
Sulfur-Oxidizing Bacteria Alleviate Salt and Cadmium Stress in Halophyte Tripolium pannonicum (Jacq.) Dobrocz.


Abstract
:1. Introduction
2. Results
2.1. Effect on Growth Parameters
2.2. Effect on Biochemical Parameters
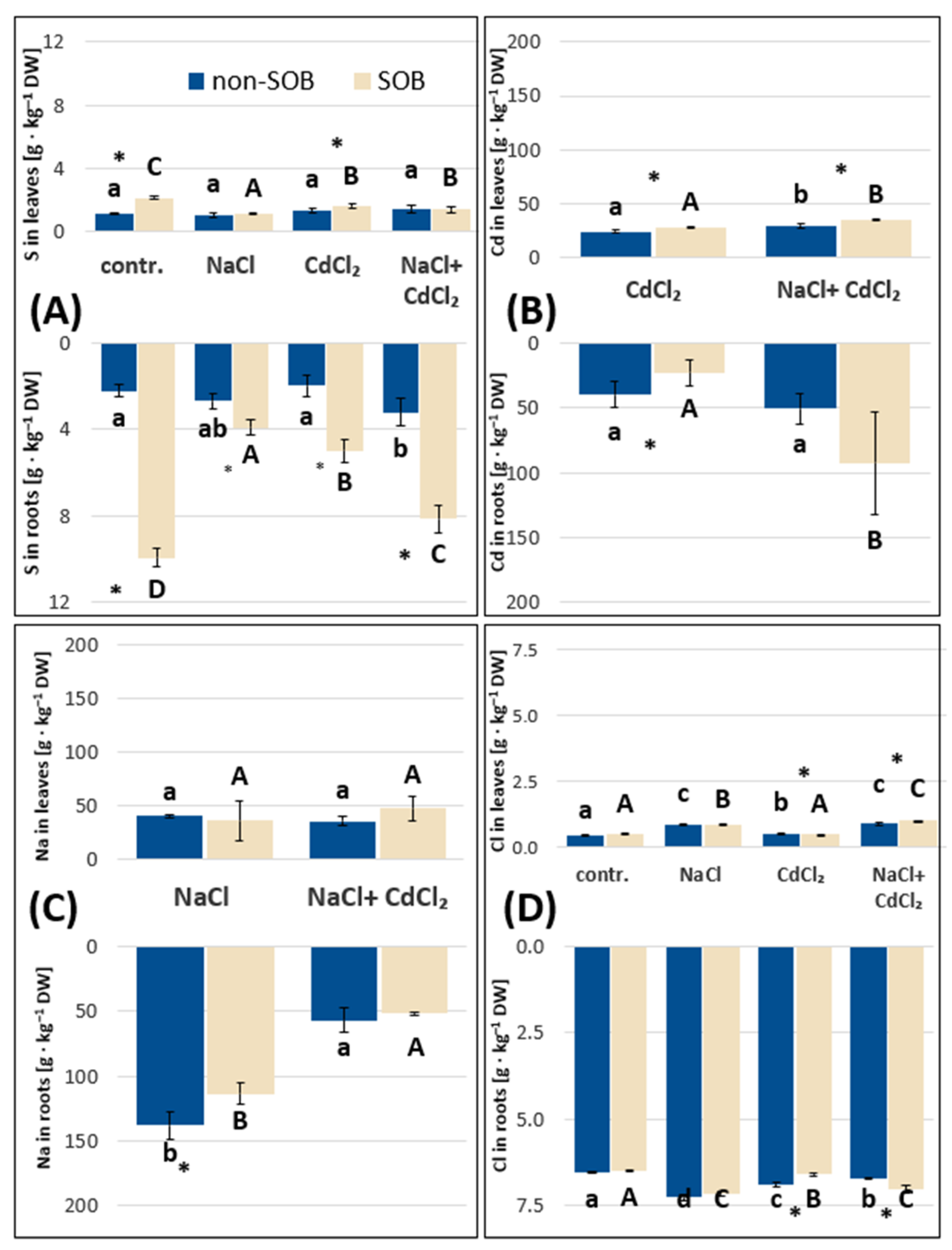
2.2.1. Sulfur, Sodium, Chloride and Cadmium Absorption by Tripolium pannonicum Plants
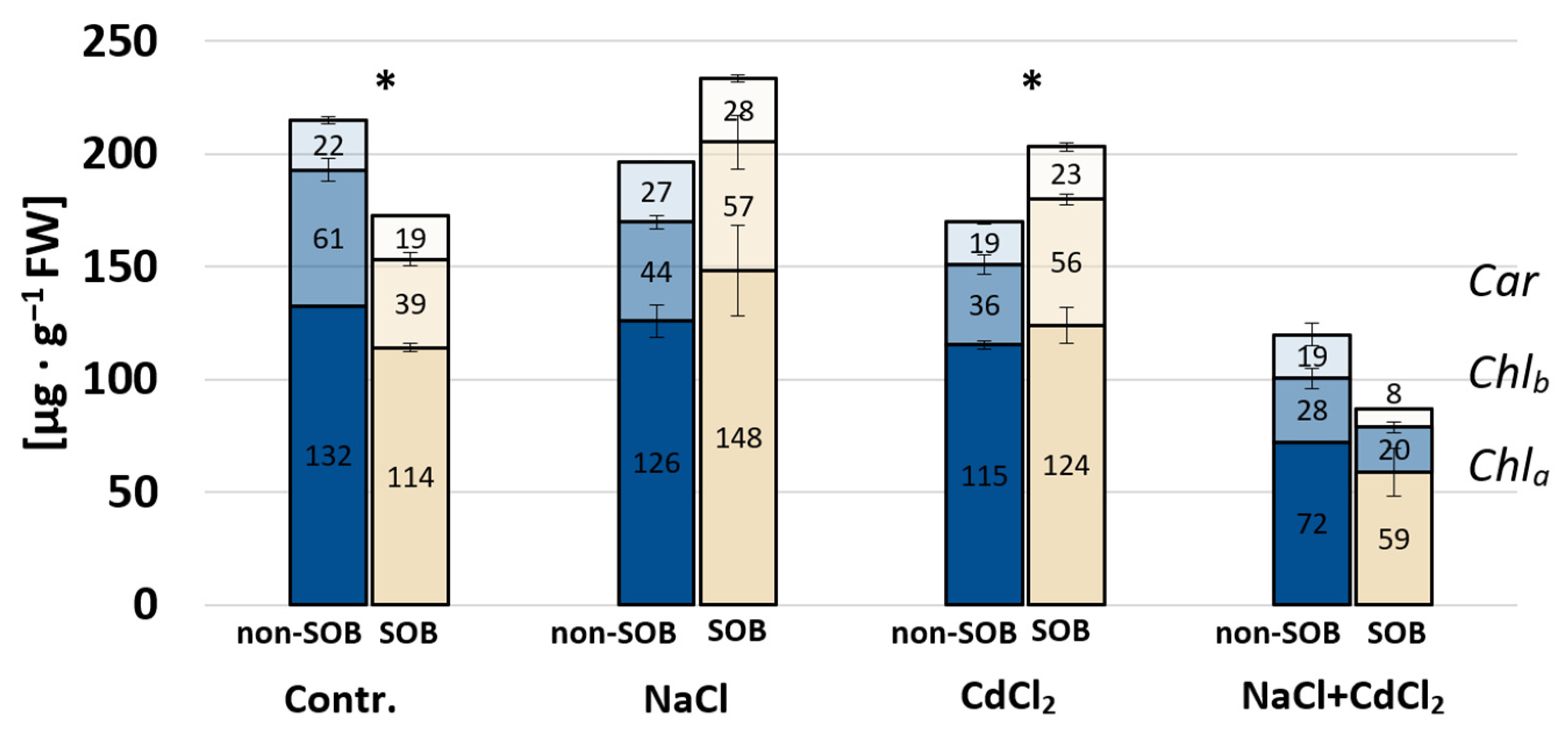
2.2.2. Photosynthetic Pigments
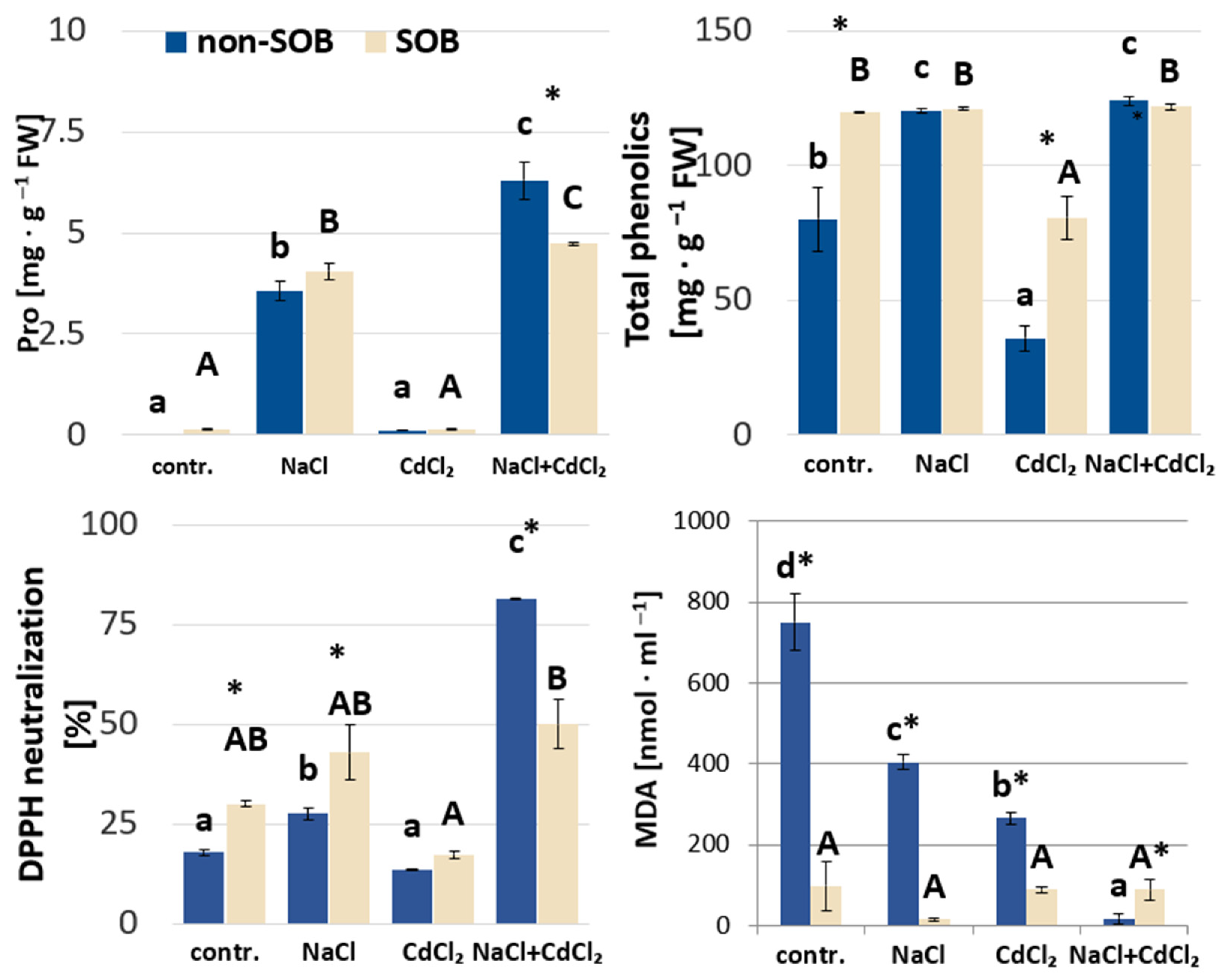
2.2.3. Proline, Phenolic Compounds, DPPH Radical and Lipid Peroxidation
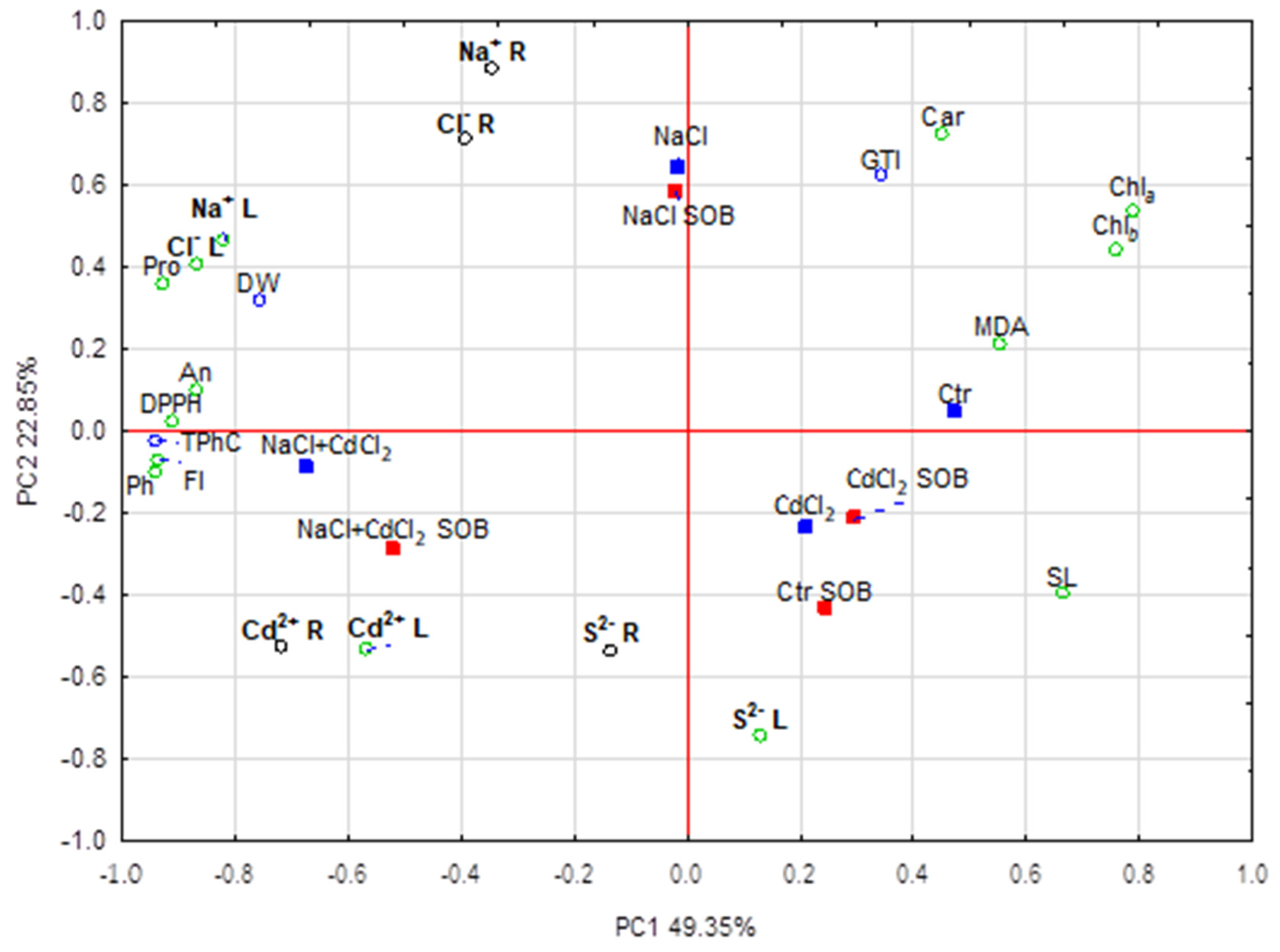
2.3. Multivariate Analysis
3. Discussion
4. Materials and Methods
4.1. Experimental Design
Bacterial Strains
4.2. Plant Growth Parameters
4.3. Concentrations of Sulfur, Sodium, Chloride and Cadmium in Plants
4.4. Plant Biochemical Analyses
4.4.1. Photosynthetic Pigments
4.4.2. Proline
4.4.3. Phenolic Compounds
4.4.4. Phenolic Profile
4.4.5. Radical Scavenging Activity with DPPH Radical
4.4.6. Lipid Peroxidation
4.5. Statistics
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Acosta, J.A.; Jansen, B.; Kalbitz, K.; Faz, A.; Martínez-Martínez, S. Salinity increases mobility of heavy metals in soils. Chemosphere 2011, 85, 1318–1324. [Google Scholar] [CrossRef] [PubMed]
- Sristava, V.; Sarkar, A.; Singh, S.; Singh, P.; de Aruajo, A.S.F.; Singh, R.P. Agoecological Responses of heavy metal pollution with special emphasis on soil health and plant performances. Front. Environ. Sci. 2017, 5, 64. [Google Scholar] [CrossRef]
- Ghori, N.H.; Ghori, T.; Hayat, M.Q.; Imadi, S.R.; Gul, A.; Altay, V.; Ozturk, M. Heavy metal stress responses in plants. Int. J. Environ. Sci. Technol. 2019, 16, 1807–1828. [Google Scholar] [CrossRef]
- Ibraheem, F.; Al-Zahrani, A.; Mosa, A. Physiological Adaptation of Three Wild halophytic Suaeda Species: Salt Tolerance Strategies and Metal Accumulation Capacity. Plants 2022, 11, 357. [Google Scholar] [CrossRef] [PubMed]
- Rahman, M.M.; Mostofa, M.G.; Keya, S.S.; Siddiqui, M.N.; Ansary, M.M.U.; Das, A.K.; Rahman, M.A.; Tran, L.S.-P. Adaptative Mechanisms of Halophytes and Their Potential in Improving Salinity Tolerance in Plants. Int. J. Mol. Sci. 2021, 22, 10733. [Google Scholar] [CrossRef] [PubMed]
- Van Oosten, J.M.; Maggio, A. Functional biology of halophytes in the phytoremediation of heavy metal contaminated soils. Environ. Exp. Bot. 2015, 111, 135–146. [Google Scholar] [CrossRef]
- Bankaji, I.; Cacador, I.; Sleimi, N. Assessing of tolerance to metallic and saline stresses in the halophyte Suaeda fruticosa: The indicator role of antioxidanr enzymes. Ecol. Indicat. 2016, 64, 297–308. [Google Scholar] [CrossRef]
- Wiszniewska, A.; Koźmińska, A.; Hanus-Fajerska, E.; Dziurka, M.; Dziurka, K. Insight into mechanisms of multiple stresses tolerance in a halophyte Aster tripolium subjected to salinity and heavy metal stress. Ecotoxicol. Environ. Saf. 2019, 180, 12–22. [Google Scholar] [CrossRef]
- Flowers, T.J.; Colmer, T.D. Plant salt tolerance: Adaptations in halophytes. Ann. Bot. 2015, 115, 327–331. [Google Scholar] [CrossRef] [PubMed]
- Kaur, G.; Asthi, B. Proline: A key player in abiotic stress tolerance. Biol. Plant. 2015, 59, 609–619. [Google Scholar] [CrossRef]
- Ghanem, A.F.M.; Mohamed, E.; Kasem, A.M.M.A.; El-Ghamery, A.A. Different Salt Tolerance Strategies in Three Halophytes from the Same Ecological habitat: Augmentation of Antioxidant Enzymes and Compounds. Plants 2021, 10, 1100. [Google Scholar] [CrossRef]
- Hamed, K.B.; Ellouzi, H.; Talbi, O.Z.; Hessini, K.; Slama, I.; Ghnaya, T.; Bosch, S.M.; Savouré, A.; Abdelly, C. Physiological response of halophytes to multiple stresses. Funct. Plant Biol. 2013, 40, 883–896. [Google Scholar] [CrossRef]
- Ellouzi, H.; Ben Hamed, K.; Asensi-Fabado, M.A.; Müller, M.; Abdelly, C.; Munné-Bosch, S. Drought and cadmium may be as effective as salinity in conferring subsequent salt stress tolerance in Cakile maritima. Planta 2013, 237, 1311–1323. [Google Scholar] [CrossRef]
- Mohamed, R.A.; Khali, W.F.; Zaghloul, M.S. Exoloring the Physiological and Molecular Mechanisms of Halophytes Adaptation to High Salinity Environments: Implications for Enhancing Plant Salinity Tolerance. Catrina 2023, 28, 93–107. [Google Scholar]
- Krämer, U. Metal hyperaccumulation in Plants. Ann. Rev. Plant Biol. 2010, 61, 517–534. [Google Scholar] [CrossRef]
- Fatma, M.; Khan, M.I.R.; Masood, A.; Khan, N.A. Coordinate changes in assimilatory sulfate reduction are correlated to salt tolerance: Involvement of phytohormones. Annu. Rev. Mater. Res. 2013, 33, 267–295. [Google Scholar]
- Kosakivska, I.V.; Babenko, L.M.; Romanenko, K.O.; Korotka, I.Y.; Potters, G. Molecular mechanisms of plant adaptative responses to heavy metal stress. Cell Biol. Int. 2021, 45, 258–272. [Google Scholar] [CrossRef]
- Bashir, H.; Ibrahim, M.M.; Bagheri, R.; Ahmad, J.; Arif, I.A.; Baig, M.A.; Qureshi, M.I. Influence of sulfur and cadmium on antioxidants, phytochelatins and growth in Indian mustard. AoB Plants 2015, 7, plv001. [Google Scholar] [CrossRef]
- Corpas, F.J.; Barroso, J.B. Reactive sulfur species (RSS): Possible new players in the oxidative metabolism of plant peroxisomes. Front. Plant Sci. 2015, 6, 116. [Google Scholar] [CrossRef]
- Fatma, M.; Asgher, M.; Masood, A.; Khan, N.A. Excess sulfur sup- plementation improves photosynthesis and growth in mustard under salt stress through increased production of glutathione. Environ. Exp. Bot. 2014, 107, 55–63. [Google Scholar] [CrossRef]
- Anjum, N.A.; Gil, R.; Kaushik, M.; Hasanuzzaman, M.; Pereira, E.; Ahmad, I.; Tuteja, N.; Gill, S.S. ATP-sulfurylase, sulfur-compounds, and plant stress tolerance. Front. Plant Sci. 2015, 6, 210. [Google Scholar] [CrossRef]
- Hussain, S.J.; Khan, N.A.; Anjum, N.A.; Masood, A.; Khan, M.I.R. Mechanistic elucidation of salicylic acid and sulphur-induced defence systems, nitrogen metabolism, photosynthetic, and growth potential of mungbean (Vigna radiata) under salt stress. J. Plant Growth Regul. 2020, 40, 1000–1016. [Google Scholar] [CrossRef]
- Jahan, B.; AlAjmi, M.F.; Rehman, M.T.; Khan, N.A. Treatment of nitric oxide supplemented with nitrogen and sulfur regulates photosynthetic performance and stomatal behavior in mustard under salt stress. Physiol. Plant. 2020, 168, 490–510. [Google Scholar] [CrossRef]
- Singhal, R.K.; Fahad, S.; Kumar, P.; Choyal, P.; Javed, T.; Jinger, D.; Singh, P.; Saha, D.; Prathhibha, M.D.; Bose, B.; et al. Beneficial elements: New Players in improving nutrient use efficiency and abiotic stress tolerance. Plant Growth Reg. 2023, 100, 237–265. [Google Scholar] [CrossRef]
- Hasanuzzaman, M.; Bhuyan, M.H.M.B.; Mahmud, J.A.; Nahar, K.; Moh-sin, S.M.; Parvin, K.; Fujita, M. Interaction of sulfur with phytohormones and signaling molecules in conferring abiotic stress tolerance to plants. Plant Signal Behav. 2018, 13, e1477905. [Google Scholar] [CrossRef]
- Rajab, H.; Khan, M.S.; Wirtz, M.; Malagoli, M.; Qahar, F.; Hell, R. Sulfur metabolic engineering enhances cadmium stress tolerance and root to shot iron translocation in Brassica napus L. Plant Physiol. Biochem. 2020, 152, 32–43. [Google Scholar] [CrossRef]
- Prasad, R.; Shivay, Y.S. Sulphur in Soil, Plant and Human Nutrition. Proc. Natl. Acad. Sci. India Sect. B Biol. Sci. 2016, 88, 429–434. [Google Scholar] [CrossRef]
- Zenda, T.; Liu, S.; Dong, A.; Duan, H. Revisting Sulphur—The Once Neglected Nutrient: It’s Roles in Plant Growth, Metabolism, Stress Tolerance and Crop Production. Agriculture 2021, 11, 626. [Google Scholar] [CrossRef]
- Usharani, K.V.; Naik, D.; Manjunatha, R.L. Sulphur Oxidizing Bacteria: Oxidation, Mechanisms and Uses—A Review. Trends Biosci. 2019, 12, 203–211. [Google Scholar]
- Nyamath, S.; Subburamu, K.; Kalyanasundram, G.T.; Balachandar, D.; Suresh, M.; Anandham, R. Multifarious charcteristics of sulfur-ozidizing bacteria residing in rice rizosphere. Folia Microbiol. 2023. [Google Scholar] [CrossRef]
- Grayston, S.J.; James, J.G. Sulfur-oxidizing bacteria as plant growth promoting rhizobacteria for canola. Can. J. Microbiol. 1991, 37, 521–529. [Google Scholar] [CrossRef]
- Nguyen, P.M.; Do, P.T.; Pham, Y.B.; Doan, T.O.; Nguyen, X.C.; Kul, L.W.; Nguyen, D.D.; Vadiveloo, A.; Um, M.J.; Ngo, H.H. Roles, mechanism of action, and potential applications of sulfur-oxidizing bacteria for environmental bioremediation. Sci. Total Environ. 2022, 852, 158203. [Google Scholar] [CrossRef]
- Boroujeni, S.R.; Kalbasi, M.; Asgharzadeh, A.; Baharlouei, J. Evaluating the Potential of Halothiobacillus Bacteria for Sulfur Oxidation and Biomass Production under Saline Soil. Geomicrobiol. J. 2021, 38, 57–65. [Google Scholar] [CrossRef]
- Carreiras, J.; Caçador, I.; Duarte, B. Bioaugmentation Improves Phytoprotection in Halimione portulacoides Exposed to Mild Salt Stress: Perspectives for Salinity Tolerance Improvement. Plants 2022, 11, 1055. [Google Scholar] [CrossRef]
- Hidri, R.; Mahmoud, O.M.; Zorrig, W.; Mahmoudi, H.; Smaoui, A.; Abdelly, C.; Azcon, R.; Debez, A. Plant Growth-Promoting Rhizobacteria Alleviate High Salinity Impact on the Halophyte Suaeda fruticosa by Modulating Antioxidant Defense and Soil Biological Activity. Front. Plant Sci. 2022, 13, 821475. [Google Scholar] [CrossRef]
- Kumar, V.; Raghuvanshi, N.; Pandey, A.K.; Kumar, A.; Thoday-Kennedy, E.; Kant, S. Role of Halotolerant Plant Growth-Promoting Rhizobacteria in Mitigating Salinity Stress: Recent Advances and Possibilities. Agriculture 2023, 13, 168. [Google Scholar] [CrossRef]
- Chaudhary, S.; Dhanker, R.; Singh, K.; Brar, B.; Goyal, S. Characterization of sulfur-oxidizing bacteria isolated from mustard (Brassica juncea L.) rhizosphere having the capability of improving sulfur and nitrogen uptake. J. Appl. Microbiol. 2022, 133, 2814–2825. [Google Scholar] [CrossRef]
- Guo, L.; Zhang, X.; Zhao, J.; Zhang, A.; Pang, Q. Enhancement of sulfur metabolism and antioxidant machinery confers Bacillus sp. Jrh14-10-induced alkaline stress tolerance in plant. Plant Physiol. Biochem. 2023, 203, 108063. [Google Scholar] [CrossRef]
- Praveen, R.; Ashwin, R.; Davis, J.B.; Ambika, H.S. Sulfur oxidizing bacteria in agro ecosystem and its role in plant productivity—A review. J. Appl. Microbiol. 2023, 134, lxad161. [Google Scholar] [CrossRef]
- Ladeiro, B. Saline agriculture in the 21st century: Using salt contaminated re- sources to cope food requirements. J. Bot. 2012, 2012, 310705. [Google Scholar] [CrossRef]
- Ventura, Y.; Myrzabayeva, M.; Alikulov, Z.; Cohen, S.; Shemer, Z.; Sagi, M. The importance of iron supply during repetitive harvesting of Aster tripolium. Funct. Plant Biol. 2013, 40, 968–976. [Google Scholar] [CrossRef]
- Duarte, B.; Goessling, J.W.; Marques, J.C.; Caçador, I. Ecophysiological constraints of Aster tripolium under ex- treme thermal events impacts: Merging biophysical, bio- chemical and genetic insights. Plant Physiol. Biochem. 2015, 97, 217–228. [Google Scholar] [CrossRef] [PubMed]
- Otte, M.L.; Dekkers, I.M.J.; Rozema, J.; Broekman, R.A. Uptake of arsenic by Aster tripolium in relation to rhizosphere oxidation. Can. J. Bot. 1991, 69, 2670–2677. [Google Scholar] [CrossRef]
- Wiszniewska, A.; Koźmińska, A.; Hanus-Fajerska, E.; Dziurka, K.; Dziurka, M. Insight into phytohormonal modulation of defense mechanisms to salt excess in a halophyte and a glycophyte from Asteraceae family. Plant Soil. 2021, 463, 55–76. [Google Scholar] [CrossRef]
- Patel, A.M.; Vaghani, J.J.; Parmar, K.B. Effect of Sulphur Sources and Sulphur Oxidizing Bacteria (SOB) on Yield and Quality of Summer Soybean in Junagadh, Gujarat, India. Int. J. Plant Soil. Sci. 2023, 35, 18–23. [Google Scholar] [CrossRef]
- Kumar, S. Abiotic Stresses and Their Effects on Plant Growth, Yield and Nutritional Quality of Agricultural Produce. Int. J. Food Sci. Agric. 2020, 4, 367–378. [Google Scholar] [CrossRef]
- Kumari, V.V.; Banerjee, P.; Verma, V.C.; Sukumaran, S.; Chandran, M.A.S.; Gopinath, K.A.; Nenkatesh, G.; Yadav, S.K.; Singh, V.K.; Awasthi, N.K. Plant Nutrition: An Effective Way to Alleviate Abiotic Stress in Agricultural Crops. Int. J. Mol. Sci. 2022, 23, 8519. [Google Scholar] [CrossRef]
- Shrama, A.; Kumar, V.; Shahzad, B.; Ramakrishnan, M.; Sidhu, G.P.S.; Bali, A.S.; Handa, N.; Kapoor, D.; Yadav, P.; Khanna, K.; et al. Photosynthetic Response of Plants Under Different Abiotic Stresses: A Review. J. Plant Growth Regul. 2020, 39, 509–531. [Google Scholar] [CrossRef]
- Masood, A.; Iqbal, N.; Khan, N.A. Role of ethylene in alleviation of cadmium-induced photosynthetic capacity inhibition by sulphur in mustard. Plant Cell Environ. 2012, 35, 524–533. [Google Scholar] [CrossRef]
- Asgher, M.; Khan, N.A.; Khan, M.I.R.; Fatma, M.; Masood, A. Ethylene production is associated with alleviation of cadmium-induced oxidative stress by sulfur in mustard types differing in ethylene sensitivity. Ecotoxicol. Environ. Saf. 2014, 106, 54–61. [Google Scholar] [CrossRef]
- Samanta, S.; Singh, A.; Roychoudhury, A. Involvement of sulfur in the regulation of abiotic stress tolerance in plants. In Protective Chemical Agents in the Amelioration of Plant Abiotic Stress: Biochemical and Molecular Perspectives; Roychoudhury, A., Tripathi, D.K., Eds.; Wiley: Hoboken, NJ, USA, 2020; pp. 437–466. [Google Scholar]
- Narayan, O.P.; Kumar, P.; Yadav, B.; Dua, M.; Johri, A.K. Sulfur nutrition and its role in plant growth and development. Plant Signal. Behav. 2023, 18, e2030082-2. [Google Scholar] [CrossRef]
- Joshi, N.; Gothalwal, R.; Singh, M.; Dave, K. Novel sulphur-oxidizing bacteria consummate sulphur deficiency in oil seed crop. Arch. Microbiol. 2021, 203, 1–6. [Google Scholar] [CrossRef]
- Kelly, D.P.; Wood, A.P. Reclassification of some species of Thiobacillus to the newly designated genera Acidithiobacillus gen. nov., Halothiobacillus gen. nov. and Thermithiobacillus gen. nov. Int. J. Syst. Evol. Microbiol. 2000, 50, 489–500. [Google Scholar] [CrossRef]
- Ali, S.; Charles, T.C.; Glick, B.R. Amelioration of damages causedby high salinity stress by plant growth promoting bacterial endo-phytes. Plant Physiol. Biochem. 2014, 80, 160–167. [Google Scholar] [CrossRef]
- Eom, H.; Ashun, E.; Toor, U.A.; Oh, S.E. A solid-phase direct contact bioassay using sulphur-oxidizing bacteria (SOB) to evaluate toxicity of solil contaminated with heavy metals. Sens. Acut. 2020, 305, 127510. [Google Scholar] [CrossRef]
- Durrenne, B.; Durat, P.; Blondel, A.; Fauconnier, M.-L. How cadmim affects the fitness and the glucosinolate content of oilseed rape plants. Environ. Exp. Bot. 2018, 155, 185–194. [Google Scholar] [CrossRef]
- Erofeeva, E.A. Plant hormensis and Shelford’s tolerance law curve. J. For. Res. 2021, 32, 1789–1802. [Google Scholar] [CrossRef]
- Mostofa, M.G.; Fujita, M.; Tran, L.S. Nitric oxide mediates hydrogen peroxide-and salicylic acid-induced salt tolerance in rice (Oryza sativa L.) seedlings. Plant Growth Regul. 2015, 77, 265–277. [Google Scholar] [CrossRef]
- Jiang, J.L.; Tian, Y.; Li, L.; Yu, M.; Hou, R.P.; Ren, X.M. H2S alleviates salinity stress in cucumber by maintaining the Na+/K+ balance and regulating H2S metabolism and oxidative stress response. Front. Plant Sci. 2019, 10, 678. [Google Scholar] [CrossRef]
- Szymańska, S.; Lis, M.I.; Piernik, A.; Hrynkiewicz, K. Pseudomonas stutzeri and Kushneria marisflavi Alleviate Salinity Stress-Associated Damages in Barley, Lettuce, and Sunflower. Front. Microbiol. 2022, 13, 788893. [Google Scholar] [CrossRef]
- Najafi Zilaie, M.; Mosleh Arani, A.; Etesami, H.; Dinarvand, M. Improved salinity and dust stress tolerance in the desert halophyte Haloxylon aphyllum by halotolerant plant growth-promoting rhizobacteria. Front. Plant Sci. 2022, 13, 948260. [Google Scholar] [CrossRef]
- Egamberdieva, D.; Wirth, S.J.; Alqarawi, A.A.; Abd_Allah, E.F.; Hashem, A. Phytohormones and Beneficial Microbes: Essential Components for Plants to Balance Stress and Fitness. Front. Microbiol. 2017, 8, 2104. [Google Scholar] [CrossRef] [PubMed]
- Abbas, R.; Rasul, S.; Aslam, K.; Baber, M.; Shahid, M.; Mubeen, F.; Naqqash, T. Halotolerant PGPR: A Hope for Cultivation of Saline Soils. J. King Saud. Univ. Sci. 2019, 31, 1195–1201. [Google Scholar] [CrossRef]
- Sharma, K.; Sharma, S.; Vaishnav, A.; Jain, R.; Singh, D.; Singh, H.B.; Goel, A.; Singh, S. Salt-Tolerant PGPR Strain Priestia endophytica SK1 Promotes Fenugreek Growth under Salt Stress by Inducing Nitrogen Assimilation and Secondary Metabolites. J. Appl. Microbiol. 2022, 133, 2802–2813. [Google Scholar] [CrossRef] [PubMed]
- Assaha, D.V.M.; Ueda, A.; Saneoka, H.; Al-Yahyai, R.; Yaish, M.W. The Role of Na+ and K+ Transporters in Salt Stress Adaptationin Glycophytes. Front. Physiol. 2017, 8, 509. [Google Scholar] [CrossRef]
- Pan, J.; Peng, F.; Xue, X.; You, Q.; Zhang, W.; Wang, T.; Huang, C. The Growth Promotion of Two Salt-Tolerant Plant Groups withPGPR Inoculation: A Meta-Analysis. Sustainability 2019, 11, 378. [Google Scholar] [CrossRef]
- Hoagland, D.R.; Arnon, D.I. The Water Culture Method for Growing Plants without Soil; California Agricultural Experimental Station Circular No. 347; University of California: Berkeley, CA, USA, 1950; pp. 1–32. [Google Scholar]
- Lichtenthaler, H.K.; Wellburn, A.R. Determinations of total carotenoids and chlorophylls a and b of leaf extracts in different solvents. Biochem. Soc. Trans. 1983, 11, 591–592. [Google Scholar] [CrossRef]
- Bates, L.S.; Waldren, R.P.; Teare, I.D. Rapid determination of free proline for water stress studies. Plant Soil. 1973, 39, 205–207. [Google Scholar] [CrossRef]
- Ainsworth, E.A.l.; Gillespie, K.M. Estimation of total phenolic content and other oxidation substrates in plant tissues using Folin-Ciocalteu reagent. Nat. Protoc. 2007, 2, 875–877. [Google Scholar] [CrossRef]
- Fukumoto, L.R.; Mazza, G. Assessing antioxidant and prooxidant activities of phenolic compounds. J. Agric. Food Chem. 2000, 48, 3597–3604. [Google Scholar] [CrossRef]
- Pekkarinen, S.S.; Stöckmann, H.; Schwarz, K.; Heinonen, I.M.; Hopia, A.I. Antioxidant activity and partitioning of phenolic acids in bulk and emulsified methyl linoleate. J. Agric. Food Chem. 1999, 47, 3036–3043. [Google Scholar] [CrossRef] [PubMed]
- Del Rio, D.; Stewart, A.J.; Pellegrini, N. A review of recent studies on malondialdehyde as toxic molecule and biological marker of oxidative stress. Nutr. Metab. Cardiovasc. Dis. 2005, 15, 316–328. [Google Scholar] [CrossRef] [PubMed]
- Hodges, D.M.; Delong, J.M.; Forney, C.F.; Prange, R.K. Improving the thiobarbituric acid-reactive-substances assay for estimating lipid peroxidation in plant tissues containing anthocyanin and other interfering compounds. Planta 1999, 207, 604–611. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| DW% | GTI% | Stem Length [cm] | |||||
|---|---|---|---|---|---|---|---|
| Non-SOB | SOB | Non-SOB | SOB | Non-SOB | SOB | ||
| leaves | control | 9 a | 10 B | 100 c | 100 B | 60 c | 49 D |
| NaCl | 27 b | 19 D * | 122 c | 121 B | 11 a | 14 B | |
| CdCl2 | 8 a | 7 A | 32 a | 90 AB * | 22 b | 41 C * | |
| NaCl + CdCl2 | 48 c | 14 C * | 54 b | 78 A | 25 b | 8 A | |
| roots | control | 12 c | 7 B * | 100 d | 100 C | ||
| NaCl | 6 b | 5 AB | 49 c | 58 B * | |||
| CdCl2 | 9 b | 20 C * | 23 b | 75 B * | |||
| NaCl + CdCl2 | 3 a | 4 A | 10 a | 13 A | |||
| Total Phenolics | Phenylpropanoids | Flavonols | Anthocyanins | |||||
|---|---|---|---|---|---|---|---|---|
| Non-SOB | SOB | Non-SOB | SOB | Non-SOB | SOB | Non-SOB | SOB | |
| control | 18.5 a | 55.5 B * | 15.9 a | 19.8 B | 5.3 a | 9.7 B * | 0.7 a | 0.7 A |
| NaCl | 54.2 b | 50.5 B | 19.6 b | 23.3 B | 10.7 b | 11.8 C | 0.9 a | 1.4 B * |
| CdCl2 | 8.0 a | 23.9 A * | 4.5 a | 10.7 A * | 3.5 a | 6.6 A * | 0.7 a | 0.7 A |
| NaCl + CdCl2 | 56.7 b | 50.8 B | 22.0 c | 28.8 C * | 30.6 c | 20.3 D * | 2.5 b | 1.3 B * |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Koźmińska, A.; Kamińska, I.; Hanus-Fajerska, E. Sulfur-Oxidizing Bacteria Alleviate Salt and Cadmium Stress in Halophyte Tripolium pannonicum (Jacq.) Dobrocz. Int. J. Mol. Sci. 2024, 25, 2455. https://doi.org/10.3390/ijms25052455
Koźmińska A, Kamińska I, Hanus-Fajerska E. Sulfur-Oxidizing Bacteria Alleviate Salt and Cadmium Stress in Halophyte Tripolium pannonicum (Jacq.) Dobrocz. International Journal of Molecular Sciences. 2024; 25(5):2455. https://doi.org/10.3390/ijms25052455
Chicago/Turabian StyleKoźmińska, Aleksandra, Iwona Kamińska, and Ewa Hanus-Fajerska. 2024. "Sulfur-Oxidizing Bacteria Alleviate Salt and Cadmium Stress in Halophyte Tripolium pannonicum (Jacq.) Dobrocz." International Journal of Molecular Sciences 25, no. 5: 2455. https://doi.org/10.3390/ijms25052455