
Transcriptome Profiling, Physiological and Biochemical Analyses Reveal Comprehensive Insights in Cadmium Stress in Brassica carinata L.
 , , ,
, , , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
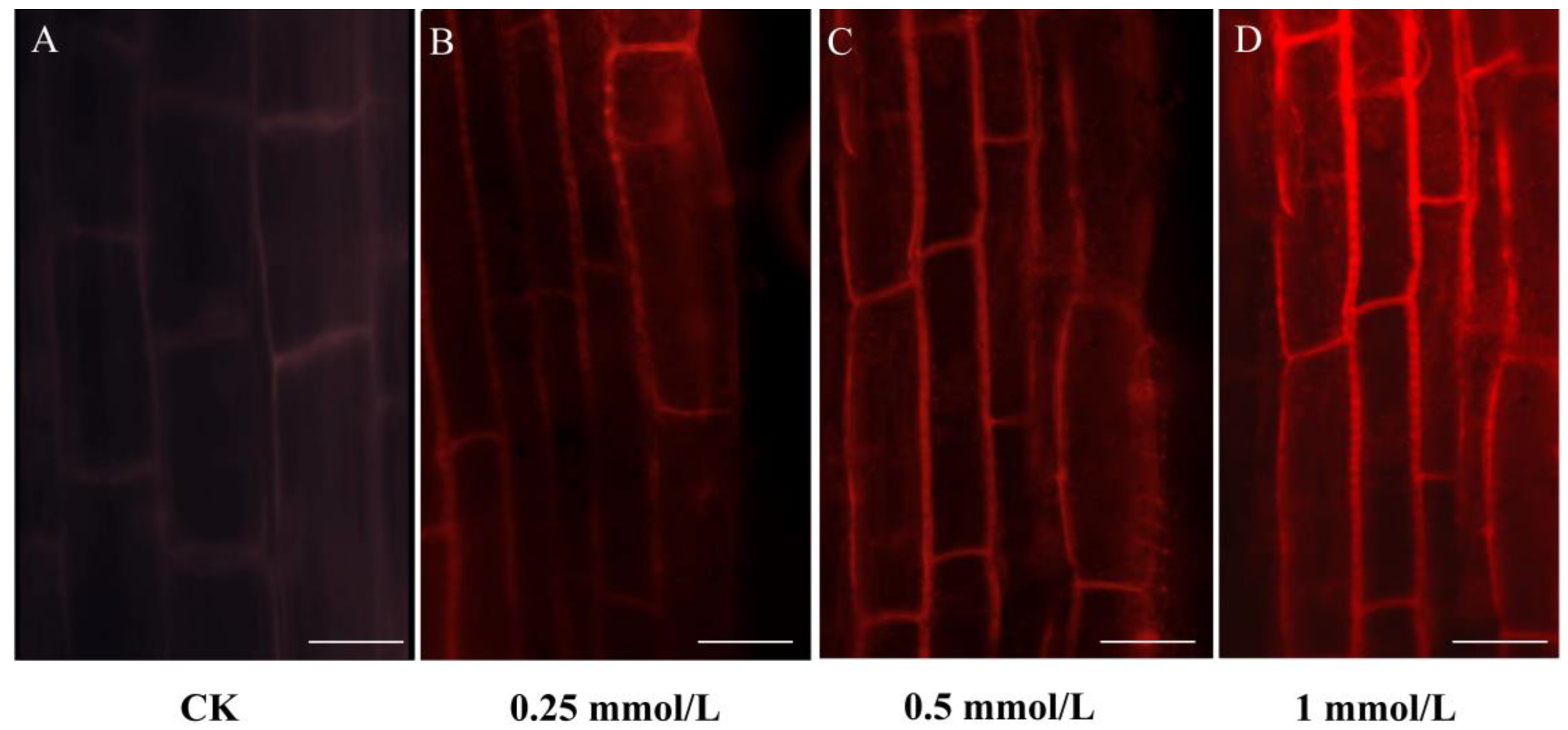
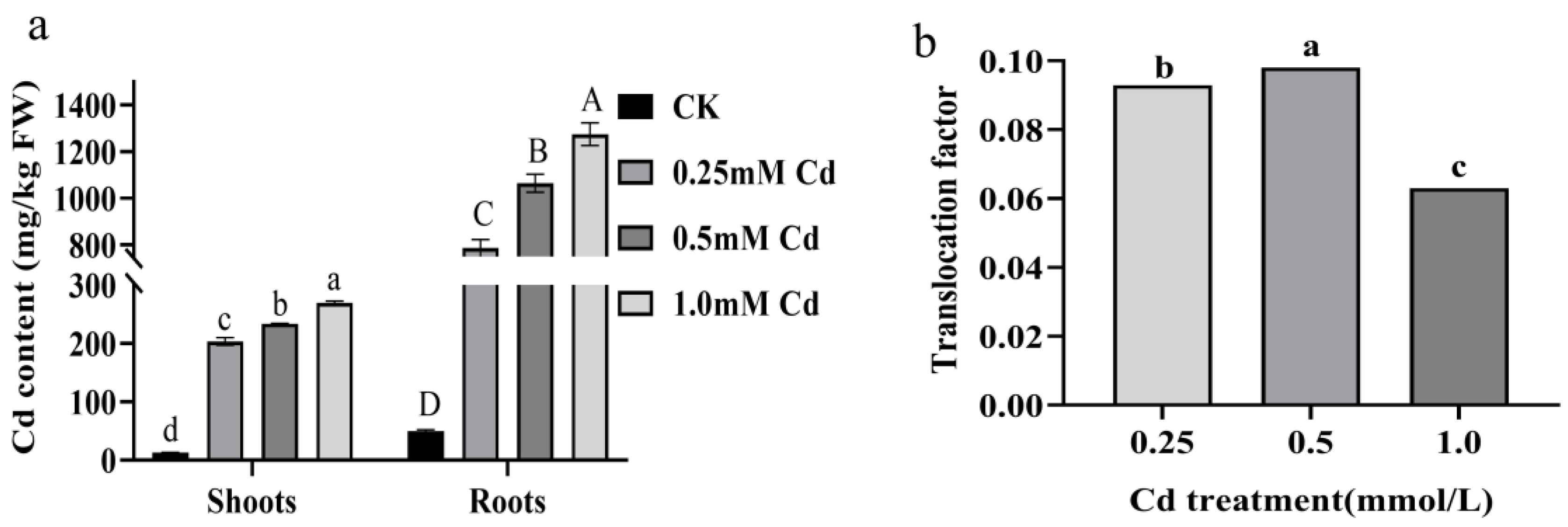
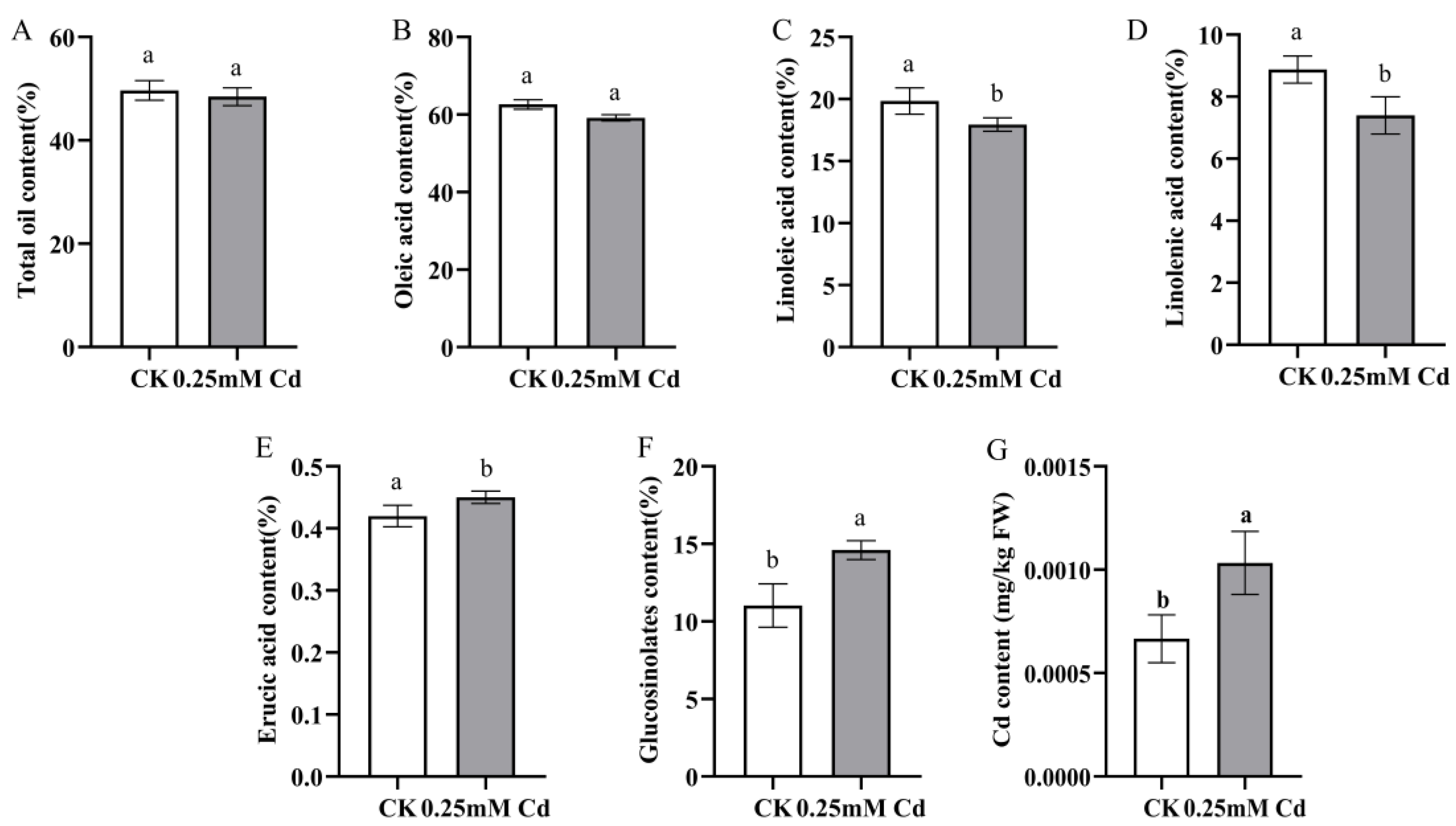
2.1. Cd Tolerance and Accumulation in B. carinata
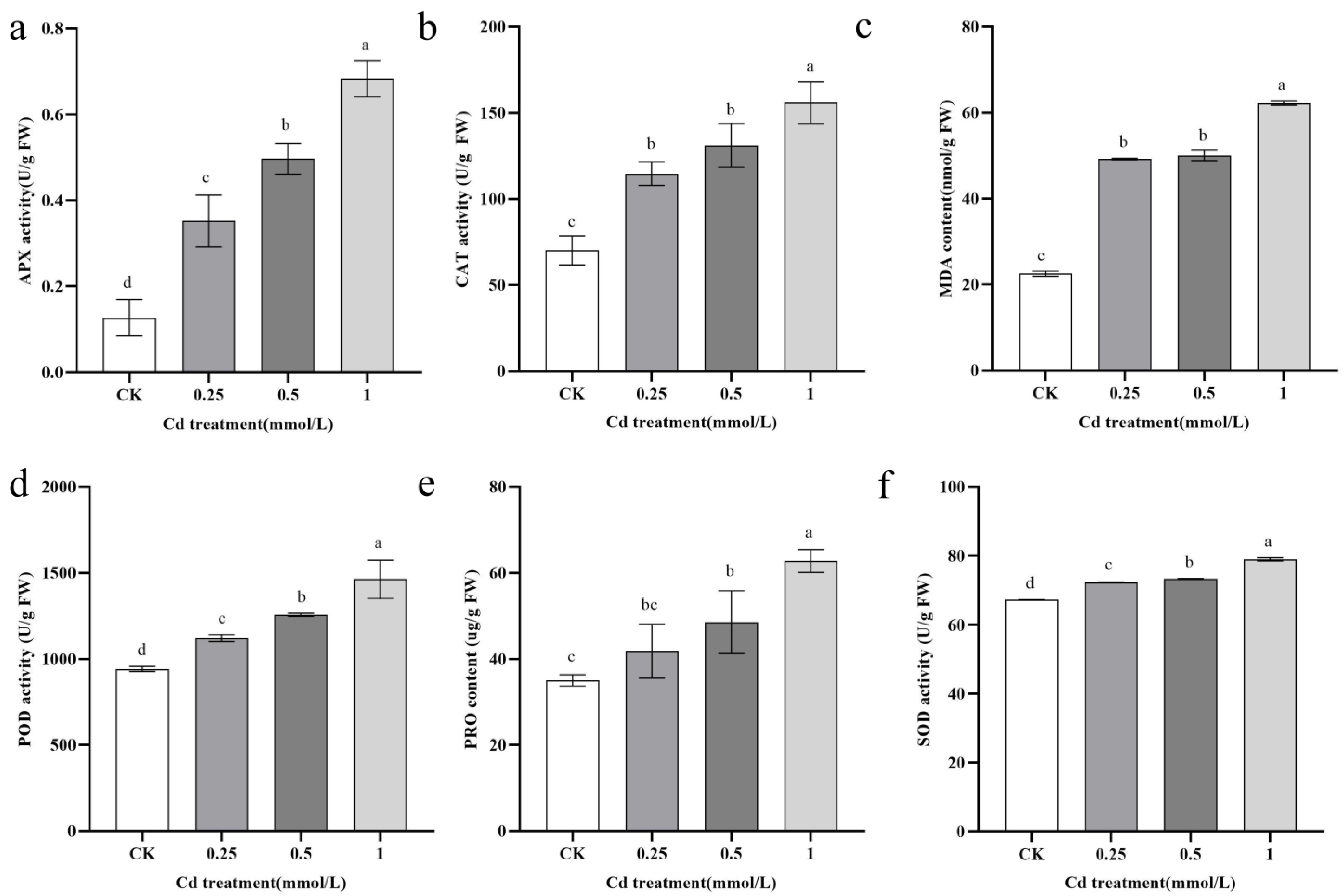
2.2. Physiological Responses of B. carinata to Cd Solutions of Varying Concentrations
2.3. Gene Expression Patterns in B. carinata in Response to Cd Stress
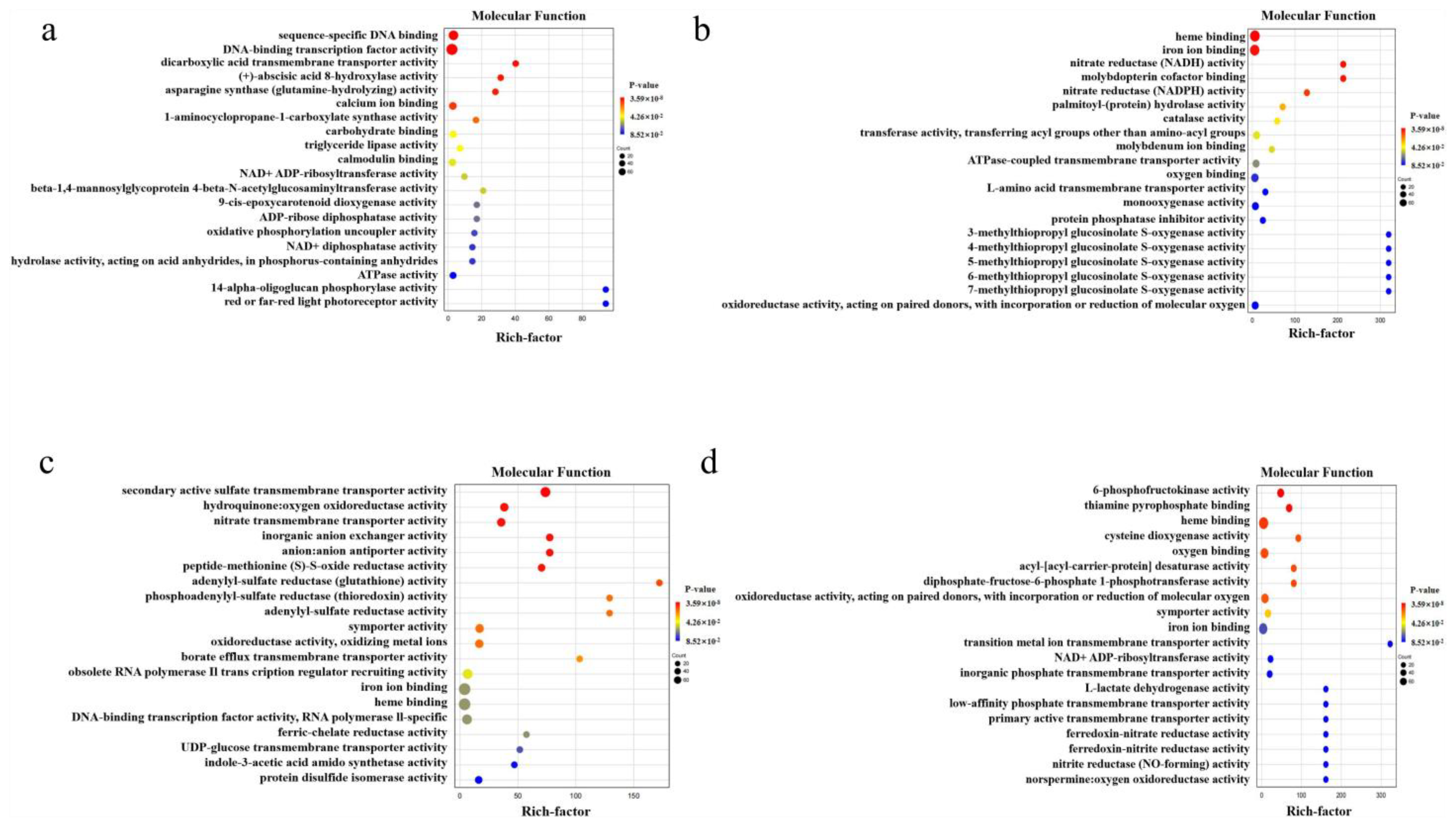
2.4. GO Functions of the DEGs
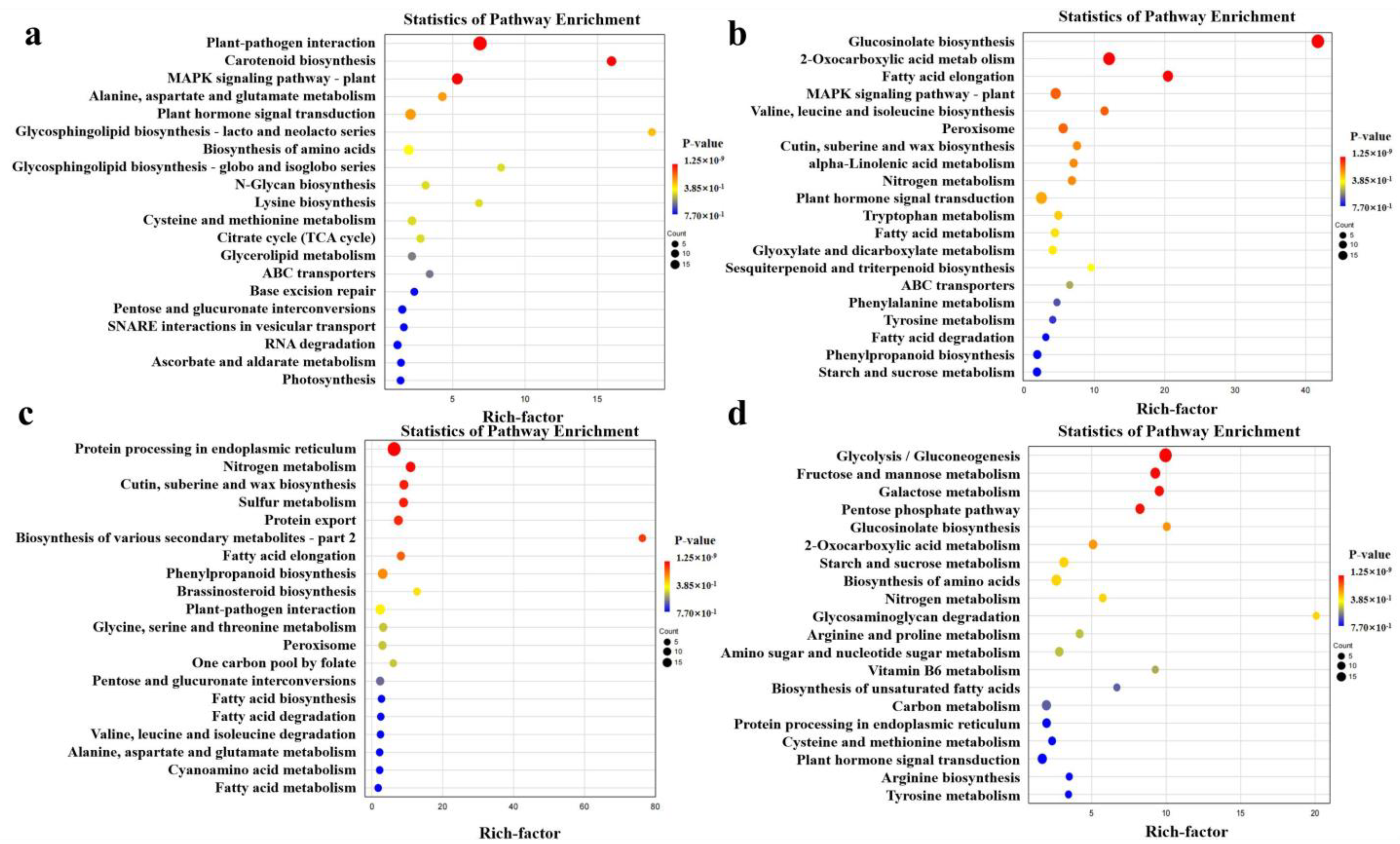
2.5. DEGs Enriched in KEGG Pathways
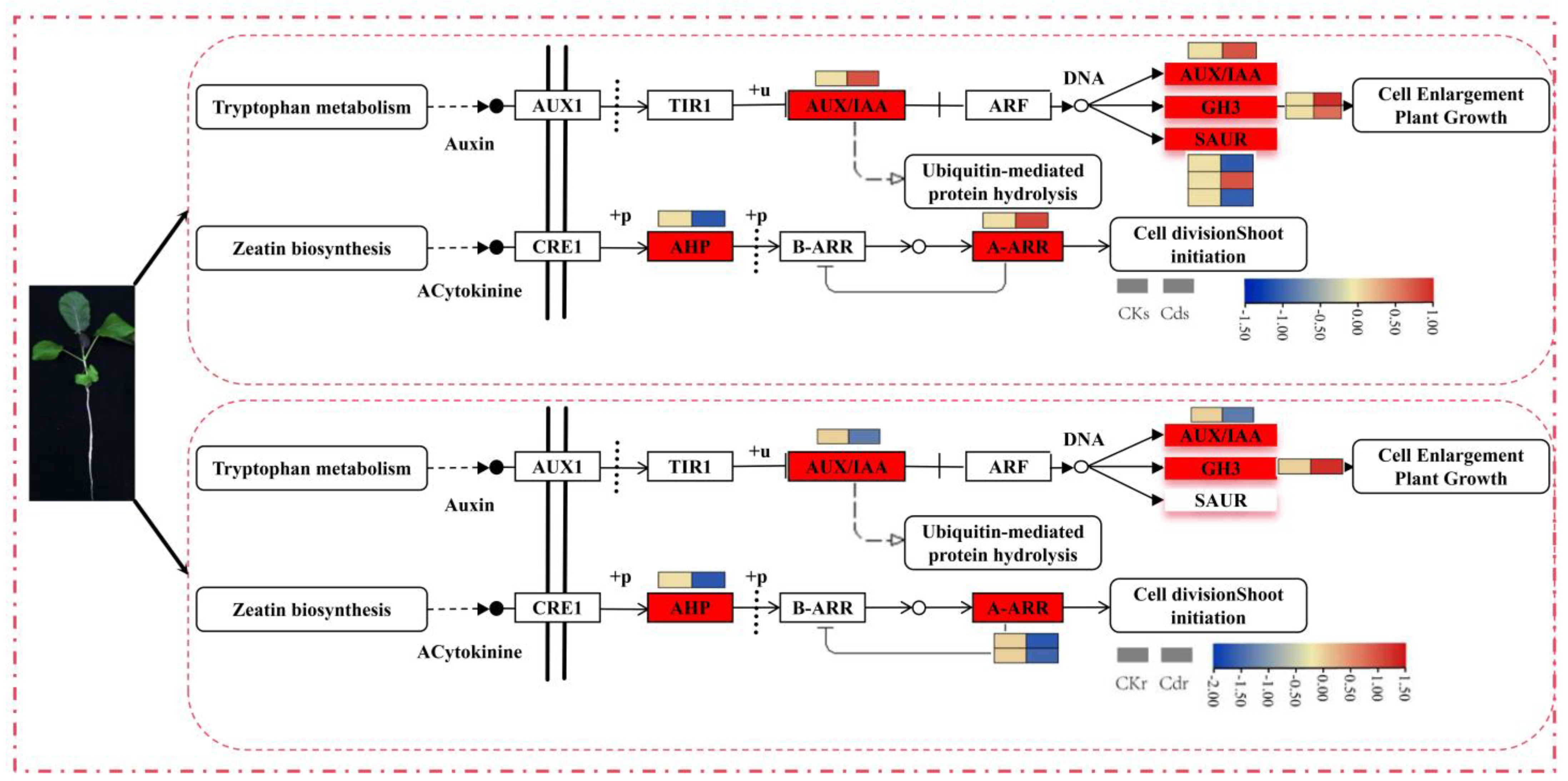
2.6. DEGs Involved in the Plant Hormone Signal Transduction Signalling Pathway
2.7. Responses of Transcription Factors to Cd Stress in B. carinata
3. Discussion
4. Materials and Methods
4.1. Seedling Preparation and Experimental Conditions
4.2. Propidium Iodide Staining
4.3. Determination of Cadmium Content in B. carinata
4.4. Determination of Oil Content and Fatty Acid Content
4.5. Determination of Physiological and Biochemical Indices
4.6. Extracting RNA, Preparing cDNA Libraries, and Identifying Differentially Expressed Genes (DEGs)
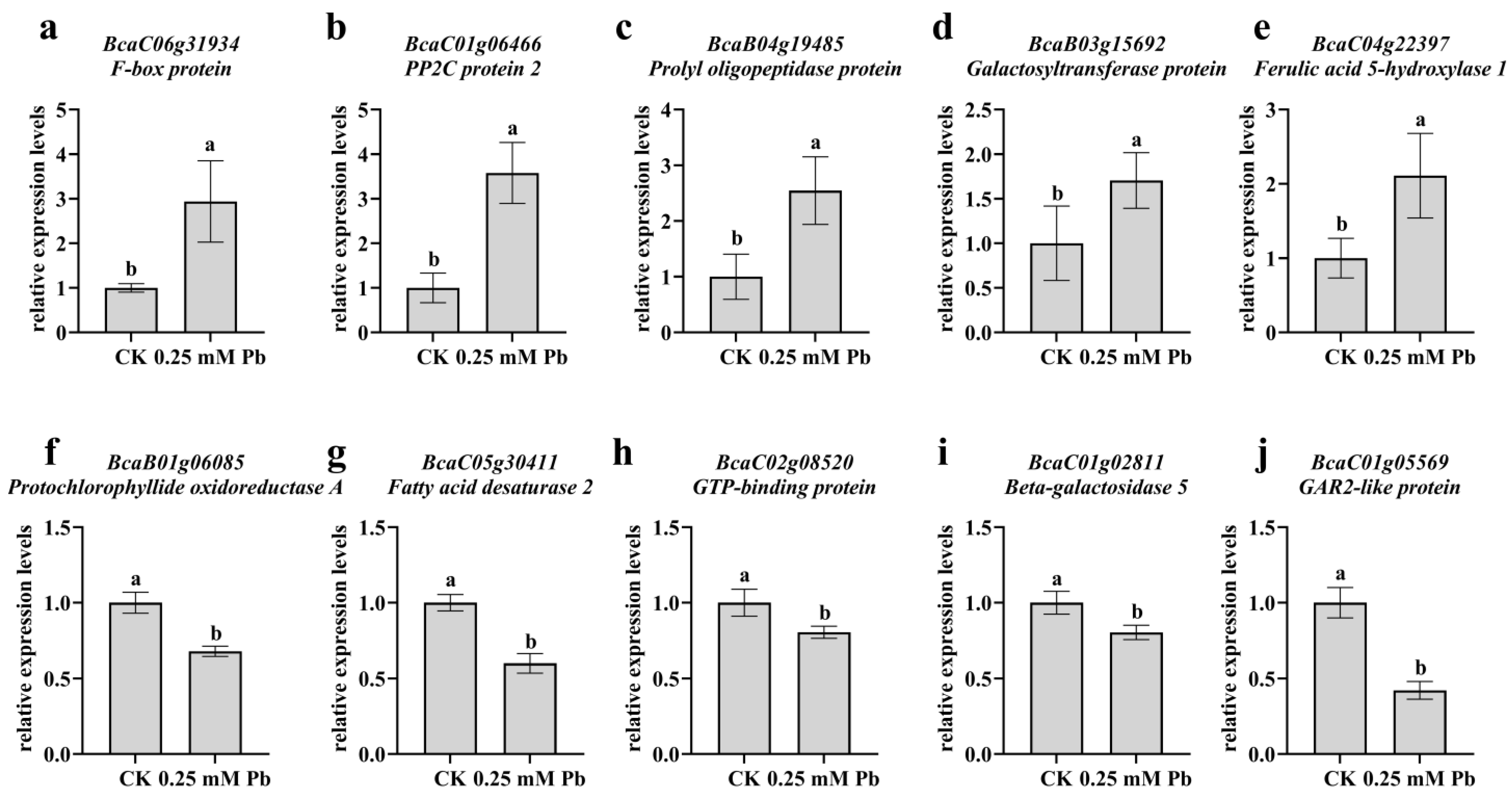
4.7. Validation of the Sequencing Data by qRT–PCR
4.8. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Plekhanova, I.O.; Zolotareva, O.A.; Tarasenko, I.D.; Yakovlev, A.S. Assessment of Ecotoxicity of Soils Contaminated by Heavy Metals. Eurasian Soil Sci. 2019, 52, 1274–1288. [Google Scholar] [CrossRef]
- Kumar, K.; Shinde, A.; Aeron, V.; Verma, A.; Arif, N.S. Genetic Engineering of Plants for Phytoremediation: Advances and Challenges. J. Plant Biochem. Biotechnol. 2023, 32, 12–30. [Google Scholar] [CrossRef]
- Ondrasek, G.; Jelovica Badovinac, I.; Peter, R.; Petravić, M.; Macan, J.; Rengel, Z. Humates and Chlorides Synergistically Increase Cd Phytoaccumulation in Strawberry Fruits, Heightening Health Risk from Cd in Human Diet. Expo. Health 2022, 14, 393–410. [Google Scholar] [CrossRef]
- Huybrechts, M.; Cuypers, A.; Deckers, J.; Iven, V.; Vandionant, S.; Jozefczak, M.; Hendrix, S. Cadmium and Plant Development: An Agony from Seed to Seed. Int. J. Mol. Sci. 2019, 20, 3971. [Google Scholar] [CrossRef]
- Birghila, S.; Matei, N.; Dobrinas, S.; Popescu, V.; Soceanu, A.; Niculescu, A. Assessment of Heavy Metal Content in Soil and Lycopersicon Esculentum (Tomato) and Their Health Implications. Biol. Trace Elem. Res. 2023, 201, 1547–1556. [Google Scholar] [CrossRef]
- Wu, X.; Cobbina, S.J.; Mao, G.; Xu, H.; Zhang, Z.; Yang, L. A Review of Toxicity and Mechanisms of Individual and Mixtures of Heavy Metals in the Environment. Environ. Sci. Pollut. Res. 2016, 23, 8244–8259. [Google Scholar] [CrossRef]
- Goncharuk, E.A.; Zagoskina, N.V. Heavy Metals, Their Phytotoxicity, and the Role of Phenolic Antioxidants in Plant Stress Responses with Focus on Cadmium: Review. Molecules 2023, 28, 3921. [Google Scholar] [CrossRef]
- Gan, C.; Liu, Z.; Pang, B.; Zuo, D.; Hou, Y.; Zhou, L.; Yu, J.; Chen, L.; Wang, H.; Gu, L.; et al. Integrative Physiological and Transcriptome Analyses Provide Insights into the Cadmium (Cd) Tolerance of a Cd Accumulator: Erigeron Canadensis. BMC Genom. 2022, 23, 778. [Google Scholar] [CrossRef]
- Tang, L.; Mao, B.; Li, Y.; Lv, Q.; Zhang, L.; Chen, C.; He, H.; Wang, W.; Zeng, X.; Shao, Y.; et al. Knockout of OsNramp5 Using the CRISPR/Cas9 System Produces Low Cd-Accumulating Indica Rice without Compromising Yield. Sci. Rep. 2017, 7, 14438. [Google Scholar] [CrossRef]
- Wu, D.; Yamaji, N.; Yamane, M.; Kashino-Fujii, M.; Sato, K.; Ma, J.F. The HvNramp5 Transporter Mediates Uptake of Cadmium and Manganese, But Not Iron. Plant Physiol. 2016, 172, 1899–1910. [Google Scholar] [CrossRef]
- Li, G.-Z.; Zheng, Y.-X.; Liu, H.-T.; Liu, J.; Kang, G.-Z. WRKY74 Regulates Cadmium Tolerance through Glutathione-Dependent Pathway in Wheat. Environ. Sci. Pollut. Res. 2022, 29, 68191–68201. [Google Scholar] [CrossRef]
- Sheng, Y.; Yan, X.; Huang, Y.; Han, Y.; Zhang, C.; Ren, Y.; Fan, T.; Xiao, F.; Liu, Y.; Cao, S. The WRKY Transcription Factor, WRKY13, Activates PDR8 Expression to Positively Regulate Cadmium Tolerance in Arabidopsis. Plant Cell Environ. 2019, 42, 891–903. [Google Scholar] [CrossRef]
- Jia, Z.; Li, M.; Wang, H.; Zhu, B.; Gu, L.; Du, X.; Ren, M. TaWRKY70 Positively Regulates TaCAT5 Enhanced Cd Tolerance in Transgenic Arabidopsis. Environ. Exp. Bot. 2021, 190, 104591. [Google Scholar] [CrossRef]
- Li, Z.-G.; Xiang, R.-H.; Wang, J.-Q. Hydrogen Sulfide–Phytohormone Interaction in Plants Under Physiological and Stress Conditions. J. Plant Growth Regul. 2021, 40, 2476–2484. [Google Scholar] [CrossRef]
- Sun, F.; Chen, Z.; Zhang, Q.; Wan, Y.; Hu, R.; Shen, S.; Chen, S.; Yin, N.; Tang, Y.; Liang, Y.; et al. Genome-Wide Identification of the TIFY Gene Family in Brassiceae and Its Potential Association with Heavy Metal Stress in Rapeseed. Plants 2022, 11, 667. [Google Scholar] [CrossRef]
- Jalal, A.; da Silva Oliveira, C.E.; Galindo, F.S.; Rosa, P.A.L.; Gato, I.M.B.; de Lima, B.H.; Teixeira Filho, M.C.M. Regulatory Mechanisms of Plant Growth-Promoting Rhizobacteria and Plant Nutrition against Abiotic Stresses in Brassicaceae Family. Life 2023, 13, 211. [Google Scholar] [CrossRef]
- Tan, P.; Zeng, C.; Wan, C.; Liu, Z.; Dong, X.; Peng, J.; Lin, H.; Li, M.; Liu, Z.; Yan, M. Metabolic Profiles of Brassica Juncea Roots in Response to Cadmium Stress. Metabolites 2021, 11, 383. [Google Scholar] [CrossRef]
- Shehzad, J.; Mustafa, G.; Arshad, H.; Ali, A.; Naveed, N.H.; Riaz, Z.; Khan, I. Morpho-Physiological and Biochemical Responses of Brassica Species toward Lead (Pb) Stress. Acta Physiol. Plant. 2023, 45, 8. [Google Scholar] [CrossRef]
- Song, X.; Wei, Y.; Xiao, D.; Gong, K.; Sun, P.; Ren, Y.; Yuan, J.; Wu, T.; Yang, Q.; Li, X.; et al. Brassica carinata Genome Characterization Clarifies U’s Triangle Model of Evolution and Polyploidy in Brassica. Plant Physiol. 2021, 186, 388–406. [Google Scholar] [CrossRef]
- Goswami, B.; Momtaz, N.; Hoque, M.I.; Sarker, R.H. Performance of F-1 Progenies Developed through Crosses between Brassicacarinata A. Braun A(Female) and Brassicarapa L (Male). Genet. Resour. Crop Evol. 2022, 69, 2745–2753. [Google Scholar] [CrossRef]
- Cui, H.; Wang, Z.; Ye, B.; Hu, K.; Xu, P.; Zhou, J.; Ge, L.; Zheng, X.; Zhou, J. Comprehensive Evaluation on Safe Utilization Potential of Ten Oilseed Rape Cultivars in a Cadmium Contaminated Soil. Environ. Technol. Innov. 2023, 32, 103329. [Google Scholar] [CrossRef]
- Skuza, L.; Szucko-Kociuba, I.; Filip, E.; Bozek, I. Natural Molecular Mechanisms of Plant Hyperaccumulation and Hypertolerance towards Heavy Metals. Int. J. Mol. Sci. 2022, 23, 9335. [Google Scholar] [CrossRef] [PubMed]
- Madhav, S.; Mishra, R.; Kumari, A.; Srivastav, A.L.; Ahamad, A.; Singh, P.; Ahmed, S.; Mishra, P.K.; Sillanpaa, M. A Review on Sources Identification of Heavy Metals in Soil and Remediation Measures by Phytoremediation-Induced Methods. Int. J. Environ. Sci. Technol. 2023, 21, 1099–1120. [Google Scholar] [CrossRef]
- Rizwan, M.; Ali, S.; Zia Ur Rehman, M.; Rinklebe, J.; Tsang, D.C.W.; Bashir, A.; Maqbool, A.; Tack, F.M.G.; Ok, Y.S. Cadmium Phytoremediation Potential of Brassica Crop Species: A Review. Sci. Total Environ. 2018, 631–632, 1175–1191. [Google Scholar] [CrossRef] [PubMed]
- Xiao, L.; Guan, D.; Peart, M.R.; Chen, Y.; Li, Q. The Respective Effects of Soil Heavy Metal Fractions by Sequential Extraction Procedure and Soil Properties on the Accumulation of Heavy Metals in Rice Grains and Brassicas. Environ. Sci. Pollut. Res. 2017, 24, 2558–2571. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Gan, C.; Luo, X.; Dong, C.; Zhou, S.; Xiong, Q.; Weng, Q.; Hu, X.; Du, X.; Zhu, B. Complete Chloroplast Genome Features of the Model Heavy Metal Hyperaccumulator Arabis paniculata Franch and Its Phylogenetic Relationships with Other Brassicaceae Species. Physiol. Mol. Biol. Plants 2022, 28, 775–789. [Google Scholar] [CrossRef] [PubMed]
- Zhang, D.; Du, Y.; He, D.; Zhou, D.; Wu, J.; Peng, J.; Liu, L.; Liu, Z.; Yan, M. Use of Comparative Transcriptomics Combined with Physiological Analyses to Identify Key Factors Underlying Cadmium Accumulation in Brassica juncea L. Front. Genet. 2021, 12, 655885. [Google Scholar] [CrossRef]
- Mansoor, S.; Ali, A.; Kour, N.; Bornhorst, J.; AlHarbi, K.; Rinklebe, J.; Abd El Moneim, D.; Ahmad, P.; Chung, Y.S. Heavy Metal Induced Oxidative Stress Mitigation and ROS Scavenging in Plants. Plants 2023, 12, 3003. [Google Scholar] [CrossRef]
- Małecka, A.; Konkolewska, A.; Hanć, A.; Ciszewska, L.; Staszak, A.M.; Jarmuszkiewicz, W.; Ratajczak, E. Activation of Antioxidative and Detoxificative Systems in Brassica juncea L. Plants against the Toxicity of Heavy Metals. Sci. Rep. 2021, 11, 22345. [Google Scholar] [CrossRef]
- Rashid, M.S.; Liu, G.; Yousaf, B.; Hamid, Y.; Rehman, A.; Munir, M.A.M.; Arif, M.; Ahmed, R.; Song, Y. Assessing the Influence of Sewage Sludge and Derived-Biochar in Immobilization and Transformation of Heavy Metals in Polluted Soil: Impact on Intracellular Free Radical Formation in Maize. Environ. Pollut. 2022, 309, 119768. [Google Scholar] [CrossRef]
- Banerjee, R.; Goswami, P.; Pathak, K.; Mukherjee, A. Vetiver Grass: An Environment Clean-up Tool for Heavy Metal Contaminated Iron Ore Mine-Soil. Ecol. Eng. 2016, 90, 25–34. [Google Scholar] [CrossRef]
- Rizvi, A.; Ahmed, B.; Khan, M.S. An Insight into Stress Induced Phytocompounds Produced in Common Crops Related to Metal Survival. Plant Growth Regul. 2023, 99, 449–464. [Google Scholar] [CrossRef]
- Li, C.; Liu, J.; Wei, Z.; Cheng, Y.; Shen, Z.; Xin, Z.; Huang, Y.; Wang, H.; Li, Y.; Mu, Z.; et al. Exogenous Melatonin Enhances the Tolerance of Tiger Nut (Cyperus esculentus L.) via DNA Damage Repair Pathway under Heavy Metal Stress (Cd2+) at the Sprout Stage. Ecotoxicol. Environ. Saf. 2023, 265, 115519. [Google Scholar] [CrossRef] [PubMed]
- Pang, B.; Zuo, D.; Yang, T.; Yu, J.; Zhou, L.; Hou, Y.; Yu, J.; Ye, L.; Gu, L.; Wang, H.; et al. BcaSOD1 Enhances Cadmium Tolerance in Transgenic Arabidopsis by Regulating the Expression of Genes Related to Heavy Metal Detoxification and Arginine Synthesis. Plant Physiol. Biochem. 2024, 206, 108299. [Google Scholar] [CrossRef] [PubMed]
- El-Enany, A.E.; Issa, A.A. Proline Alleviates Heavy Metal Stress inScenedesmus Armatus. Folia Microbiol. 2001, 46, 227–230. [Google Scholar] [CrossRef] [PubMed]
- Hosseinifard, M.; Stefaniak, S.; Ghorbani Javid, M.; Soltani, E.; Wojtyla, Ł.; Garnczarska, M. Contribution of Exogenous Proline to Abiotic Stresses Tolerance in Plants: A Review. Int. J. Mol. Sci. 2022, 23, 5186. [Google Scholar] [CrossRef]
- Alia; Saradhi, P.P. Proline Accumulation Under Heavy Metal Stress. J. Plant Physiol. 1991, 138, 554–558. [Google Scholar] [CrossRef]
- Zhang, T.; Wang, J.; Sun, Y.; Zhang, L.; Zheng, S. Versatile Roles of Melatonin in Growth and Stress Tolerance in Plants. J. Plant Growth Regul. 2022, 41, 507–523. [Google Scholar] [CrossRef]
- Amjadi, Z.; Namdjoyan, S.; Abolhasani Soorki, A. Exogenous Melatonin and Salicylic Acid Alleviates Cadmium Toxicity in Safflower (Carthamus tinctorius L.) Seedlings. Ecotoxicology 2021, 30, 387–401. [Google Scholar] [CrossRef]
- Liu, Z.; Zhou, L.; Gan, C.; Hu, L.; Pang, B.; Zuo, D.; Wang, G.; Wang, H.; Liu, Y. Transcriptomic Analysis Reveals Key Genes and Pathways Corresponding to Cd and Pb in the Hyperaccumulator Arabis paniculata. Ecotoxicol. Environ. Saf. 2023, 254, 114757. [Google Scholar] [CrossRef]
- Wang, K.; He, J.; Gao, Y.; Han, K.; Liu, J.; Wang, Y. Exogenous Melatonin Improved the Growth and Development of Naked Oat Seedlings under Cadmium Stress. Environ. Sci. Pollut. Res. 2022, 29, 88109–88118. [Google Scholar] [CrossRef] [PubMed]
- Danilova, M.N.; Doroshenko, A.S.; Kudryakova, N.V.; Andreeva, A.A.; Kusnetsov, V.V. Plastome Transcription Machinery and Peculiarities of the Expression of Its Genes during Cytokinin-Dependent Deetiolation of Arabidopsis thaliana. Russ. J. Plant Physiol. 2018, 65, 801–812. [Google Scholar] [CrossRef]
- Gujjar, R.S.; Banyen, P.; Chuekong, W.; Worakan, P.; Roytrakul, S.; Supaibulwatana, K. A Synthetic Cytokinin Improves Photosynthesis in Rice under Drought Stress by Modulating the Abundance of Proteins Related to Stomatal Conductance, Chlorophyll Contents, and Rubisco Activity. Plants 2020, 9, 1106. [Google Scholar] [CrossRef] [PubMed]
- Lu, G.; Casaretto, J.A.; Ying, S.; Mahmood, K.; Liu, F.; Bi, Y.-M.; Rothstein, S.J. Overexpression of OsGATA12 Regulates Chlorophyll Content, Delays Plant Senescence and Improves Rice Yield under High Density Planting. Plant Mol. Biol. 2017, 94, 215–227. [Google Scholar] [CrossRef] [PubMed]
- Piotrowska-Niczyporuk, A.; Bajguz, A.; Kotowska, U.; Zambrzycka-Szelewa, E.; Sienkiewicz, A. Auxins and Cytokinins Regulate Phytohormone Homeostasis and Thiol-Mediated Detoxification in the Green Alga Acutodesmus obliquus Exposed to Lead Stress. Sci. Rep. 2020, 10, 10193. [Google Scholar] [CrossRef]
- Guo, X.; Ullah, A.; Siuta, D.; Kukfisz, B.; Iqbal, S. Role of WRKY Transcription Factors in Regulation of Abiotic Stress Responses in Cotton. Life 2022, 12, 1410. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Lai, Z.; Shi, J.; Xiao, Y.; Chen, Z.; Xu, X. Roles of Arabidopsis WRKY18, WRKY40 and WRKY60 Transcription Factors in Plant Responses to Abscisic Acid and Abiotic Stress. BMC Plant Biol. 2010, 10, 281. [Google Scholar] [CrossRef]
- Jue, D.; Sang, X.; Liu, L.; Shu, B.; Wang, Y.; Liu, C.; Xie, J.; Shi, S. Identification of WRKY Gene Family from Dimocarpus longan and Its Expression Analysis during Flower Induction and Abiotic Stress Responses. Int. J. Mol. Sci. 2018, 19, 2169. [Google Scholar] [CrossRef]
- Li, W.; Pang, S.; Lu, Z.; Jin, B. Function and Mechanism of WRKY Transcription Factors in Abiotic Stress Responses of Plants. Plants 2020, 9, 1515. [Google Scholar] [CrossRef]
- Luo, S.; Calderón-Urrea, A.; Yu, J.; Liao, W.; Xie, J.; Lv, J.; Feng, Z.; Tang, Z. The Role of Hydrogen Sulfide in Plant Alleviates Heavy Metal Stress. Plant Soil 2020, 449, 1–10. [Google Scholar] [CrossRef]
- Yan, L.; Liu, Z.-Q.; Xu, Y.-H.; Lu, K.; Wang, X.-F.; Zhang, D.-P. Auto- and Cross-Repression of Three Arabidopsis WRKY Transcription Factors WRKY18, WRKY40, and WRKY60 Negatively Involved in ABA Signaling. J. Plant Growth Regul. 2013, 32, 399–416. [Google Scholar] [CrossRef]
- Dogan, H.; Coteli, E.; Karatas, F. Determination of Glutathione, Selenium, and Malondialdehyde in Different Edible Mushroom Species. Biol. Trace Elem. Res. 2016, 174, 459–463. [Google Scholar] [CrossRef] [PubMed]
- Yim, W.C.; Swain, M.L.; Ma, D.; An, H.; Bird, K.A.; Curdie, D.D.; Wang, S.; Don Ham, H.; Luzuriaga-Neira, A.; Kirkwood, J.S.; et al. The Last Missing Piece of the Triangle of U: The Evolution of the Tetraploid Brassica carinata Genome. bioRxiv 2022. [Google Scholar] [CrossRef]
- Kim, D.; Paggi, J.M.; Park, C.; Bennett, C.; Salzberg, S.L. Graph-Based Genome Alignment and Genotyping with HISAT2 and HISAT-Genotype. Nat. Biotechnol. 2019, 37, 907–915. [Google Scholar] [CrossRef]









Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, T.; Pang, B.; Zhou, L.; Gu, L.; Wang, H.; Du, X.; Wang, H.; Zhu, B. Transcriptome Profiling, Physiological and Biochemical Analyses Reveal Comprehensive Insights in Cadmium Stress in Brassica carinata L. Int. J. Mol. Sci. 2024, 25, 1260. https://doi.org/10.3390/ijms25021260
Yang T, Pang B, Zhou L, Gu L, Wang H, Du X, Wang H, Zhu B. Transcriptome Profiling, Physiological and Biochemical Analyses Reveal Comprehensive Insights in Cadmium Stress in Brassica carinata L. International Journal of Molecular Sciences. 2024; 25(2):1260. https://doi.org/10.3390/ijms25021260
Chicago/Turabian StyleYang, Tinghai, Biao Pang, Lizhou Zhou, Lei Gu, Hongcheng Wang, Xuye Du, Huinan Wang, and Bin Zhu. 2024. "Transcriptome Profiling, Physiological and Biochemical Analyses Reveal Comprehensive Insights in Cadmium Stress in Brassica carinata L." International Journal of Molecular Sciences 25, no. 2: 1260. https://doi.org/10.3390/ijms25021260