
Phosphate (Pi) Transporter PIT1 Induces Pi Starvation in Salmonella-Containing Vacuole in HeLa Cells
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. S. Typhimurium Infection Induces PIT1 Expression in HeLa Cells
2.2. PIT1 Affects S. Typhimurium Intracellular Replication
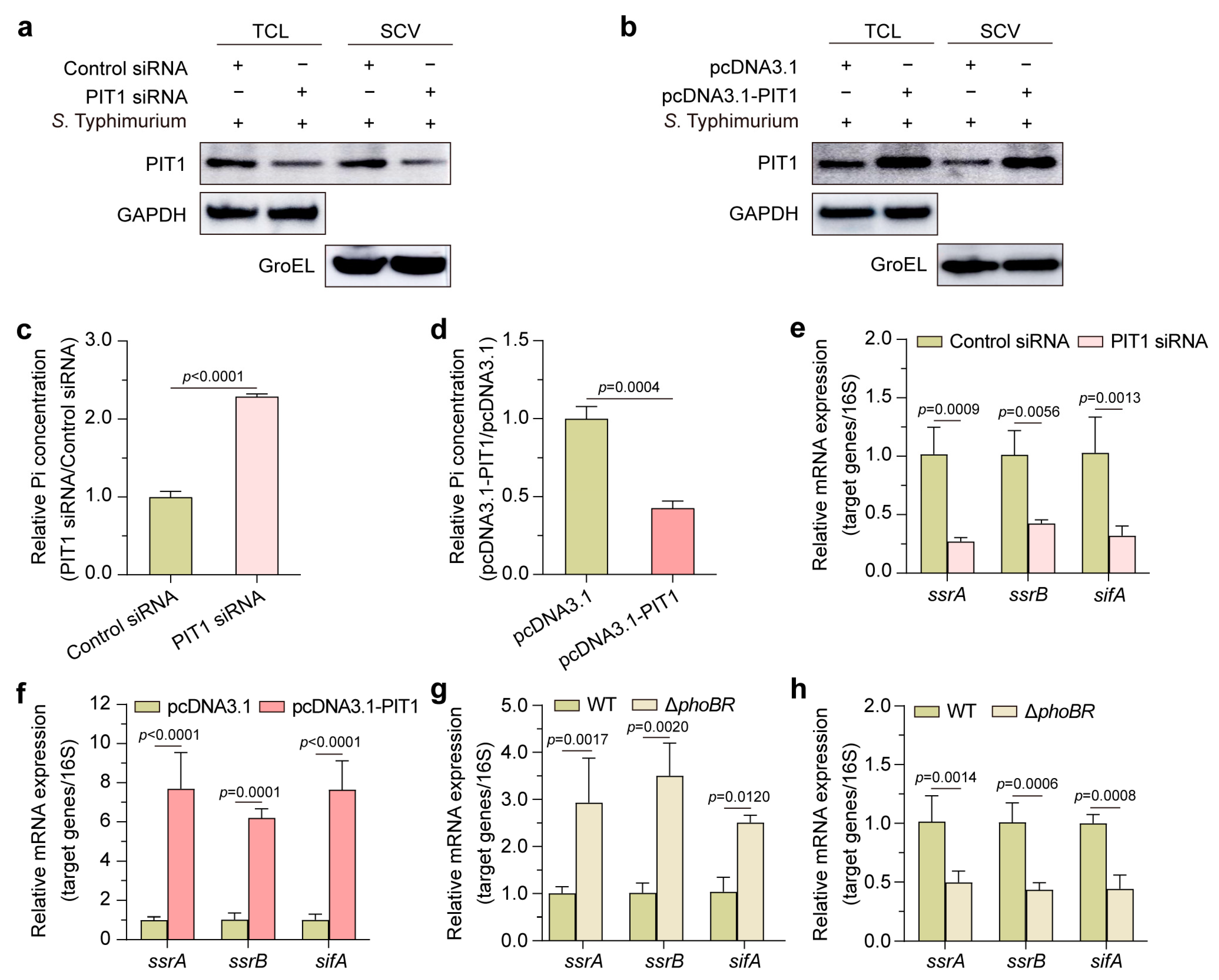
2.3. PIT1 Transports Pi into Cytosol and Regulates SPI-2 Genes via PhoBR
2.4. Upregulated PIT1 Prevents the Fusion of SCVs with Lysosomes
2.5. LPS of S. Typhimurium Triggers PIT1 in HeLa Cells
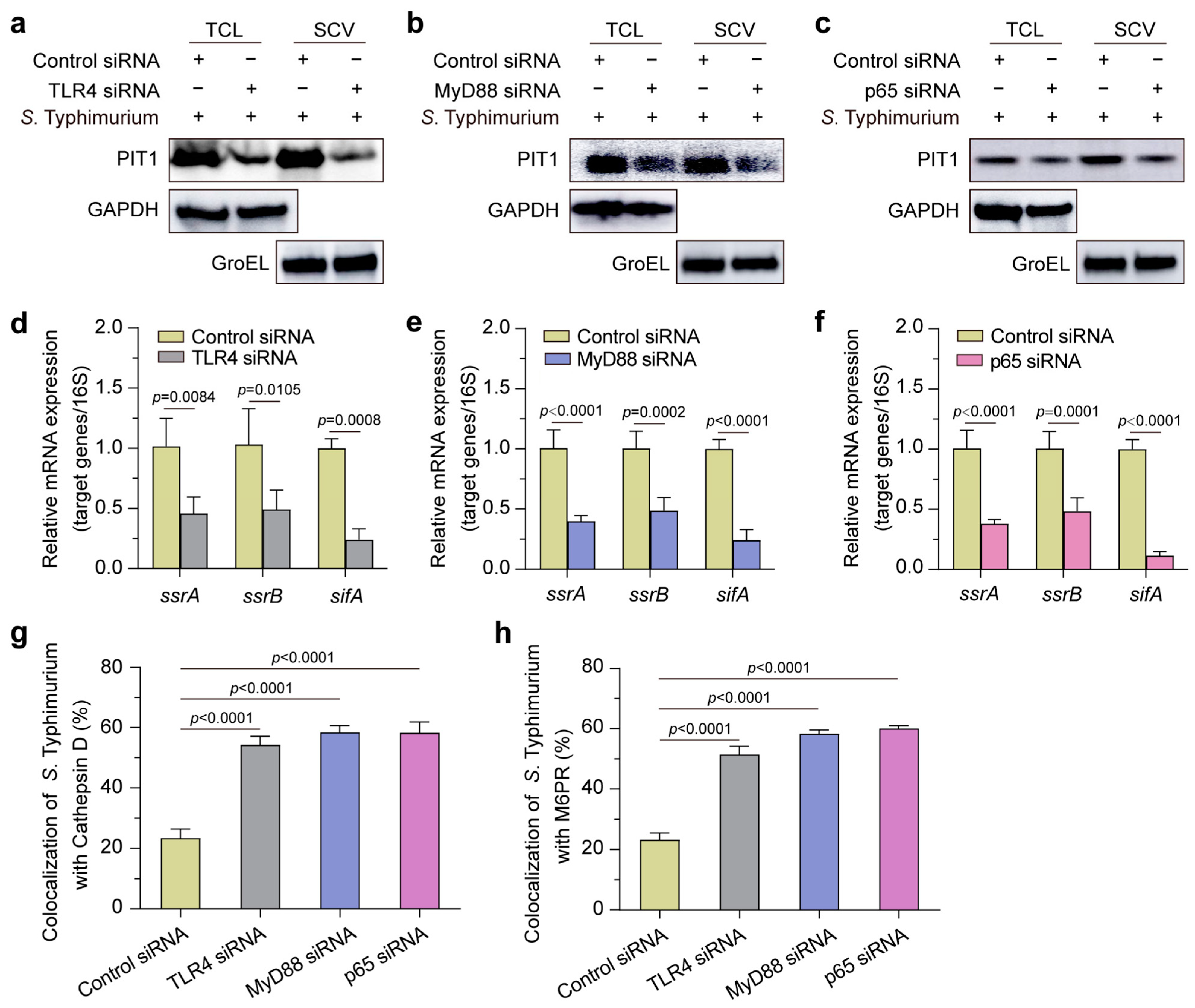
2.6. S. Typhimurium Exploits TLR4-MyD88-NF-kB Pathway to Regulate PIT1
3. Discussion
4. Materials and Methods
4.1. Bacterial Strains
4.2. Antibodies
4.3. Cell Culture and Transfection
4.4. RNA Extraction and qRT-PCR
4.5. Quantitative PCR Analysis
4.6. Bacterial Infection and Replication Assays
4.7. Immunofluorescence Analysis
4.8. Isolation of Intracellular SCVs
4.9. Western Blotting Analysis
4.10. Determination of the Phosphate Concentration in SCVs
4.11. LPS Stimulation
4.12. Quantification and Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Haraga, A.; Ohlson, M.B.; Miller, S.I. Salmonellae interplay with host cells. Nat. Rev. Microbiol. 2008, 6, 53–66. [Google Scholar] [CrossRef] [PubMed]
- Besser, J.M. Salmonella epidemiology: A whirlwind of change. Food Microbiol. 2018, 71, 55–59. [Google Scholar] [CrossRef] [PubMed]
- Kotloff, K.L. The burden and etiology of diarrheal illness in developing countries. Pediatr. Clin. N. Am. 2017, 64, 799–814. [Google Scholar] [CrossRef] [PubMed]
- Majowicz, S.E.; Musto, J.; Scallan, E.; Angulo, F.J.; Kirk, M.; O’Brien, S.J.; Jones, T.F.; Fazil, A.; Hoekstra, R.M. The global burden of nontyphoidal Salmonella gastroenteritis. Clin. Infect. Dis. 2010, 50, 882–889. [Google Scholar] [CrossRef]
- Fabrega, A.; Vila, J. Salmonella enterica serovar Typhimurium skills to succeed in the host: Virulence and regulation. Clin. Microbiol. Rev. 2013, 26, 308–341. [Google Scholar] [CrossRef]
- Pui, C.F.; Wong, W.C.; Chai, L.C.; Tunung, R.; Jeyaletchumi, P.; Noor Hidayah, M.S.; Ubong, A.; Farinazleen, M.G.; Cheah, Y.K.; Son, R. Salmonella: A foodborne pathogen. Int. Food Res. J. 2011, 18, 465–473. [Google Scholar]
- Patra, S.D.; Mohakud, N.K.; Panda, R.K.; Sahu, B.R.; Suar, M. Prevalence and multidrug resistance in Salmonella enterica Typhimurium: An overview in South East Asia. World J. Microbiol. Biotechnol. 2021, 37, 185. [Google Scholar] [CrossRef] [PubMed]
- Monack, D.M.; Bouley, D.M.; Falkow, S. Salmonella typhimurium persists within macrophages in the mesenteric lymph nodes of chronically infected Nramp1+/+ mice and can be reactivated by IFNγ neutralization. J. Exp. Med. 2004, 199, 231–241. [Google Scholar] [CrossRef]
- Prost, L.R.; Sanowar, S.; Miller, S.I. Salmonella sensing of anti-microbial mechanisms to promote survival within macrophages. Immunol. Rev. 2007, 219, 55–65. [Google Scholar] [CrossRef]
- Leung, K.Y.; Finlay, B.B. Intracellular replication is essential for the virulence of Salmonella typhimurium. Proc. Natl. Acad. Sci. USA 1991, 88, 11470–11474. [Google Scholar] [CrossRef]
- Hautefort, I.; Thompson, A.; Eriksson-Ygberg, S.; Parker, M.L.; Lucchini, S.; Danino, V.; Bongaerts, R.J.; Ahmad, N.; Rhen, M.; Hinton, J.C. During infection of epithelial cells Salmonella enterica serovar Typhimurium undergoes a time-dependent transcriptional adaptation that results in simultaneous expression of three type 3 secretion systems. Cell. Microbiol. 2008, 10, 958–984. [Google Scholar] [CrossRef] [PubMed]
- Steele-Mortimer, O. The Salmonella-containing vacuole: Moving with the times. Curr. Opin. Microbiol. 2008, 11, 38–45. [Google Scholar] [CrossRef] [PubMed]
- Brumell, J.H.; Tang, P.; Mills, S.D.; Finlay, B.B. Characterization of Salmonella-induced filaments (Sifs) reveals a delayed interaction between Salmonella-containing vacuoles and late endocytic compartments. Traffic 2001, 2, 643–653. [Google Scholar] [CrossRef] [PubMed]
- Liss, V.; Swart, A.L.; Kehl, A.; Hermanns, N.; Zhang, Y.; Chikkaballi, D.; Bohles, N.; Deiwick, J.; Hensel, M. Salmonella enterica remodels the host cell endosomal system for efficient intravacuolar nutrition. Cell Host Microbe 2017, 21, 390–402. [Google Scholar] [CrossRef] [PubMed]
- Portillo, F.G.; Finlay, B.B. Targeting of Salmonella typhimurium to vesicles containing lysosomal membrane glycoproteins bypasses compartments with mannose 6-phosphate receptors. J. Cell. Biol. 1995, 129, 81–97. [Google Scholar] [CrossRef] [PubMed]
- Garvis, S.G.; Beuzon, C.R.; Holden, D.W. A role for the PhoP/Q regulon in inhibition of fusion between lysosomes and Salmonella-containing vacuoles in macrophages. Cell. Microbiol. 2001, 3, 731–744. [Google Scholar] [CrossRef]
- Marzan, L.W.; Shimizu, K. Metabolic regulation of Escherichia coli and its phoB and phoR genes knockout mutants under phosphate and nitrogen limitations as well as at acidic condition. Microb. Cell Fact. 2011, 10, 39. [Google Scholar] [CrossRef]
- Schuhmacher, T.; Löffler, M.; Hurler, T.; Takors, R. Phosphate limited fed-batch processes: Impact on carbon usage and energy metabolism in Escherichia coli. J. Biotechnol. 2014, 190, 96–104. [Google Scholar] [CrossRef]
- Jiang, J.; Yu, K.; Qi, L.; Liu, Y.; Cheng, S.; Wu, M.; Wang, Z.; Fu, J.; Liu, X. A proteomic view of Salmonella Typhimurium in response to phosphate limitation. Proteomes 2018, 6, 19. [Google Scholar] [CrossRef]
- Garmendia, J.; Beuzón, C.R.; Ruiz-Albert, J.; Holden, D.W. The roles of SsrA–SsrB and OmpR–EnvZ in the regulation of genes encoding the Salmonella typhimurium SPI-2 type III secretion system. Microbiology 2003, 149, 2385–2396. [Google Scholar] [CrossRef]
- McGhie, E.J.; Brawn, L.C.; Hume, P.J.; Humphreys, D.; Koronakis, V. Salmonella takes control: Effector-driven manipulation of the host. Curr. Opin. Microbiol. 2009, 12, 117–124. [Google Scholar] [CrossRef] [PubMed]
- Tuli, A.; Sharma, M. How to do business with lysosomes: Salmonella leads the way. Curr. Opin. Microbiol. 2019, 47, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Beuzón, C.R.; Méresse, S.; Unsworth, K.E.; Ruíz-Albert, J.; Garvis, S.; Waterman, S.R.; Ryder, T.A.; Boucrot, E.; Holden, D.W. Salmonella maintains the integrity of its intracellular vacuole through the action of SifA. EMBO J. 2000, 19, 3235–3249. [Google Scholar] [CrossRef] [PubMed]
- McGourty, K.; Thurston, T.L.; Matthews, S.A.; Pinaud, L.; Mota, L.J.; Holden, D.W. Salmonella inhibits retrograde trafficking of mannose-6-phosphate receptors and lysosome function. Science 2012, 338, 963–967. [Google Scholar] [CrossRef]
- Lensmire, J.M.; Pratt, Z.L.; Wong, A.C.L.; Kaspar, C.W. Phosphate and carbohydrate facilitate the formation of filamentous Salmonella enterica during osmotic stress. Microbiology 2018, 164, 1503–1513. [Google Scholar] [CrossRef]
- Nishimura, M.; Naito, S. Tissue-specific mRNA expression profiles of human solute carrier transporter superfamilies. Drug Metab. Pharmacokinet. 2008, 23, 22–44. [Google Scholar] [CrossRef] [PubMed]
- Pang, Y.; Cheng, Z.; Zhang, S.; Li, S.; Li, X.; Li, X.; Zhang, X.; Li, X.; Feng, Y.; Cui, H.; et al. Bladder epithelial cell phosphate transporter inhibition protects mice against uropathogenic Escherichia coli infection. Cell Rep. 2022, 39, 110698. [Google Scholar] [CrossRef] [PubMed]
- Kroger, C.; Colgan, A.; Srikumar, S.; Handler, K.; Sivasankaran, S.K.; Hammarlof, D.L.; Canals, R.; Grissom, J.E.; Conway, T.; Hokamp, K.; et al. An infection-relevant transcriptomic compendium for Salmonella enterica Serovar Typhimurium. Cell Host Microbe 2013, 14, 683–695. [Google Scholar] [CrossRef]
- Jiang, L.; Wang, P.; Li, X.; Lv, R.; Wang, L.; Yang, B.; Huang, D.; Feng, L.; Liu, B. PagR mediates the precise regulation of Salmonella pathogenicity island 2 gene expression in response to magnesium and phosphate signals in Salmonella Typhimurium. Cell. Microbiol. 2020, 22, e13125. [Google Scholar] [CrossRef]
- Jiang, W.; Metcalf, W.; Li, K.-S.; Wanner, B.L. Molecular cloning, mapping, and regulation of Pho regulon genes for phosphonate breakdown by the phosphonatase pathway of Salmonella typhimurium LT2. J. Bacteriol. 1995, 177, 6411–6421. [Google Scholar] [CrossRef]
- Allgood, S.C.; Neunuebel, M.R. The recycling endosome and bacterial pathogens. Cell. Microbiol. 2018, 20, e12857. [Google Scholar] [CrossRef] [PubMed]
- Koumakis, E.; Millet-Botti, J.; Benna, J.E.; Leroy, C.; Boitez, V.; Codogno, P.; Friedlander, G.; Forand, A. Novel function of PiT1/SLC20A1 in LPS-related inflammation and wound healing. Sci. Rep. 2019, 9, 1808. [Google Scholar] [CrossRef] [PubMed]
- Kong, Q.; Six, D.A.; Liu, Q.; Gu, L.; Roland, K.L.; Raetz, C.R.; Curtiss, R., 3rd. Palmitoylation state impacts induction of innate and acquired immunity by the Salmonella enterica serovar typhimurium msbB mutant. Infect. Immun. 2011, 79, 5027–5038. [Google Scholar] [CrossRef] [PubMed]
- Kumar, H.; Kawai, T.; Akira, S. Toll-like receptors and innate immunity. Biochem. Biophys. Res. Commun. 2009, 388, 621–625. [Google Scholar] [CrossRef] [PubMed]
- Runyen-Janecky, L.J.; Payne, S.M. Identification of chromosomal Shigella flexneri genes induced by the eukaryotic intracellular environment. Infect. Immun. 2002, 70, 4379–4388. [Google Scholar] [CrossRef] [PubMed]
- Steele-Mortimer, O.; MeÂresse, S.; Gorvel, J.; Toh, B.; Finlay, B. Biogenesis of Salmonella typhimurium-containing vacuoles in epithelial cells involves interactions with the early endocytic pathway. Cell. Microbiol. 1999, 1, 33–49. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Yin, H.; Zhao, M.; Lu, Q. TLR2 and TLR4 in autoimmune diseases: A comprehensive review. Clin. Rev. Allergy Immunol. 2014, 47, 136–147. [Google Scholar] [CrossRef] [PubMed]
- Michigami, T. Extracellular phosphate as a signaling molecule. Contrib. Nephrol. 2013, 180, 14–24. [Google Scholar]
- Bruna, R.E.; Kendra, C.G.; Pontes, M.H. Coordination of phosphate and magnesium metabolism in bacteria. Adv. Exp. Med. Biol. 2022, 1362, 135–150. [Google Scholar]
- Kaushal, B.; Paul, S.; Hulett, F.M. Direct regulation of bacillus subtilis phoPR transcription by transition state regulator ScoC. J. Bacteriol. 2010, 192, 3103–3113. [Google Scholar] [CrossRef]
- Botella, E.; Devine, S.K.; Hubner, S.; Salzberg, L.I.; Gale, R.T.; Brown, E.D.; Link, H.; Sauer, U.; Codée, J.D.; Noone, D.; et al. PhoR autokinase activity is controlled by an intermediate in wall teichoic acid metabolism that is sensed by the intracellular PAS domain during the PhoPR-mediated phosphate limitation response of Bacillus subtilis. Mol. Microbiol. 2014, 94, 1242–1259. [Google Scholar] [CrossRef] [PubMed]
- Sun, G.; Birkey, S.M.; Hulett, F.M. Three two-component signal-transduction systems interact for Pho regulation in Bacillus subtilis. Mol. Microbiol. 1996, 19, 941–948. [Google Scholar] [CrossRef] [PubMed]
- Wanner, B.L. Gene regulation by phosphate in enteric bacteria. J. Cell. Biochem. 1993, 51, 47–54. [Google Scholar] [CrossRef] [PubMed]
- Salzberg, L.I.; Botella, E.; Hokamp, K.; Antelmann, H.; Maaß, S.; Becher, D.; Noone, D.; Devine, K.M.; Silhavy, T.J. Genome-wide analysis of phosphorylated PhoP binding to chromosomal DNA reveals several novel features of the PhoPR-mediated phosphate limitation response in Bacillus subtilis. J. Bacteriol. 2015, 197, 1492–1506. [Google Scholar] [CrossRef] [PubMed]
- Alix, E.; Mukherjee, S.; Roy, C.R. Subversion of membrane transport pathways by vacuolar pathogens. J. Cell. Biol. 2011, 195, 943–952. [Google Scholar] [CrossRef] [PubMed]
- Figueira, R.; Holden, D.W. Functions of the Salmonella pathogenicity island 2 (SPI-2) type III secretion system effectors. Microbiology 2012, 158, 1147–1161. [Google Scholar] [CrossRef] [PubMed]
- Brumell, J.H.; Grinstein, S. Salmonella redirects phagosomal maturation. Curr. Opin. Microbiol. 2004, 7, 78–84. [Google Scholar] [CrossRef]
- D’Costa, V.M.; Braun, V.; Landekic, M.; Shi, R.; Proteau, A.; McDonald, L.; Cygler, M.; Grinstein, S.; Brumell, J.H. Salmonella disrupts host endocytic trafficking by SopD2-mediated inhibition of Rab7. Cell Rep. 2015, 12, 1508–1518. [Google Scholar] [CrossRef]
- Ohlson, M.B.; Fluhr, K.; Birmingham, C.L.; Brumell, J.H.; Miller, S.I. SseJ deacylase activity by Salmonella enterica serovar Typhimurium promotes virulence in mice. Infect. Immun. 2005, 73, 6249–6259. [Google Scholar] [CrossRef]
- Jiang, L.; Wang, P.; Song, X.; Zhang, H.; Ma, S.; Wang, J.; Li, W.; Lv, R.; Liu, X.; Ma, S.; et al. Salmonella Typhimurium reprograms macrophage metabolism via T3SS effector SopE2 to promote intracellular replication and virulence. Nat. Commun. 2021, 12, 879. [Google Scholar] [CrossRef]
- Li, W.; Li, L.; Yan, X.; Wu, P.; Zhang, T.; Fan, Y.; Ma, S.; Wang, X.; Jiang, L. Nitrate utilization promotes systemic infection of Salmonella Typhimurium in mice. Int. J. Mol. Sci. 2022, 23, 7220. [Google Scholar] [CrossRef] [PubMed]
- Schmittgen, T.D.; Livak, K.J. Analyzing real-time PCR data by the comparative C(T) method. Nat. Protoc. 2008, 3, 1101–1108. [Google Scholar] [CrossRef] [PubMed]
- Cheng, Z.; Kumagai, Y.; Lin, M.; Zhang, C.; Rikihisa, Y. Intra-leukocyte expression of two-component systems in Ehrlichia chaffeensis and Anaplasma phagocytophilum and effects of the histidine kinase inhibitor closantel. Cell. Microbiol. 2006, 8, 1241–1252. [Google Scholar] [CrossRef] [PubMed]
- Ma, S.; Jiang, L.; Wang, J.; Liu, X.; Li, W.; Ma, S.; Feng, L. Downregulation of a novel flagellar synthesis regulator AsiR promotes intracellular replication and systemic pathogenicity of Salmonella typhimurium. Virulence 2021, 12, 298–311. [Google Scholar] [CrossRef]
- Chen, S.; Li, Y.; Chu, B.; Yuan, L.; Liu, N.; Zhu, Y.; Wang, J. Lactobacillus johnsonii L531 alleviates the damage caused by Salmonella Typhimurium via inhibiting TLR4, NF-κB, and NLRP3 inflammasome signaling pathways. Microorganisms 2021, 9, 1983. [Google Scholar] [CrossRef]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, W.; Feng, Y.; Yan, J.; Kang, C.; Yao, T.; Sun, H.; Cheng, Z. Phosphate (Pi) Transporter PIT1 Induces Pi Starvation in Salmonella-Containing Vacuole in HeLa Cells. Int. J. Mol. Sci. 2023, 24, 17216. https://doi.org/10.3390/ijms242417216
Yang W, Feng Y, Yan J, Kang C, Yao T, Sun H, Cheng Z. Phosphate (Pi) Transporter PIT1 Induces Pi Starvation in Salmonella-Containing Vacuole in HeLa Cells. International Journal of Molecular Sciences. 2023; 24(24):17216. https://doi.org/10.3390/ijms242417216
Chicago/Turabian StyleYang, Wen, Yingxing Feng, Jun Yan, Chenbo Kang, Ting Yao, Hongmin Sun, and Zhihui Cheng. 2023. "Phosphate (Pi) Transporter PIT1 Induces Pi Starvation in Salmonella-Containing Vacuole in HeLa Cells" International Journal of Molecular Sciences 24, no. 24: 17216. https://doi.org/10.3390/ijms242417216