107087. [Google Scholar] [CrossRef] [PubMed]
- Maeshima, N.; Fernandez, R.C. Recognition of lipid A variants by the TLR4-MD-2 receptor complex. Front. Cell. Infect. Microbiol. 2013, 3, 3. [Google Scholar] [CrossRef]
- Kong, Q.; Six, D.A.; Liu, Q.; Gu, L.; Wang, S.; Alamuri, P.; Raetz, C.R.H.; Curtiss, R. Phosphate Groups of Lipid A Are Essential for Salmonella enterica Serovar Typhimurium Virulence and Affect Innate and Adaptive Immunity. Infect. Immun. 2012, 80, 3215–3224. [Google Scholar] [CrossRef]
- Richards, S.M.; Strandberg, K.L.; Gunn, J.S. Salmonella-Regulated Lipopolysaccharide Modifications. Subcell. Biochem. 2010, 53, 101–122. [Google Scholar] [CrossRef]
- Needham, B.D.; Trent, M.S. Fortifying the barrier: The impact of lipid A remodelling on bacterial pathogenesis. Nat. Rev. Microbiol. 2013, 11, 467–481. [Google Scholar] [CrossRef]
- Zhou, Z.; Ribeiro, A.A.; Lin, S.; Cotter, R.J.; Miller, S.I.; Raetz, C.R. Lipid A Modifications in Polymyxin-resistant Salmonella Typhimurium: PmrA-Dependent 4-Amino-4-Deoxy-L-Arabinose, and Phosphoethanolamine Incorporation. J. Biol. Chem. 2001, 276, 43111–43121. [Google Scholar] [CrossRef]
- Matsuura, M. Structural Modifications of Bacterial Lipopolysaccharide that Facilitate Gram-Negative Bacteria Evasion of Host Innate Immunity. Front. Immunol. 2013, 4, 109. [Google Scholar] [CrossRef] [PubMed]
- Gunn, J.S.; Lim, K.B.; Krueger, J.; Kim, K.; Guo, L.; Hackett, M.; Miller, S.I. PmrA–PmrB-regulated genes necessary for 4-aminoarabinose lipid A modification and polymyxin resistance. Mol. Microbiol. 1998, 27, 1171–1182. [Google Scholar] [CrossRef] [PubMed]
- Kawasaki, K.; Ernst, R.K.; Miller, S.I. Inhibition of Salmonella enterica Serovar Typhimurium Lipopolysaccharide Deacylation by Aminoarabinose Membrane Modification. J. Bacteriol. 2005, 187, 2448–2457. [Google Scholar] [CrossRef] [PubMed]
- Moffat, J.H.; Harper, M.; Boyce, J.D. Mechanisms of polymyxin resistance. In Polymyxin Antibiotics: From Laboratory Bench to Bedside; Springer: Berlin/Heidelberg, Germany, 2019; Volume 1145, pp. 55–71. [Google Scholar]
- Lou, L.; Zhang, P.; Piao, R.; Wang, Y. Salmonella Pathogenicity Island 1 (SPI-1) and Its Complex Regulatory Network. Front. Cell. Infect. Microbiol. 2019, 9, 270. [Google Scholar] [CrossRef] [PubMed]
- Fass, E.; Groisman, E.A. Control of Salmonella pathogenicity island-2 gene expression. Curr. Opin. Microbiol. 2009, 12, 199–204. [Google Scholar] [CrossRef]
- Hapfelmeier, S.; Stecher, B.; Barthel, M.; Kremer, M.; Müller, A.J.; Heikenwalder, M.; Stallmach, T.; Hensel, M.; Pfeffer, K.; Akira, S.; et al. The Salmonella Pathogenicity Island (SPI)-2 and SPI-1 Type III Secretion Systems Allow Salmonella Serovar typhimurium to Trigger Colitis via MyD88-Dependent and MyD88-Independent Mechanisms. J. Immunol. 2005, 174, 1675–1685. [Google Scholar] [CrossRef] [PubMed]
- Norris, M.H.; Somprasong, N.; Schweizer, H.P.; Tuanyok, A. Lipid A Remodeling Is a Pathoadaptive Mechanism That Impacts Lipopolysaccharide Recognition and Intracellular Survival of Burkholderia pseudomallei. Infect. Immun. 2018, 86, e00360-18. [Google Scholar] [CrossRef]
- Yun, J.; Wang, X.; Zhang, L.; Li, Y. Effects of lipid A acyltransferases on the pathogenesis of F. novicida. Microb. Pathog. 2017, 109, 313–318. [Google Scholar] [CrossRef]
- Vogeleer, P.; Vincent, A.T.; Chekabab, S.M.; Charette, S.J.; Novikov, A.; Caroff, M.; Beaudry, F.; Jacques, M.; Harel, J. Escherichia coli O157:H7 Responds to Phosphate Starvation by Modifying LPS Involved in Biofilm Formation. bioRxiv 2019. bioRxiv:536201. [Google Scholar] [CrossRef]
- Ciornei, C.D.; Novikov, A.; Beloin, C.; Fitting, C.; Caroff, M.; Ghigo, J.-M.; Cavaillon, J.-M.; Adib-Conquy, M. Biofilm-forming Pseudomonas aeruginosa bacteria undergo lipopolysaccharide structural modifications and induce enhanced inflammatory cytokine response in human monocytes. Innate Immun. 2010, 16, 288–301. [Google Scholar] [CrossRef]
- Mireles, J.R.; Toguchi, A.; Harshey, R.M. Salmonella enterica Serovar Typhimurium Swarming Mutants with Altered Biofilm-Forming Abilities: Surfactin Inhibits Biofilm Formation. J. Bacteriol. 2001, 183, 5848–5854. [Google Scholar] [CrossRef]
- Hathroubi, S.; Beaudry, F.; Provost, C.; Martelet, L.; Segura, M.; Gagnon, C.A.; Jacques, M. Impact of Actinobacillus pleuropneumoniae biofilm mode of growth on the lipid A structures and stimulation of immune cells. Innate Immun. 2016, 22, 353–362. [Google Scholar] [CrossRef]
- Hewawaduge, C.; Senevirathne, A.; Sivasankar, C.; Lee, J.H. The impact of lipid A modification on biofilm and related pathophysiological phenotypes, endotoxicity, immunogenicity, and protection of Salmonella Typhimurium. Vet. Microbiol. 2023, 282, 109759. [Google Scholar] [CrossRef] [PubMed]
- Rana, K.; Nayak, S.R.; Bihary, A.; Sahoo, A.K.; Mohanty, K.C.; Palo, S.K.; Sahoo, D.; Pati, S.; Dash, P. Association of quorum sensing and biofilm formation with Salmonella virulence: Story beyond gathering and cross-talk. Arch. Microbiol. 2021, 203, 5887–5897. [Google Scholar] [CrossRef] [PubMed]
- Lee, C.; Mannaa, M.; Kim, N.; Kim, J.; Choi, Y.; Kim, S.H.; Jung, B.; Lee, H.-H.; Lee, J.; Seo, Y.-S. Stress Tolerance and Virulence-Related Roles of Lipopolysaccharide in Burkholderia glumae. Plant Pathol. J. 2019, 35, 445–458. [Google Scholar] [CrossRef] [PubMed]
- Bowden, S.D.; Hale, N.; Chung, J.C.S.; Hodgkinson, J.T.; Spring, D.R.; Welch, M. Surface swarming motility by Pectobacterium atrosepticum is a latent phenotype that requires O antigen and is regulated by quorum sensing. Microbiology 2013, 159, 2375–2385. [Google Scholar] [CrossRef] [PubMed]
- Mastroeni, P.; Bryant, C. Cytokines in Salmonellosis. EcoSal Plus 2004, 1, 10-1128. [Google Scholar] [CrossRef]
- Nawab, A.; An, L.; Wu, J.; Li, G.; Liu, W.; Zhao, Y.; Wu, Q.; Xiao, M. Chicken toll-like receptors and their significance in immune response and disease resistance. Int. Rev. Immunol. 2019, 38, 284–306. [Google Scholar] [CrossRef]
- Rolin, O.; Muse, S.J.; Safi, C.; Elahi, S.; Gerdts, V.; Hittle, L.E.; Ernst, R.K.; Harvill, E.T.; Preston, A. Enzymatic Modification of Lipid A by ArnT Protects Bordetella bronchiseptica against Cationic Peptides and Is Required for Transmission. Infect. Immun. 2014, 82, 491–499. [Google Scholar] [CrossRef]
- Sivasankar, C.; Hewawaduge, C.; Lee, J.H. Screening of lipid-A related genes and development of low-endotoxicity live-attenuated Salmonella gallinarum by arnT deletion that elicits immune responses and protection against fowl typhoid in chickens. Dev. Comp. Immunol. 2023, 145, 104707. [Google Scholar] [CrossRef]
- Giulietti, A.; Overbergh, L.; Valckx, D.; Decallonne, B.; Bouillon, R.; Mathieu, C. An Overview of Real-Time Quantitative PCR: Applications to Quantify Cytokine Gene Expression. Methods 2001, 25, 386–401. [Google Scholar] [CrossRef]
- Lee, S.; Kwak, J.-H.; Kim, S.H.; Bin Jeong, T.; Son, S.W.; Kim, J.-H.; Lim, Y.; Cho, J.-Y.; Hwang, D.Y.; Kim, K.S.; et al. Comparative study of liver injury induced by high-fat methionine- and choline-deficient diet in ICR mice originating from three different sources. Lab. Anim. Res. 2019, 35, 15. [Google Scholar] [CrossRef] [PubMed]
- Doublet, B.; Douard, G.; Targant, H.; Meunier, D.; Madec, J.-Y.; Cloeckaert, A. Antibiotic marker modifications of λ Red and FLP helper plasmids, pKD46 and pCP20, for inactivation of chromosomal genes using PCR products in multidrug-resistant strains. J. Microbiol. Methods 2008, 75, 359–361. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Berglund, B.; Zhu, Y.; Luo, Q.; Xiao, Y. Performance of different methods for testing polymyxin B: Comparison of broth microdilution, agar dilution and MIC test strip in mcr-1 positive and negative Escherichia coli. Lett. Appl. Microbiol. 2021, 73, 197–205. [Google Scholar] [CrossRef] [PubMed]
- Behera, B.; Mathur, P.; Das, A.; Kapil, A.; Gupta, B.; Bhoi, S.; Farooque, K.; Sharma, V.; Misra, M.; Behera, B.; et al. Evaluation of susceptibility testing methods for polymyxin. Int. J. Infect. Dis. 2010, 14, e596–e601. [Google Scholar] [CrossRef] [PubMed]
- Arunima, A.; Suar, M. Glucose Starvation, Magnesium Ion Starvation, and Bile Stress Assays. Bio-Protocol 2021, 11, e4157. [Google Scholar] [CrossRef] [PubMed]
- Neiger, M.R.; González, J.F.; Gonzalez-Escobedo, G.; Kuck, H.; White, P.; Gunn, J.S. Pathoadaptive Alteration of Salmonella Biofilm Formation in Response to the Gallbladder Environment. J. Bacteriol. 2019, 201, 10–1128. [Google Scholar] [CrossRef]
- Hamadi, F.; Latrache, H.; Zahir, H.; Bengourram, J.; Kouider, N.; Elghmari, A.; Habbari, K. Evaluation of the relative cell surface charge by using microbial adhesion to hydrocarbon. Microbiology 2011, 80, 488–491. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D.; Livak, K.J.; Schmittgen, T.D. Analysis of Relative Gene Expression Data Using Real-Time Quantitative PCR and the 2−ΔΔCT Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Zou, Y.; Feng, S.; Xu, C.; Zhang, B.; Zhou, S.; Zhang, L.; He, X.; Li, J.; Yang, Z.; Liao, M. The role of galU and galE of Haemophilus parasuis SC096 in serum resistance and biofilm formation. Vet. Microbiol. 2012, 162, 278–284. [Google Scholar] [CrossRef] [PubMed]
- Guo, L.; Dai, H.; Feng, S.; Zhao, Y. Contribution of GalU to biofilm formation, motility, antibiotic and serum resistance, and pathogenicity of Salmonella Typhimurium. Front. Cell. Infect. Microbiol. 2023, 13, 1149541. [Google Scholar] [CrossRef]
- Abou-Shleib, H.; Elkhouly, A.; Roantree, R.J. Effect of selected plasmids on the serum sensitivity of Salmonella Typhimurium strains with defined lipopolysaccharide core defects. FEMS Microbiol. Lett. 1986, 36, 27–30. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Bacterial Strains/ Plasmids/Primers | Reference | |
|---|---|---|
| Bacterial Strains | ||
| JOL401 | Salmonella Typhimurium wild-type | Lab stock |
| JOL2943 | Salmonella Typhimurium ∆arnT mutant | This study |
| Plasmids | ||
| pKD46 | Ori101-repA101ts; encodes Lambda Red genes (exo, bet, gam); native terminator (tL3); arabinose-inducible for expression (ParaB); bla | [35] |
| pKD3 | oriR6K gamma, bla (ampR), rgnB (Ter), catR, FRT | [35] |
| pCP20 | Helper plasmid containing a temperature-inducible flp gene for removing the FRT-flanked chloramphenicol gene | [35] |
| Gene Deletion primers | ||
| arnT pKD3 | Forward— GAGCTGACCGCCAACGCTGAGCAGACTGGCAAGCACCAGAATGACGCCGAGTGTAGGCTGGAGCTGCTTC Reverse— ATCCCTGGCCGTGAAGGTTGGCTGGGGTGCCAACAGGCAGCGAGCGCCTCATGGGAATTAGCCATGGTCC | [35] |
| arnT inner | Forward—GCAACGCGGTACGTTTATCC Reverse—GAAACGCGCTATGCCGAAAT | [35] |
| arnT flanking | Forward—GAGCTGACCGCCAACGCTGA Reverse—ATCCCTGGCCGTGAAGGTTG | [35] |
| qPCR cytokine primers | ||
| INF-γ | Forward—TCAAGTGGCATAGATGTGGAAGAA Reverse—TGGCTCTGCAGGATTTTCATG | [36] |
| TNF-α | Forward—CATCTTCTCAAAATTCGAGTGACAA Reverse—TGGGAGTAGACAAGGTACAACCC | [36] |
| IL-1β | Forward—TTCACCATGGAATCCGTGTC Reverse—GTCTTGGCCGAGGACTAAGG | [37] |
| GAPDH | Forward—TCACCACCATGGAGAAGGC Reverse—GCTAAGCAGTTGGTGGTGCA | [36] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sivasankar, C.; Lloren, K.K.S.; Lee, J.H. Deciphering the Interrelationship of arnT Involved in Lipid-A Alteration with the Virulence of Salmonella Typhimurium. Int. J. Mol. Sci. 2024, 25, 2760. https://doi.org/10.3390/ijms25052760
Sivasankar C, Lloren KKS, Lee JH. Deciphering the Interrelationship of arnT Involved in Lipid-A Alteration with the Virulence of Salmonella Typhimurium. International Journal of Molecular Sciences. 2024; 25(5):2760. https://doi.org/10.3390/ijms25052760
Chicago/Turabian StyleSivasankar, Chandran, Khristine Kaith Sison Lloren, and John Hwa Lee. 2024. "Deciphering the Interrelationship of arnT Involved in Lipid-A Alteration with the Virulence of Salmonella Typhimurium" International Journal of Molecular Sciences 25, no. 5: 2760. https://doi.org/10.3390/ijms25052760