

Intrinsic Lipid Curvature and Bilayer Elasticity as Regulators of Channel Function: A Comparative Single-Molecule Study


Abstract
:1. Introduction

2. Results
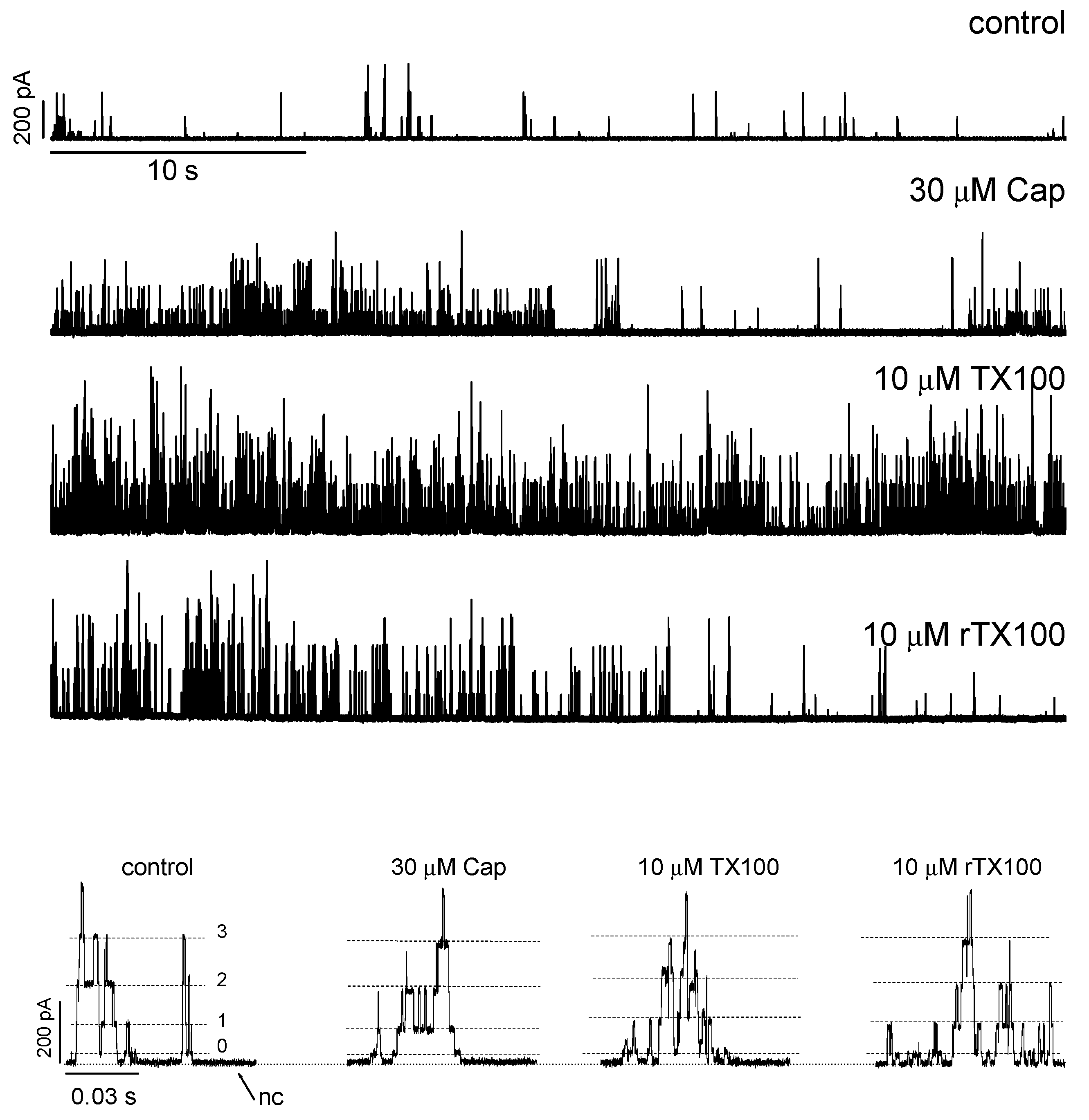
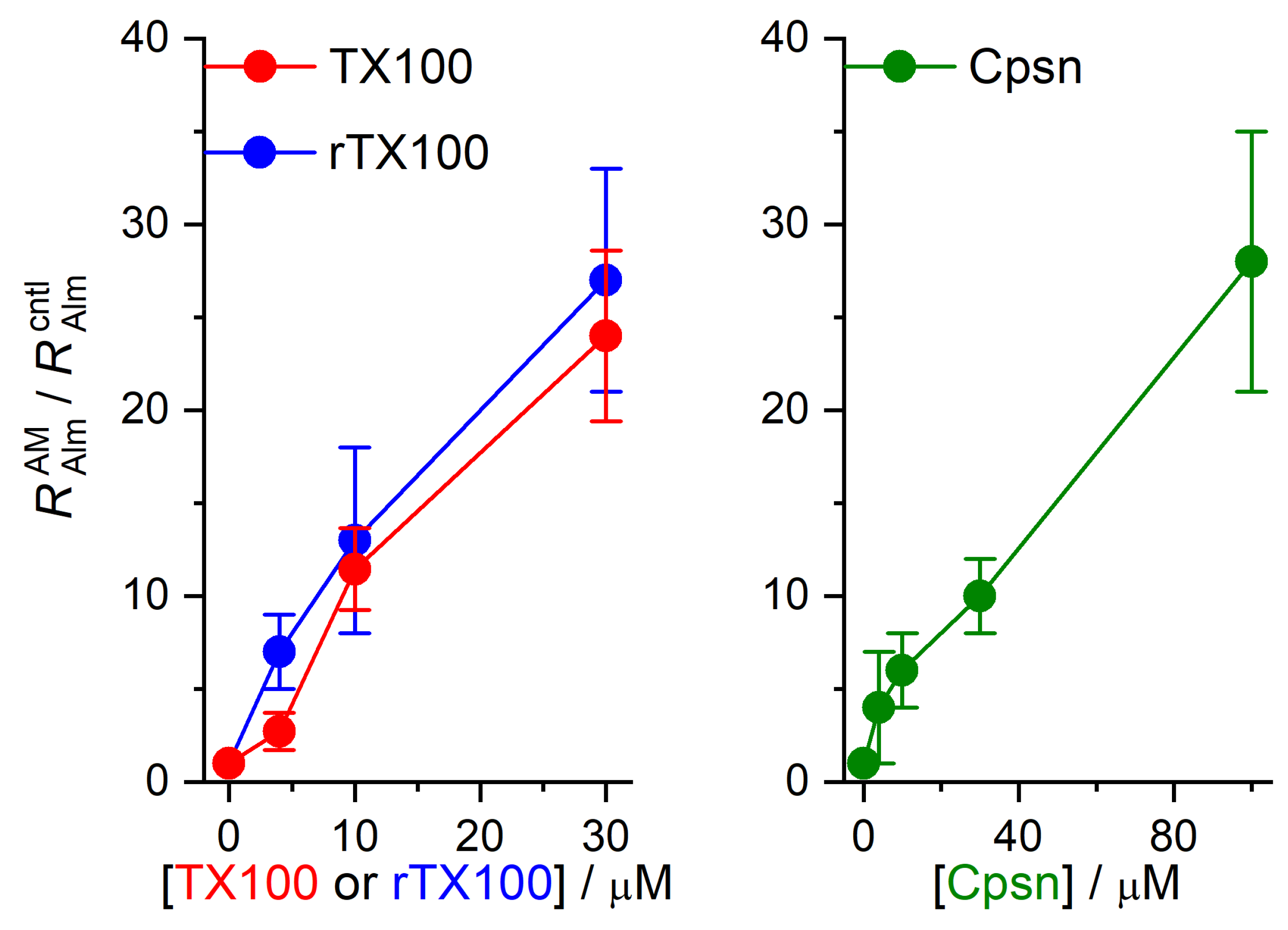
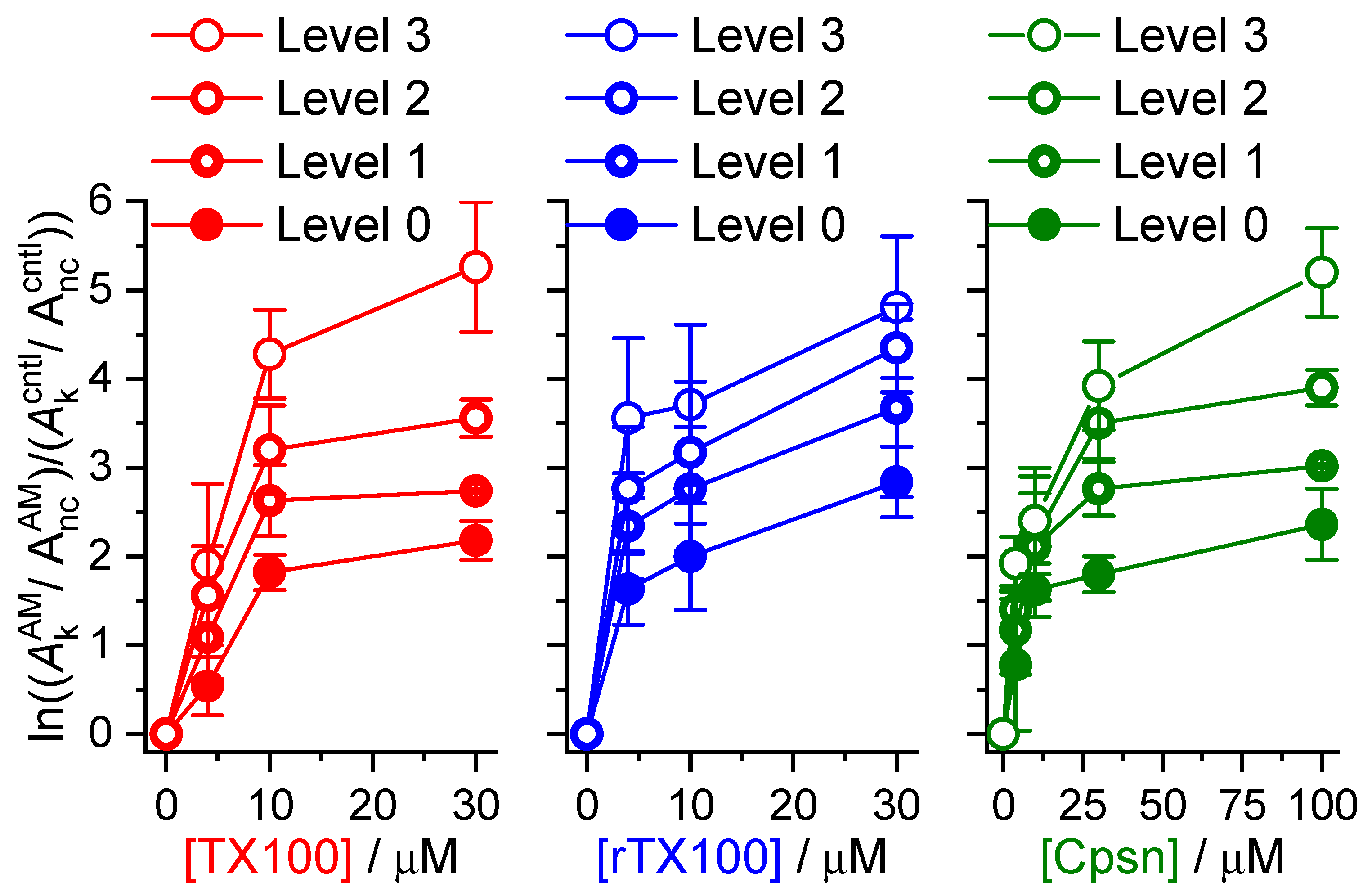
2.1. Amphiphiles Modulate Alm Channel Function
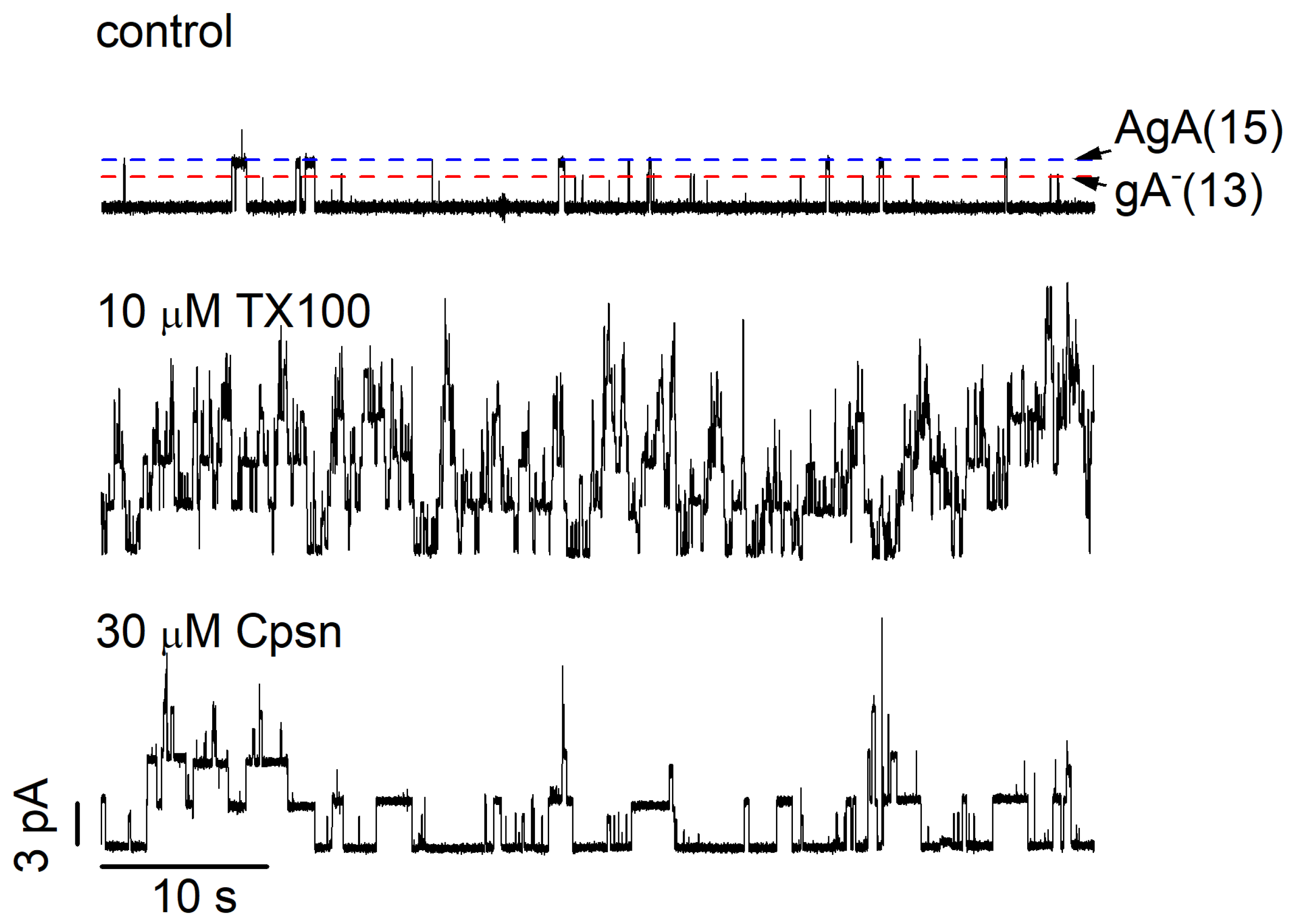
2.2. Amphiphile Modulation of gA Channel Stability
2.3. Comparison of Amphiphile Effects on Alm and gA Channel Function
3. Discussion
4. Materials and Methods
4.1. Materials
4.2. Methods
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| Alm | Alamethicin |
| gA | gramicidin A |
| AgA(15) | [Ala1]gramicidin A |
| gA−(13) | des-Val1,Gly2-gramicidin A |
| Cpsn | Capsaicin |
| TX100 | Triton X-100 |
| rTX100 | reduced Triton X-100 |
References
- Sandermann, H.J. Regulation of membrane enzymes by lipids. Biochim. Biophys. Acta 1978, 515, 209–237. [Google Scholar] [CrossRef]
- Spector, A.A.; Yorek, M.A. Membrane lipid composition and cellular function. J. Lipid Res. 1985, 26, 1015–1035. [Google Scholar] [CrossRef] [PubMed]
- Bienvenüe, A.; Marie, J.S. Modulation of protein function by lipids. Curr. Top. Membr. 1994, 40, 319–354. [Google Scholar]
- Brown, M.F. Modulation of rhodopsin function by properties of the membrane bilayer. Chem. Phys. Lipids 1994, 73, 159–180. [Google Scholar] [CrossRef]
- Lee, A.G. How lipids and proteins interact in a membrane: A molecular approach. Mol. Biosyst. 2005, 1, 203–212. [Google Scholar] [CrossRef] [PubMed]
- Andersen, O.S.; Koeppe, R.E., II. Bilayer thickness and membrane protein function: An energetic perspective. Annu. Rev. Biophys. Biomol. Struct. 2007, 36, 107–130. [Google Scholar] [CrossRef]
- Marsh, D. Protein modulation of lipids, and vice-versa, in membranes. Biochim. Biophys. Acta 2008, 1778, 1545–1575. [Google Scholar] [CrossRef]
- Lundbæk, J.A.; Collingwood, S.A.; Ingólfsson, H.I.; Kapoor, R.; Andersen, O.S. Lipid bilayer regulation of membrane protein function: Gramicidin channels as molecular force probes. J. R. Soc. Interface 2010, 7, 373–395. [Google Scholar] [CrossRef]
- Szlenk, C.T.; GC, J.B.; Natesan, S. Does the Lipid Bilayer Orchestrate Access and Binding of Ligands to Transmembrane Orthosteric/Allosteric Sites of G Protein-Coupled Receptors? Mol. Pharmacol. 2019, 96, 527–541. [Google Scholar] [CrossRef]
- Levental, K.R.; Malmberg, E.; Symons, J.L.; Fan, Y.Y.; Chapkin, R.S.; Ernst, R.; Levental, I. Lipidomic and biophysical homeostasis of mammalian membranes counteracts dietary lipid perturbations to maintain cellular fitness. Nat. Commun. 2020, 11, 1339. [Google Scholar] [CrossRef]
- Payandeh, J.; Volgraf, M. Ligand binding at the protein-lipid interface: Strategic considerations for drug design. Nat. Rev. Drug Discov. 2021, 20, 710–722. [Google Scholar] [CrossRef]
- Levental, I.; Lyman, E. Regulation of membrane protein structure and function by their lipid nano-environment. Nat. Rev. Mol. Cell Biol. 2023, 24, 107–122. [Google Scholar] [CrossRef]
- Ashrafuzzaman, M.; Lampson, M.A.; Greathouse, D.V.; Koeppe, R.E.I.; Andersen, O.S. Manipulating lipid bilayer material properties using biologically active amphipathic molecules. J. Phys. Condens. Matter 2006, 18, S1235–S1255. [Google Scholar] [CrossRef]
- Rusinova, R.; Herold, K.F.; Sanford, R.L.; Greathouse, D.V.; Hemmings, H.C.J.; Andersen, O.S. Thiazolidinedione insulin sensitizers alter lipid bilayer properties and voltage-dependent sodium channel function: Implications for drug discovery. J. Gen. Physiol. 2011, 138, 249–270. [Google Scholar] [CrossRef]
- Nielsen, C.; Andersen, O.S. Inclusion-induced bilayer deformations: Effects of monolayer equilibrium curvature. Biophys. J. 2000, 79, 2583–2604. [Google Scholar] [CrossRef]
- Hatzakis, N.S.; Bhatia, V.K.; Larsen, J.; Madsen, K.L.; Bolinger, P.Y.; Kunding, A.H.; Castillo, J.; Gether, U.; Hedegard, P.; Stamou, D. How curved membranes recruit amphipathic helices and protein anchoring motifs. Nat. Chem. Biol. 2009, 5, 835–841. [Google Scholar] [CrossRef]
- Iversen, L.; Mathiasen, S.; Larsen, J.B.; Stamou, D. Membrane curvature bends the laws of physics and chemistry. Nat. Chem. Biol. 2015, 11, 822–825. [Google Scholar] [CrossRef] [PubMed]
- Nagle, J.F.; Wiener, M.C. Structure of fully hydrated bilayer dispersions. Biochim. Biophys. Acta 1988, 942, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Kucerka, N.; Tristram-Nagle, S.; Nagle, J.F. Structure of fully hydrated fluid phase lipid bilayers with monounsaturated chains. J. Membr. Biol. 2005, 208, 193–202. [Google Scholar] [CrossRef] [PubMed]
- Rostovtseva, T.K.; Petrache, H.I.; Kazemi, N.; Hassanzadeh, E.; Bezrukov, S.M. Interfacial polar interactions affect gramicidin channel kinetics. Biophys. J. 2008, 94, L23–L25. [Google Scholar] [CrossRef] [PubMed]
- Maer, A.M.; Rusinova, R.; Providence, L.L.; Ingólfsson, H.I.; Collingwood, S.A.; Lundbæk, J.A.; Andersen, O.S. Regulation of Gramicidin Channel Function Solely by Changes in Lipid Intrinsic Curvature. Front. Physiol. 2022, 13, 836789. [Google Scholar] [CrossRef]
- Jung, D.; Powers, J.P.; Straus, S.K.; Hancock, R.E. Lipid-specific binding of the calcium-dependent antibiotic daptomycin leads to changes in lipid polymorphism of model membranes. Chem. Phys. Lipids 2008, 154, 120–128. [Google Scholar] [CrossRef] [PubMed]
- Hakizimana, P.; Masureel, M.; Gbaguidi, B.; Ruysschaert, J.M.; Govaerts, C. Interactions between phosphatidylethanolamine headgroup and LmrP, a multidrug transporter: A conserved mechanism for proton gradient sensing? J. Biol. Chem. 2008, 283, 9369–9376. [Google Scholar] [CrossRef] [PubMed]
- Lundbæk, J.A.; Birn, P.; Tape, S.E.; Toombes, G.E.; Søgaard, R.; Koeppe, R.E., II; Gruner, S.M.; Hansen, A.J.; Andersen, O.S. Capsaicin regulates voltage-dependent sodium channels by altering lipid bilayer elasticity. Mol. Pharmacol. 2005, 68, 680–689. [Google Scholar] [CrossRef] [PubMed]
- Evans, E.; Rawicz, W.; Hofmann, A.F. Lipid bilayer expansion and mechanical disruption in solutions of water-soluble bile acid. In Bile Acids in Gastroenterology: Basic and Clinical Advances; Hofmann, A.F., Paumgartner, G., Stiehl, A., Eds.; Kluwer Academic Publishers: Dordrecht, The Netherlands, 1995; pp. 59–68. [Google Scholar]
- Zhelev, D.V. Material property characteristics for lipid bilayers containing lysolipid. Biophys. J. 1998, 75, 321–330. [Google Scholar] [CrossRef] [PubMed]
- Bruno, M.J.; Koeppe, R.E., II; Andersen, O.S. Docosahexaenoic acid alters bilayer elastic properties. Proc. Natl. Acad. Sci. USA 2007, 104, 9638–9643. [Google Scholar] [CrossRef] [PubMed]
- Bruno, M.J.; Rusinova, R.; Gleason, N.J.; Koeppe, R.E., II; Andersen, O.S. Interactions of drugs and amphiphiles with membranes: Modulation of lipid bilayer elastic properties by changes in acyl chain unsaturation and protonation. Faraday Disc. 2013, 161, 461–480. [Google Scholar] [CrossRef]
- Keller, S.L.; Bezrukov, S.M.; Gruner, S.M.; Tate, M.W.; Vodyanoy, I.; Parsegian, V.A. Probability of alamethicin conductance states varies with nonlamellar tendency of bilayer phospholipids. Biophys. J. 1993, 65, 23–27. [Google Scholar] [CrossRef]
- Lundbæk, J.A.; Maer, A.M.; Andersen, O.S. Lipid bilayer electrostatic energy, curvature stress, and assembly of gramicidin channels. Biochemistry 1997, 36, 5695–5701. [Google Scholar] [CrossRef]
- Lundbæk, J.A.; Birn, P.; Girshman, J.; Hansen, A.J.; Andersen, O.S. Membrane stiffness and channel function. Biochemistry 1996, 35, 3825–3830. [Google Scholar] [CrossRef]
- Kleinkauf, H.; Von Döhren, H. A nonribosomal system of peptide biosynthesis. Eur. J. Biochem. 1996, 236, 335–351. [Google Scholar] [CrossRef] [PubMed]
- Bezrukov, S.M. Functional consequences of lipid packing stress. Curr. Opin. Colloid Interface Sci. 2000, 5, 237–243. [Google Scholar] [CrossRef]
- Sarges, R.; Witkop, B.; Gramicidin, A.V. The structure of valine- and isoleucine-gramicidin A. J. Am. Chem. Soc. 1965, 87, 2011–2019. [Google Scholar] [CrossRef]
- Abo-Riziq, A.; Crews, B.O.; Callahan, M.P.; Grace, L.; de Vries, M.S. Spectroscopy of isolated gramicidin peptides. Angew. Chem. Int. Ed. Engl. 2006, 45, 5166–5169. [Google Scholar] [CrossRef]
- O’Connell, A.M.; Koeppe, R.E., II; Andersen, O.S. Kinetics of gramicidin channel formation in lipid bilayers: Transmembrane monomer association. Science 1990, 250, 1256–1259. [Google Scholar] [CrossRef]
- Rinehart, K.L.J.; Cook, J.C.J.; Meng, H.; Olson, K.L.; Pandey, R.C. Mass spectrometric determination of molecular formulas for membrane-modifying antibiotics. Nature 1977, 269, 832–833. [Google Scholar] [CrossRef]
- Huang, H.W. Molecular mechanism of antimicrobial peptides: The origin of cooperativity. Biochim. Biophys. Acta 2006, 1758, 1292–1302. [Google Scholar] [CrossRef] [PubMed]
- Bamberg, E.; Läuger, P. Channel formation kinetics of gramicidin A in lipid bilayer membranes. J. Membr. Biol. 1973, 11, 177–194. [Google Scholar] [CrossRef]
- Zingsheim, H.P.; Neher, E. The equivalence of fluctuation analysis and chemical relaxation measurements: A kinetic study of ion pore formation in thin lipid membranes. Biophys. Chem. 1974, 2, 197–207. [Google Scholar] [CrossRef]
- Veatch, W.R.; Mathies, R.; Eisenberg, M.; Stryer, L. Simultaneous fluorescence and conductance studies of planar bilayer membranes containing a highly active and fluorescent analog of gramicidin A. J. Mol. Biol. 1975, 99, 75–92. [Google Scholar] [CrossRef]
- Hickok, N.J.; Kustin, K.; Veatch, W. Relaxation spectra of gramicidin dimerization in a lipid bilayer membrane. Biochim. Biophys. Acta 1986, 858, 99–106. [Google Scholar] [CrossRef] [PubMed]
- Rokitskaya, T.I.; Antonenko, Y.N.; Kotova, E.A. Photodynamic inactivation of gramicidin channels: A flash-photolysis study. Biochim. Biophys. Acta 1996, 1275, 221–226. [Google Scholar] [CrossRef] [PubMed]
- Lum, K.; Ingólfsson, H.I.; Koeppe, R.E., II; Andersen, O.S. Exchange of Gramicidin between Lipid Bilayers: Implications for the Mechanism of Channel Formation. Biophys. J. 2017, 113, 1757–1767. [Google Scholar] [CrossRef] [PubMed]
- Arseniev, A.S.; Barsukov, I.L.; Bystrov, V.F.; Ovchinnikov, Y.A. Spatial structure of a gramicidin A transmembrane ion channel. NMR analysis in micelles. Biol. Membr. 1986, 3, 437–462. [Google Scholar]
- Townsley, L.E.; Tucker, W.A.; Sham, S.; Hinton, J.F. Structures of gramicidins A, B, and C incorporated into sodium dodecyl sulfate micelles. Biochemistry 2001, 40, 11676–11686. [Google Scholar] [CrossRef] [PubMed]
- Ketchem, R.R.; Hu, W.; Cross, T.A. High-resolution conformation of gramicidin A in a lipid bilayer by solid-state NMR. Science 1993, 261, 1457–1460. [Google Scholar] [CrossRef] [PubMed]
- Ketchem, R.; Roux, B.; Cross, T. High-resolution polypeptide structure in a lamellar phase lipid environment from solid state NMR derived orientational constraints. Structure 1997, 5, 1655–1669. [Google Scholar] [CrossRef]
- Allen, T.W.; Andersen, O.S.; Roux, B. Structure of gramicidin a in a lipid bilayer environment determined using molecular dynamics simulations and solid-state NMR data. J. Am. Chem. Soc. 2003, 125, 9868–9877. [Google Scholar] [CrossRef]
- Greathouse, D.V.; Hinton, J.F.; Kim, K.S.; Koeppe, R.E., II. Gramicidin A/short-chain phospholipid dispersions: Chain length dependence of gramicidin conformation and lipid organization. Biochemistry 1994, 33, 4291–4299. [Google Scholar] [CrossRef]
- Mobashery, N.; Nielsen, C.; Andersen, O.S. The conformational preference of gramicidin channels is a function of lipid bilayer thickness. FEBS Lett. 1997, 412, 15–20. [Google Scholar] [CrossRef]
- Galbraith, T.P.; Wallace, B.A. Phospholipid chain length alters the equilibrium between pore and channel forms of gramicidin. Faraday Discuss. 1998, 111, 159–164; discussion 225–246. [Google Scholar] [CrossRef] [PubMed]
- Höfer, N.; Aragão, D.; Caffrey, M. Crystallizing transmembrane peptides in lipidic mesophases. Biophys. J. 2010, 99, L23–L25. [Google Scholar] [CrossRef] [PubMed]
- Wallace, B.A.; Veatch, W.R.; Blout, E.R. Conformation of gramicidin A in phospholipid vesicles: Circular dichroism studies of effects of ion binding, chemical modification, and lipid structure. Biochemistry 1981, 20, 5754–5760. [Google Scholar] [CrossRef] [PubMed]
- Katsaras, J.; Prosser, R.S.; Stinson, R.H.; Davis, J.H. Constant helical pitch of the gramicidin channel in phospholipid bilayers. Biophys. J. 1992, 61, 827–830. [Google Scholar] [CrossRef] [PubMed]
- Busath, D.D.; Andersen, O.S.; Koeppe, R.E., II. On the conductance heterogeneity in membrane channels formed by gramicidin A. A cooperative study. Biophys. J. 1987, 51, 79–88. [Google Scholar] [CrossRef] [PubMed]
- Hladky, S.B.; Haydon, D.A. Ion transfer across lipid membranes in the presence of gramicidin A. I. Studies of the unit conductance channel. Biochim. Biophys. Acta 1972, 274, 294–312. [Google Scholar] [CrossRef] [PubMed]
- Kolb, H.A.; Bamberg, E. Influence of membrane thickness and ion concentration on the properties of the gramicidin a channel. Autocorrelation, spectral power density, relaxation and single-channel studies. Biochim. Biophys. Acta. 1977, 464, 127–141. [Google Scholar] [CrossRef]
- Elliott, J.R.; Needham, D.; Dilger, J.P.; Haydon, D.A. The effects of bilayer thickness and tension on gramicidin single-channel lifetime. Biochim. Biophys. Acta 1983, 735, 95–103. [Google Scholar] [CrossRef]
- Huang, H.W. Deformation free energy of bilayer membrane and its effect on gramicidin channel lifetime. Biophys. J. 1986, 50, 1061–1070. [Google Scholar] [CrossRef]
- Beaven, A.H.; Maer, A.M.; Sodt, A.J.; Rui, H.; Pastor, R.W.; Andersen, O.S.; Im, W. Gramicidin A Channel Formation Induces Local Lipid Redistribution I: Experiment and Simulation. Biophys. J. 2017, 112, 1185–1197. [Google Scholar] [CrossRef]
- Sodt, A.J.; Beaven, A.H.; Andersen, O.S.; Im, W.; Pastor, R.W. Gramicidin A Channel Formation Induces Local Lipid Redistribution II: A 3D Continuum Elastic Model. Biophys. J. 2017, 112, 1198–1213. [Google Scholar] [CrossRef] [PubMed]
- Gordon, L.G.M.; Haydon, D.A. The unit conductance channel of alamethicin. Biochim. Biophys. Acta 1972, 255, 1014–1018. [Google Scholar] [CrossRef] [PubMed]
- Boheim, G. Statistical analysis of alamethicin channels in black lipid membranes. J. Membr. Biol. 1974, 19, 277–303. [Google Scholar] [CrossRef] [PubMed]
- Latorre, R.; Alvarez, O. Voltage-dependent channels in planar lipid bilayer membranes. Physiol. Rev. 1981, 61, 77–150. [Google Scholar] [CrossRef] [PubMed]
- Woolley, G.A.; Dunn, A.; Wallace, B.A. Gramicidin-lipid interactions induce specific tryptophan side-chain conformations. Biochem. Soc. Trans. 1992, 20, 864–867. [Google Scholar] [CrossRef] [PubMed]
- Cafiso, D.S. Alamethicin: A peptide model for voltage gating and protein-membrane interactions. Annu. Rev. Biophys. Biomol. Struct. 1994, 23, 141–165. [Google Scholar] [CrossRef] [PubMed]
- Bechinger, B. The structure, dynamics and orientation of antimicrobial peptides in membranes by multidimensional solid-state NMR spectroscopy. Biochim. Biophys. Acta 1999, 1462, 157–183. [Google Scholar] [CrossRef] [PubMed]
- Fox, R.O.J.; Richards, F.M. A voltage-gated ion channel model inferred from the crystal structure of alamethicin at 1.5-Å resolution. Nature 1982, 300, 325–330. [Google Scholar] [CrossRef]
- Archer, S.J.; Ellena, J.F.; Cafiso, D.S. Dynamics and aggregation of the peptide ion channel alamethicin. Measurements using spin-labeled peptides. Biophys. J. 1991, 60, 389–398. [Google Scholar] [CrossRef]
- Huang, H.W.; Wu, Y. Lipid-alamethicin interactions influence alamethicin orientation. Biophys. J. 1991, 60, 1079–1087. [Google Scholar] [CrossRef]
- Chen, F.Y.; Lee, M.T.; Huang, H.W. Evidence for membrane thinning effect as the mechanism for peptide-induced pore formation. Biophys. J. 2003, 84, 3751–3758. [Google Scholar] [CrossRef]
- Lewis, J.R.; Cafiso, D.S. Correlation between the free energy of a channel-forming voltage-gated peptide and the spontaneous curvature of bilayer lipids. Biochemistry 1999, 38, 5932–5938. [Google Scholar] [CrossRef]
- Huang, H.W. Elasticity of lipid bilayer interacting with amphiphilic helical peptides. J. Phys. II 1995, 5, 1427–1431. [Google Scholar] [CrossRef]
- He, K.; Ludtke, S.J.; Worcester, D.L.; Huang, H.W. Neutron scattering in the plane of membranes: Structure of alamethicin pores. Biophys. J. 1996, 70, 2659–2666. [Google Scholar] [CrossRef]
- Qian, S.; Wang, W.; Yang, L.; Huang, H.W. Structure of the alamethicin pore reconstructed by x-ray diffraction analysis. Biophys. J. 2008, 94, 3512–3522. [Google Scholar] [CrossRef]
- Pan, J.; Tieleman, D.P.; Nagle, J.F.; Kucerka, N.; Tristram-Nagle, S. Alamethicin in lipid bilayers: Combined use of X-ray scattering and MD simulations. Biochim. Biophys. Acta 2009, 1788, 1387–1397. [Google Scholar] [CrossRef]
- Baumann, G.; Mueller, P. A molecular model of membrane excitability. J. Supramol. Struct. 1974, 2, 538–557. [Google Scholar] [CrossRef]
- Opsahl, L.R.; Webb, W.W. Transduction of membrane tension by the ion channel alamethicin. Biophys. J. 1994, 66, 71–74. [Google Scholar] [CrossRef] [PubMed]
- Bezrukov, S.M.; Vodyanoy, I. Probing alamethicin channels with water-soluble polymers. Effect on conductance of channel states. Biophys. J. 1993, 64, 16–25. [Google Scholar] [CrossRef] [PubMed]
- He, K.; Ludtke, S.J.; Huang, H.W.; Worcester, D.L. Antimicrobial peptide pores in membranes detected by neutron in-plane scattering. Biochemistry 1995, 34, 15614–15618. [Google Scholar] [CrossRef] [PubMed]
- Bak, M.; Bywater, R.P.; Hohwy, M.; Thomsen, J.K.; Adelhorst, K.; Jakobsen, H.J.; Sorensen, O.W.; Nielsen, N.C. Conformation of alamethicin in oriented phospholipid bilayers determined by 15N solid-state nuclear magnetic resonance. Biophys. J. 2001, 81, 1684–1698. [Google Scholar] [CrossRef]
- Rizzo, V.; Schwarz, G.; Voges, K.P.; Jung, G. Molecular shape and dipole moment of alamethicin-like synthetic peptides. Eur. Biophys. J. 1985, 12, 67–73. [Google Scholar] [CrossRef] [PubMed]
- Hanke, W.; Boheim, G. The lowest conductance state of the alamethicin pore. Biochim. Biophys. Acta 1980, 596, 456–462. [Google Scholar] [CrossRef] [PubMed]
- Bezrukov, S.M.; Rand, R.P.; Vodyanoy, I.; Parsegian, V.A. Lipid packing stress and polypeptide aggregation: Alamethicin channel probed by proton titration of lipid charge. Faraday Discuss. 1998, 111, 173–183; discussion 225–246. [Google Scholar] [CrossRef] [PubMed]
- Ly, H.V.; Longo, M.L. The influence of short-chain alcohols on interfacial tension, mechanical properties, area/molecule, and permeability of fluid lipid bilayers. Biophys. J. 2004, 87, 1013–1033. [Google Scholar] [CrossRef]
- Lundbæk, J.A.; Koeppe, R.E., II; Andersen, O.S. Amphiphile regulation of ion channel function by changes in the bilayer spring constant. Proc. Natl. Acad. Sci. USA 2010, 107, 15427–15430. [Google Scholar] [CrossRef]
- Kapoor, R.; Peyear, T.A.; Koeppe, R.E., II; Andersen, O.S. Antidepressants are modifiers of lipid bilayer properties. J. Gen. Physiol. 2019, 151, 342–356. [Google Scholar] [CrossRef] [PubMed]
- Lundbæk, J.A.; Birn, P.; Hansen, A.J.; Søgaard, R.; Nielsen, C.; Girshman, J.; Bruno, M.J.; Tape, S.E.; Egebjerg, J.; Greathouse, D.V.; et al. Regulation of sodium channel function by bilayer elasticity: The importance of hydrophobic coupling. Effects of Micelle-forming amphiphiles and cholesterol. J. Gen. Physiol. 2004, 123, 599–621. [Google Scholar] [CrossRef] [PubMed]
- Søgaard, R.; Werge, T.M.; Bertelsen, C.; Lundbye, C.; Madsen, K.L.; Nielsen, C.H.; Lundbæk, J.A. GABAA receptor function is regulated by lipid bilayer elasticity. Biochemistry 2006, 45, 13118–13129. [Google Scholar] [CrossRef]
- Andersen, O.S.; Finkelstein, A.; Katz, I.; Cass, A. Effect of phloretin on the permeability of thin lipid membranes. J. Gen. Physiol. 1976, 67, 749–771. [Google Scholar] [CrossRef]
- Duffin, R.L.; Garrett, M.P.; Busath, D.D. Modulation of Lipid Bilayer Interfacial Dipole Potential by Phloretin, RH421, and 6-Ketocholestanol as Probed by Gramicidin Channel Conductance. Langmuir 2003, 19, 1439–1442. [Google Scholar] [CrossRef]
- Mereuta, L.; Asandei, A.; Luchian, T. Meet me on the other side: Trans-bilayer modulation of a model voltage-gated ion channel activity by membrane electrostatics asymmetry. PLoS ONE 2011, 6, e25276. [Google Scholar] [CrossRef]
- Colquhoun, D.; Sigworth, F.J. Fitting and statistical analysis of single-channel records. In Single-Channel Recording, 2nd ed.; Sakmann, B., Neher, E., Eds.; Plenum Press: New York, NY, USA, 1995; pp. 483–587. [Google Scholar]
- Sawyer, D.B.; Koeppe, R.E., II; Andersen, O.S. Induction of conductance heterogeneity in gramicidin channels. Biochemistry 1989, 28, 6571–6583. [Google Scholar] [CrossRef]
- Andersen, O.S. Ion movement through gramicidin A channels. Single-channel measurements at very high potentials. Biophys. J. 1983, 41, 119–133. [Google Scholar] [CrossRef] [PubMed]
- Ingólfson, H.; Kapoor, R.; Collingwood, S.A.; Andersen, O.S. Single molecule methods for monitoring changes in bilayer elastic properties. J. Vis. Exp. 2008, 21, e1032. [Google Scholar]
- Artigas, P.; Al’aref, S.J.; Hobart, E.A.; Diaz, L.F.; Sakaguchi, M.; Straw, S.; Andersen, O.S. 2,3-Butanedione monoxime affects Cystic Fibrosis Transmembrane Conductance Regulator channel function through phosphorylation-dependent and phosphorylation-independent mechanisms: The role of bilayer material properties. Mol. Pharmacol. 2006, 70, 2015–2026. [Google Scholar] [CrossRef]
- Nielsen, C.; Goulian, M.; Andersen, O.S. Energetics of inclusion-induced bilayer deformations. Biophys. J. 1998, 74, 1966–1983. [Google Scholar] [CrossRef]
- Lundbæk, J.A.; Andersen, O.S. Spring constants for channel-induced lipid bilayer deformations—Estimates using gramicidin channels. Biophys. J. 1999, 76, 889–895. [Google Scholar] [CrossRef]
- White, S.H.; Thompson, T.E. Capacitance, area, and thickness variations in thin lipid films. Biochim. Biophys. Acta 1973, 323, 7–22. [Google Scholar] [CrossRef] [PubMed]
- White, S.H. Comments on “electrical breakdown of bimolecular lipid membranes as an electromechanical instability”. Biophys. J. 1974, 14, 155–158. [Google Scholar] [CrossRef]
- Evans, E.A.; Simon, S. Mechanics of electrocompression of lipid bilayer membranes. Biophys. J. 1975, 15, 850–852. [Google Scholar] [CrossRef]
- Requena, J.; Haydon, D.A.; Hladky, S.B. Lenses and the compression of black lipid membranes by an electric field. Biophys. J. 1975, 15, 77–81. [Google Scholar]
- Rusinova, R.; Koeppe, R.E., II; Andersen, O.S. A general mechanism for drug promiscuity: Studies with amiodarone and other antiarrhythmics. J. Gen. Physiol. 2015, 146, 463–475. [Google Scholar] [CrossRef] [PubMed]
- Tate, M.W.; Eikenberry, E.F.; Turner, D.C.; Shyamsunder, E.; Gruner, S.M. Nonbilayer phases of membrane lipids. Chem. Phys. Lipids 1991, 57, 147–164. [Google Scholar] [CrossRef]
- McLaughlin, S. Salicylates and phospholipid bilayer membranes. Nature 1973, 243, 234–236. [Google Scholar] [CrossRef] [PubMed]
- McLaughlin, S.; Harary, H. The hydrophobic adsorption of charged molecules to bilayer membranes: A test of the applicability of the Stern equation. Biochemistry 1976, 15, 1941–1948. [Google Scholar] [CrossRef] [PubMed]
- Schwarz, G.; Stankowski, S.; Rizzo, V. Thermodynamic analysis of incorporation and aggregation in a membrane: Application to the pore-forming peptide alamethicin. Biochim. Biophys. Acta 1986, 861, 141–151. [Google Scholar] [CrossRef] [PubMed]
- Keller, S.L.; Gruner, S.M.; Gawrisch, K. Small concentrations of alamethicin induce a cubic phase in bulk phosphatidylethanolamine mixtures. Biochim. Biophys. Acta 1996, 1278, 241–246. [Google Scholar] [CrossRef] [PubMed]
- He, K.; Ludtke, S.J.; Heller, W.T.; Huang, H.W. Mechanism of alamethicin insertion into lipid bilayers. Biophys. J. 1996, 71, 2669–2679. [Google Scholar] [CrossRef]
- Pan, J.; Tristram-Nagle, S.; Nagle, J.F. Alamethicin aggregation in lipid membranes. J. Membr. Biol. 2009, 231, 11–27. [Google Scholar] [CrossRef]
- Greathouse, D.V.; Koeppe, R.E., II; Providence, L.L.; Shobana, S.; Andersen, O.S. Design and characterization of gramicidin channels. Methods Enzymol. 1999, 294, 525–550. [Google Scholar] [PubMed]
- Szabo, G.; Eisenman, G.; Ciani, S. The effects of macrotetralide actin antibiotics on the electrical properties of phospholipid bilayer membranes. J. Membr. Biol. 1969, 1, 346–382. [Google Scholar] [CrossRef] [PubMed]
- Kapoor, R.; Kim, J.H.; Ingolfson, H.; Andersen, O.S. Preparation of artificial bilayers for electrophysiology experiments. J. Vis. Exp. 2008, 20, e1033. [Google Scholar]
- de Kroon, A.I.P.M.; Timmermans, J.W.; Killian, J.A.; de Kruijff, B. The pH dependence of headgroup and acyl chain structure and dynamics of phosphatidylserine, studied by 2H-NMR. Chem. Phys. Lipids 1990, 54, 33–42. [Google Scholar] [CrossRef]
- Fuller, N.; Benatti, C.R.; Rand, R.P. Curvature and bending constants for phosphatidylserine-containing membranes. Biophys. J. 2003, 85, 1667–1674. [Google Scholar] [CrossRef]
- Aguilella, V.M.; Bezrukov, S.M. Alamethicin channel conductance modified by lipid charge. Eur. Biophys. J. 2001, 30, 233–241. [Google Scholar]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Level 0 (pA) | Level 1 (pA) | Level 2 (pA) | Level 3 (pA) | |
|---|---|---|---|---|
| DOPC | 4.5 ± 0.2 | 18.0 ± 0.5 | 38.4 ± 0.8 | 61 ± 1 |
| +30 µM Cpsn | 4.4 ± 0.3 | 17.7 ± 0.8 | 37.8 ± 0.9 | 60 ± 1 |
| +30 µM TX100 | 4.5 ± 0.1 | 18.6 ± 0.6 | 39.5 ± 0.3 | 60 ± 1 |
| +30 µM rTX100 | 4.5 ± 0.2 | 16.7 ± 0.6 | 36.8 ± 0.8 | 59 ± 1 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ashrafuzzaman, M.; Koeppe, R.E., II; Andersen, O.S. Intrinsic Lipid Curvature and Bilayer Elasticity as Regulators of Channel Function: A Comparative Single-Molecule Study. Int. J. Mol. Sci. 2024, 25, 2758. https://doi.org/10.3390/ijms25052758
Ashrafuzzaman M, Koeppe RE II, Andersen OS. Intrinsic Lipid Curvature and Bilayer Elasticity as Regulators of Channel Function: A Comparative Single-Molecule Study. International Journal of Molecular Sciences. 2024; 25(5):2758. https://doi.org/10.3390/ijms25052758
Chicago/Turabian StyleAshrafuzzaman, Mohammad, Roger E. Koeppe, II, and Olaf S. Andersen. 2024. "Intrinsic Lipid Curvature and Bilayer Elasticity as Regulators of Channel Function: A Comparative Single-Molecule Study" International Journal of Molecular Sciences 25, no. 5: 2758. https://doi.org/10.3390/ijms25052758