
Molecular Mechanisms of Seasonal Gene Expression in Trees

Abstract
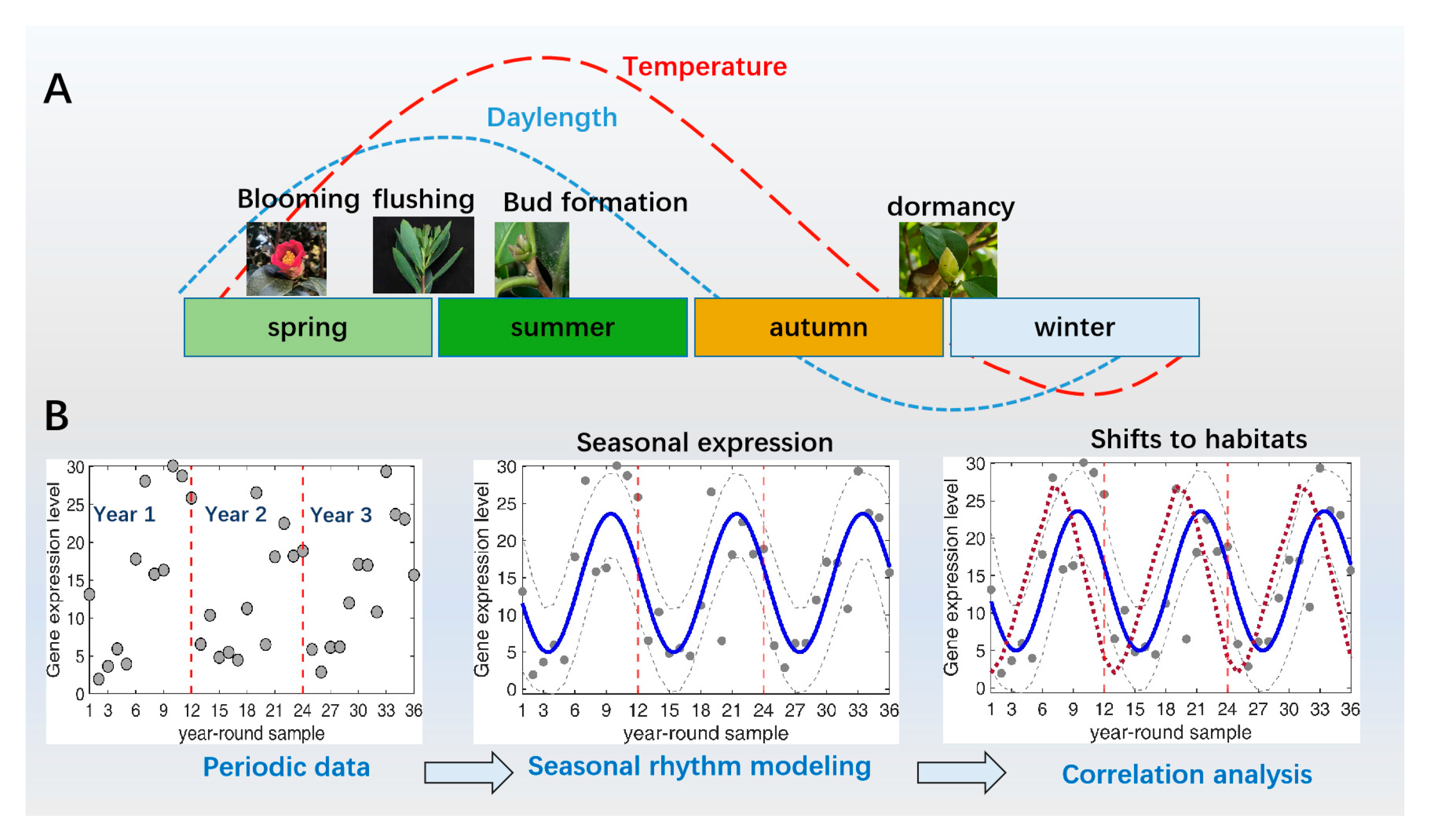
:1. Introduction
2. Seasonal Gene Expression Underlying the Annual Oscillation of Dormancy and Growth
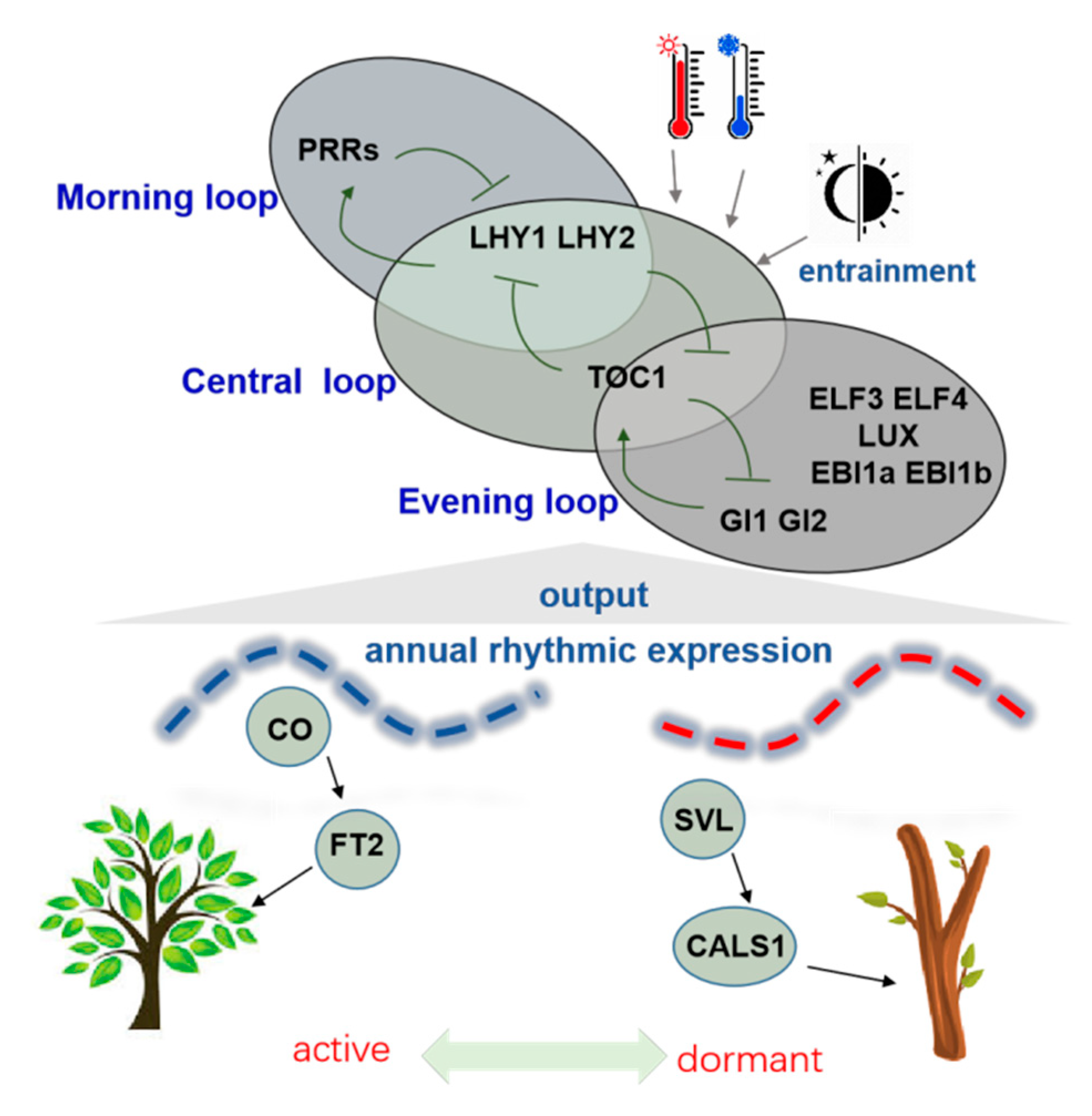
3. Alterations in Circadian Rhythms Are Involved in Regulating Dormancy and Activity of Seasonal Buds
4. Annual Temperature Variations Determine the Seasonal Pattern of Tree Growth
5. DAM and SVP-like Genes Are Dormancy Related MADS-Box Genes in Trees
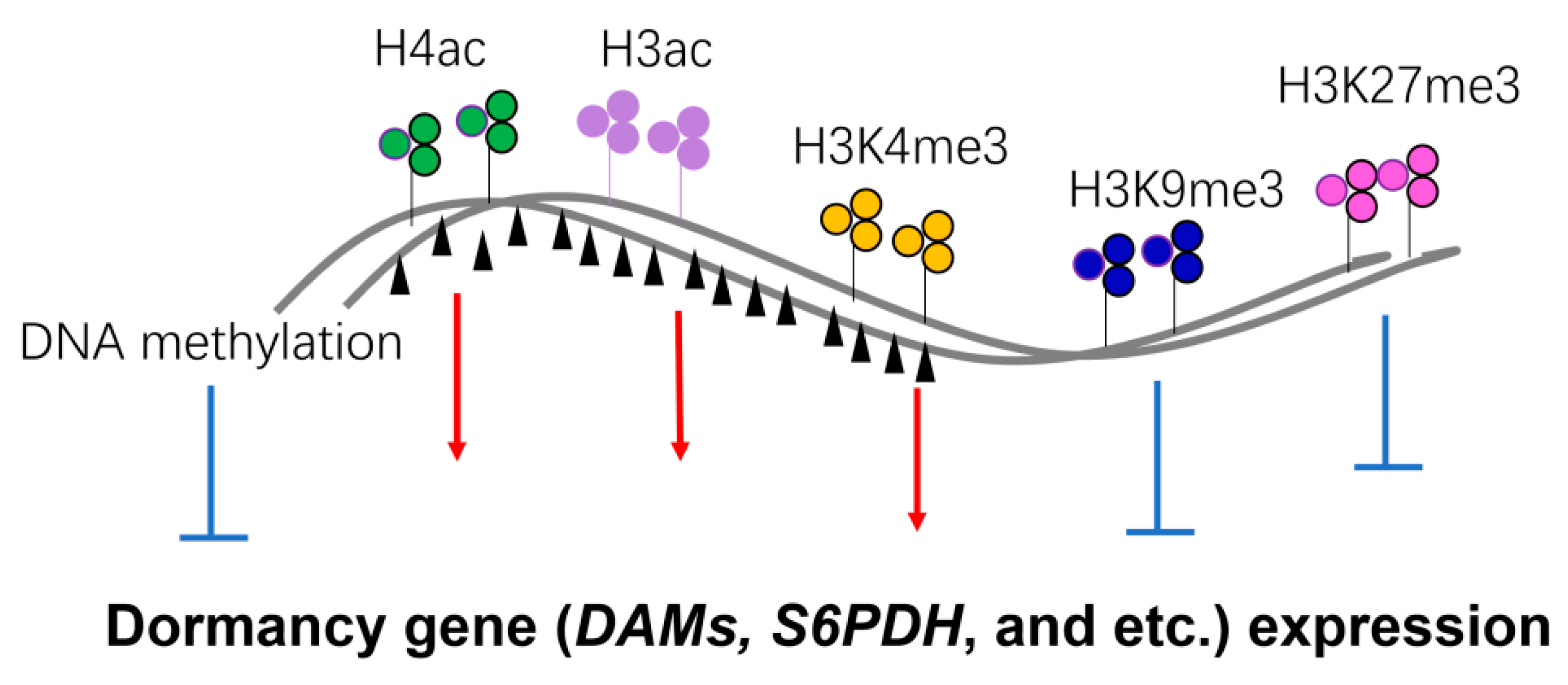
6. Epigenetic Mechanisms of Seasonal Gene Expression
7. Conclusions and Outlook
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Andrés, F.; Coupland, G. The genetic basis of flowering responses to seasonal cues. Nat. Rev. Genet. 2012, 13, 627–639. [Google Scholar] [CrossRef] [PubMed]
- Cooke, J.E.; Eriksson, M.E.; Junttila, O. The dynamic nature of bud dormancy in trees: Environmental control and molecular mechanisms. Plant Cell Environ. 2012, 35, 1707–1728. [Google Scholar] [CrossRef]
- Maurya, J.P.; Bhalerao, R.P. Photoperiod-and temperature-mediated control of growth cessation and dormancy in trees: A molecular perspective. Ann. Bot. 2017, 120, 351–360. [Google Scholar] [CrossRef]
- Horvath, D.P.; Anderson, J.V.; Chao, W.S.; Foley, M.E. Knowing when to grow: Signals regulating bud dormancy. Trends Plant Sci. 2003, 8, 534–540. [Google Scholar] [CrossRef] [PubMed]
- Rohde, A.; Bhalerao, R.P. Plant dormancy in the perennial context. Trends Plant Sci. 2007, 12, 217–223. [Google Scholar] [CrossRef]
- Liu, J.; Sherif, S.M. Hormonal orchestration of bud dormancy cycle in deciduous woody perennials. Front. Plant Sci. 2019, 10, 1136. [Google Scholar] [CrossRef] [PubMed]
- Yang, Q.; Gao, Y.; Wu, X.; Moriguchi, T.; Bai, S.; Teng, Y. Bud endodormancy in deciduous fruit trees: Advances and prospects. Hortic. Res. 2021, 8, 139. [Google Scholar] [CrossRef]
- Körner, C.; Basler, D. Phenology under global warming. Science 2010, 327, 1461–1462. [Google Scholar] [CrossRef]
- Campoy, J.A.; Ruiz, D.; Egea, J. Dormancy in temperate fruit trees in a global warming context: A review. Sci. Hortic. 2011, 130, 357–372. [Google Scholar] [CrossRef]
- Guo, L.; Liu, X.; Alatalo, J.M.; Wang, C.; Xu, J.; Yu, H.; Chen, J.; Yu, Q.; Peng, C.; Dai, J. Climatic drivers and ecological implications of variation in the time interval between leaf-out and flowering. Curr. Biol. 2023, 33, 3338–3349. [Google Scholar] [CrossRef]
- Nagano, A.J.; Kawagoe, T.; Sugisaka, J.; Honjo, M.N.; Iwayama, K.; Kudoh, H. Annual transcriptome dynamics in natural environments reveals plant seasonal adaptation. Nat. Plants 2019, 5, 74–83. [Google Scholar] [CrossRef]
- Hughes, M.E.; Abruzzi, K.C.; Allada, R.; Anafi, R.; Arpat, A.B.; Asher, G.; Baldi, P.; De Bekker, C.; Bell-Pedersen, D.; Blau, J. Guidelines for genome-scale analysis of biological rhythms. J. Biol. Rhythm. 2017, 32, 380–393. [Google Scholar] [CrossRef]
- Deckard, A.; Anafi, R.C.; Hogenesch, J.B.; Haase, S.B.; Harer, J. Design and analysis of large-scale biological rhythm studies: A comparison of algorithms for detecting periodic signals in biological data. Bioinformatics 2013, 29, 3174–3180. [Google Scholar] [CrossRef]
- Cornelissen, G. Cosinor-based rhythmometry. Theor. Biol. Med. Model. 2014, 11, 16. [Google Scholar] [CrossRef] [PubMed]
- Carlucci, M.; Kriščiūnas, A.; Li, H.; Gibas, P.; Koncevičius, K.; Petronis, A.; Oh, G. DiscoRhythm: An Easy-to-Use Web Application and R Package for Discovering Rhythmicity; Oxford University Press: Oxford, UK, 2020; Volume 36, pp. 1952–1954. [Google Scholar]
- Shim, D.; Ko, J.-H.; Kim, W.-C.; Wang, Q.; Keathley, D.E.; Han, K.-H. A molecular framework for seasonal growth-dormancy regulation in perennial plants. Hortic. Res. 2014, 1, 14059. [Google Scholar] [CrossRef] [PubMed]
- Ding, J.; Nilsson, O. Molecular regulation of phenology in trees—Because the seasons they are a-changin’. Curr. Opin. Plant Biol. 2016, 29, 73–79. [Google Scholar] [CrossRef] [PubMed]
- Yanovsky, M.J.; Kay, S.A. Molecular basis of seasonal time measurement in Arabidopsis. Nature 2002, 419, 308–312. [Google Scholar] [CrossRef] [PubMed]
- Devlin, P.F. Signs of the time: Environmental input to the circadian clock. J. Exp. Bot. 2002, 53, 1535–1550. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.A.; Kim, H.-S.; Choi, S.-H.; Jang, J.-Y.; Jeong, M.-J.; Lee, S.I. The importance of the circadian clock in regulating plant metabolism. Int. J. Mol. Sci. 2017, 18, 2680. [Google Scholar] [CrossRef]
- Xu, X.; Yuan, L.; Yang, X.; Zhang, X.; Wang, L.; Xie, Q. Circadian clock in plants: Linking timing to fitness. J. Integr. Plant Biol. 2022, 64, 792–811. [Google Scholar] [CrossRef]
- Srivastava, D.; Shamim, M.; Kumar, M.; Mishra, A.; Maurya, R.; Sharma, D.; Pandey, P.; Singh, K. Role of circadian rhythm in plant system: An update from development to stress response. Environ. Exp. Bot. 2019, 162, 256–271. [Google Scholar] [CrossRef]
- Petterle, A.; Karlberg, A.; Bhalerao, R.P. Daylength mediated control of seasonal growth patterns in perennial trees. Curr. Opin. Plant Biol. 2013, 16, 301–306. [Google Scholar] [CrossRef]
- Ibáñez, C.; Kozarewa, I.; Johansson, M.; Ögren, E.; Rohde, A.; Eriksson, M.E. Circadian clock components regulate entry and affect exit of seasonal dormancy as well as winter hardiness in Populus trees. Plant Physiol. 2010, 153, 1823–1833. [Google Scholar] [CrossRef]
- Singh, R.K.; Bhalerao, R.P.; Eriksson, M.E. Growing in time: Exploring the molecular mechanisms of tree growth. Tree Physiol. 2021, 41, 657–678. [Google Scholar] [CrossRef]
- Takata, N.; Saito, S.; Tanaka Saito, C.; Nanjo, T.; Shinohara, K.; Uemura, M. Molecular phylogeny and expression of poplar circadian clock genes, LHY1 and LHY2. New Phytol. 2009, 181, 808–819. [Google Scholar] [CrossRef] [PubMed]
- Edwards, K.D.; Takata, N.; Johansson, M.; Jurca, M.; Novák, O.; Hényková, E.; Liverani, S.; Kozarewa, I.; Strnad, M.; Millar, A.J. Circadian clock components control daily growth activities by modulating cytokinin levels and cell division-associated gene expression in Populus trees. Plant Cell Environ. 2018, 41, 1468–1482. [Google Scholar] [CrossRef]
- Böhlenius, H.; Huang, T.; Charbonnel-Campaa, L.; Brunner, A.M.; Jansson, S.; Strauss, S.H.; Nilsson, O. CO/FT regulatory module controls timing of flowering and seasonal growth cessation in trees. Science 2006, 312, 1040–1043. [Google Scholar] [CrossRef] [PubMed]
- Koornneef, M.; Hanhart, C.; Van der Veen, J. A genetic and physiological analysis of late flowering mutants in Arabidopsis thaliana. Mol. Gen. Genet. MGG 1991, 229, 57–66. [Google Scholar] [CrossRef] [PubMed]
- André, D.; Marcon, A.; Lee, K.C.; Goretti, D.; Zhang, B.; Delhomme, N.; Schmid, M.; Nilsson, O. FLOWERING LOCUS T paralogs control the annual growth cycle in Populus trees. Curr. Biol. 2022, 32, 2988–2996. [Google Scholar] [CrossRef] [PubMed]
- Ramos-Sanchez, J.M.; Triozzi, P.M.; Alique, D.; Geng, F.; Gao, M.; Jaeger, K.E.; Wigge, P.A.; Allona, I.; Perales, M. LHY2 integrates night-length information to determine timing of poplar photoperiodic growth. Curr. Biol. 2019, 29, 2402–2406. [Google Scholar] [CrossRef] [PubMed]
- Singh, R.K.; Miskolczi, P.; Maurya, J.P.; Bhalerao, R.P. A tree ortholog of SHORT VEGETATIVE PHASE floral repressor mediates photoperiodic control of bud dormancy. Curr. Biol. 2019, 29, 128–133. [Google Scholar] [CrossRef] [PubMed]
- Tylewicz, S.; Petterle, A.; Marttila, S.; Miskolczi, P.; Azeez, A.; Singh, R.K.; Immanen, J.; Mähler, N.; Hvidsten, T.R.; Eklund, D.M. Photoperiodic control of seasonal growth is mediated by ABA acting on cell-cell communication. Science 2018, 360, 212–215. [Google Scholar] [CrossRef] [PubMed]
- Heide, O.; Prestrud, A. Low temperature, but not photoperiod, controls growth cessation and dormancy induction and release in apple and pear. Tree Physiol. 2005, 25, 109–114. [Google Scholar] [CrossRef] [PubMed]
- Rohde, A.; Bastien, C.; Boerjan, W. Temperature signals contribute to the timing of photoperiodic growth cessation and bud set in poplar. Tree Physiol. 2011, 31, 472–482. [Google Scholar] [CrossRef] [PubMed]
- Singh, R.K.; Svystun, T.; AlDahmash, B.; Jönsson, A.M.; Bhalerao, R.P. Photoperiod-and temperature-mediated control of phenology in trees–a molecular perspective. New Phytol. 2017, 213, 511–524. [Google Scholar] [CrossRef]
- Cai, F.; Jin, X.; Tian, Y.; Huang, Z.; Wang, X.; Zhang, Y.; Sun, Y.; Shao, C. Molecular regulation of bud dormancy in perennial plants. Plant Growth Regul. 2024, 102, 1–11. [Google Scholar] [CrossRef]
- Rinne, P.L.; Paul, L.K.; van der Schoot, C. Decoupling photo-and thermoperiod by projected climate change perturbs bud development, dormancy establishment and vernalization in the model tree Populus. BMC Plant Biol. 2018, 18, 220. [Google Scholar] [CrossRef]
- Yang, Y.; Wu, Z.; Guo, L.; He, H.S.; Ling, Y.; Wang, L.; Zong, S.; Na, R.; Du, H.; Li, M.-H. Effects of winter chilling vs. spring forcing on the spring phenology of trees in a cold region and a warmer reference region. Sci. Total Environ. 2020, 725, 138323. [Google Scholar] [CrossRef]
- Zeng, X.; Du, Y.; Vitasse, Y. Untangling winter chilling and spring forcing effects on spring phenology of subtropical tree seedlings. Agric. For. Meteorol. 2023, 335, 109456. [Google Scholar] [CrossRef]
- Camargo, H.; Salazar, M.; Keller, M.; Hoogenboom, G. Modeling the effect of temperature on bud dormancy of grapevines. Agric. For. Meteorol. 2019, 280, 1016. [Google Scholar]
- Lang, G.A.; Early, J.D.; Martin, G.C.; Darnell, R.L. Endo-, para-, and ecodormancy: Physiological terminology and classification for dormancy research. HortScience 1987, 22, 371–377. [Google Scholar] [CrossRef]
- Shi, P.; Chen, Z.; Reddy, G.V.; Hui, C.; Huang, J.; Xiao, M. Timing of cherry tree blooming: Contrasting effects of rising winter low temperatures and early spring temperatures. Agric. For. Meteorol. 2017, 240, 78–89. [Google Scholar] [CrossRef]
- Baumgarten, F.; Zohner, C.M.; Gessler, A.; Vitasse, Y. Chilled to be forced: The best dose to wake up buds from winter dormancy. New Phytol. 2021, 230, 1366–1377. [Google Scholar] [CrossRef] [PubMed]
- Gregis, V.; Sessa, A.; Dorca-Fornell, C.; Kater, M.M. The Arabidopsis floral meristem identity genes AP1, AGL24 and SVP directly repress class B and C floral homeotic genes. Plant J. 2009, 60, 626–637. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.H.; Yoo, S.J.; Park, S.H.; Hwang, I.; Lee, J.S.; Ahn, J.H. Role of SVP in the control of flowering time by ambient temperature in Arabidopsis. Genes Dev. 2007, 21, 397–402. [Google Scholar] [CrossRef]
- Michaels, S.D.; Ditta, G.; Gustafson-Brown, C.; Pelaz, S.; Yanofsky, M.; Amasino, R.M. AGL24 acts as a promoter of flowering in Arabidopsis and is positively regulated by vernalization. Plant J. 2003, 33, 867–874. [Google Scholar] [CrossRef] [PubMed]
- Singh, R.K.; Maurya, J.P.; Azeez, A.; Miskolczi, P.; Tylewicz, S.; Stojkovič, K.; Delhomme, N.; Busov, V.; Bhalerao, R.P. A genetic network mediating the control of bud break in hybrid aspen. Nat. Commun. 2018, 9, 4173. [Google Scholar] [CrossRef] [PubMed]
- Falavigna, V.d.S.; Guitton, B.; Costes, E.; Andrés, F. I want to (bud) break free: The potential role of DAM and SVP-like genes in regulating dormancy cycle in temperate fruit trees. Front. Plant Sci. 2019, 9, 1990. [Google Scholar] [CrossRef]
- Bielenberg, D.G.; Wang, Y.; Li, Z.; Zhebentyayeva, T.; Fan, S.; Reighard, G.L.; Scorza, R.; Abbott, A.G. Sequencing and annotation of the evergrowing locus in peach [Prunus persica (L.) Batsch] reveals a cluster of six MADS-box transcription factors as candidate genes for regulation of terminal bud formation. Tree Genet. Genomes 2008, 4, 495–507. [Google Scholar] [CrossRef]
- Yamane, H. Regulation of bud dormancy and bud break in Japanese apricot (Prunus mume Siebold & Zucc.) and peach [Prunus persica (L.) Batsch]: A summary of recent studies. J. Jpn. Soc. Hortic. Sci. 2014, 83, 187–202. [Google Scholar]
- Wu, R.; Tomes, S.; Karunairetnam, S.; Tustin, S.D.; Hellens, R.P.; Allan, A.C.; Macknight, R.C.; Varkonyi-Gasic, E. SVP-like MADS box genes control dormancy and budbreak in apple. Front. Plant Sci. 2017, 8, 477. [Google Scholar] [CrossRef]
- Sasaki, R.; Yamane, H.; Ooka, T.; Jotatsu, H.; Kitamura, Y.; Akagi, T.; Tao, R. Functional and expressional analyses of PmDAM genes associated with endodormancy in Japanese apricot. Plant Physiol. 2011, 157, 485–497. [Google Scholar] [CrossRef] [PubMed]
- Wu, R.-M.; Walton, E.F.; Richardson, A.C.; Wood, M.; Hellens, R.P.; Varkonyi-Gasic, E. Conservation and divergence of four kiwifruit SVP-like MADS-box genes suggest distinct roles in kiwifruit bud dormancy and flowering. J. Exp. Bot. 2012, 63, 797–807. [Google Scholar] [CrossRef] [PubMed]
- Mazzitelli, L.; Hancock, R.D.; Haupt, S.; Walker, P.G.; Pont, S.D.; McNicol, J.; Cardle, L.; Morris, J.; Viola, R.; Brennan, R. Co-ordinated gene expression during phases of dormancy release in raspberry (Rubus idaeus L.) buds. J. Exp. Bot. 2007, 58, 1035–1045. [Google Scholar] [CrossRef] [PubMed]
- Yamane, H.; Kashiwa, Y.; Ooka, T.; Tao, R.; Yonemori, K. Suppression subtractive hybridization and differential screening reveals endodormancy-associated expression of an SVP/AGL24-type MADS-box gene in lateral vegetative buds of Japanese apricot. J. Am. Soc. Hortic. Sci. 2008, 133, 708–716. [Google Scholar] [CrossRef]
- Takemura, Y.; Kuroki, K.; Matsumoto, K.; Ban, Y.; Moriguchi, T.; Tamura, F. Identification and expression analysis of candidate genes related to endodormancy induction and breaking in Pyrus pyrifolia. Sci. Hortic. 2013, 155, 65–71. [Google Scholar] [CrossRef]
- Mimida, N.; Saito, T.; Moriguchi, T.; Suzuki, A.; Komori, S.; Wada, M. Expression of DORMANCY-ASSOCIATED MADS-BOX (DAM)-like genes in apple. Biol. Plant. 2015, 59, 237–244. [Google Scholar] [CrossRef]
- Dong, Y.; Khalil-Ur-Rehman, M.; Liu, X.; Wang, X.; Yang, L.; Tao, J.; Zheng, H. Functional characterisation of five SVP genes in grape bud dormancy and flowering. Plant Growth Regul. 2022, 97, 511–522. [Google Scholar] [CrossRef]
- Wu, R.; Wang, T.; Richardson, A.C.; Allan, A.C.; Macknight, R.C.; Varkonyi-Gasic, E. Histone modification and activation by SOC1-like and drought stress-related transcription factors may regulate AcSVP2 expression during kiwifruit winter dormancy. Plant Sci. 2019, 281, 242–250. [Google Scholar] [CrossRef]
- Li, Y.; Zhou, Y.; Yang, W.; Cheng, T.; Wang, J.; Zhang, Q. Isolation and functional characterization of SVP-like genes in Prunus mume. Sci. Hortic. 2017, 215, 91–101. [Google Scholar] [CrossRef]
- Moser, M.; Asquini, E.; Miolli, G.V.; Weigl, K.; Hanke, M.-V.; Flachowsky, H.; Si-Ammour, A. The MADS-box gene MdDAM1 controls growth cessation and bud dormancy in apple. Front. Plant Sci. 2020, 11, 1003. [Google Scholar] [CrossRef]
- Yarur, A.; Soto, E.; León, G.; Almeida, A.M. The sweet cherry (Prunus avium) FLOWERING LOCUS T gene is expressed during floral bud determination and can promote flowering in a winter-annual Arabidopsis accession. Plant Reprod. 2016, 29, 311–322. [Google Scholar] [CrossRef]
- Carmona, M.J.; Calonje, M.; Martínez-Zapater, J.M. The FT/TFL1 gene family in grapevine. Plant Mol. Biol. 2007, 63, 637–650. [Google Scholar] [CrossRef] [PubMed]
- Hsu, C.-Y.; Liu, Y.; Luthe, D.S.; Yuceer, C. Poplar FT2 shortens the juvenile phase and promotes seasonal flowering. Plant Cell 2006, 18, 1846–1861. [Google Scholar] [CrossRef] [PubMed]
- Zuo, X.; Wang, S.; Xiang, W.; Yang, H.; Tahir, M.M.; Zheng, S.; An, N.; Han, M.; Zhao, C.; Zhang, D. Genome-wide identification of the 14–3-3 gene family and its participation in floral transition by interacting with TFL1/FT in apple. BMC Genom. 2021, 22, 41. [Google Scholar] [CrossRef] [PubMed]
- Ito, A.; Saito, T.; Sakamoto, D.; Sugiura, T.; Bai, S.; Moriguchi, T. Physiological differences between bud breaking and flowering after dormancy completion revealed by DAM and FT/TFL1 expression in Japanese pear (Pyrus pyrifolia). Tree Physiol. 2016, 36, 109–120. [Google Scholar] [CrossRef] [PubMed]
- Niu, Q.; Li, J.; Cai, D.; Qian, M.; Jia, H.; Bai, S.; Hussain, S.; Liu, G.; Teng, Y.; Zheng, X. Dormancy-associated MADS-box genes and microRNAs jointly control dormancy transition in pear (Pyrus pyrifolia white pear group) flower bud. J. Exp. Bot. 2016, 67, 239–257. [Google Scholar] [CrossRef]
- Khalil-Ur-Rehman, M.; Dong, Y.; Faheem, M.; Zeng, J.; Wang, W.; Tao, J. Expression profiling of ABA and GA signaling cascades regulating bud dormancy in grape. Sci. Hortic. 2019, 246, 44–50. [Google Scholar] [CrossRef]
- André, D.; Zambrano, J.A.; Zhang, B.; Lee, K.C.; Rühl, M.; Marcon, A.; Nilsson, O. Populus SVL acts in leaves to modulate the timing of growth cessation and bud set. Front. Plant Sci. 2022, 13, 823019. [Google Scholar] [CrossRef]
- Tuan, P.A.; Bai, S.; Saito, T.; Ito, A.; Moriguchi, T. Dormancy-Associated MADS-Box (DAM) and the abscisic acid pathway regulate pear endodormancy through a feedback mechanism. Plant Cell Physiol. 2017, 58, 1378–1390. [Google Scholar] [CrossRef]
- Zhang, Y.-z.; Chen, X.; Lu, W.-l.; Wang, X.-z.; Ning, W.; Meng, X.-g.; Fang, Y.-h.; Tan, Q.-p.; Chen, X.-d.; Fu, X.-l. PpMAPK6 regulates peach bud endodormancy release through interactions with PpDAM6. J. Integr. Agric. 2023, 22, 139–148. [Google Scholar] [CrossRef]
- Yang, Q.; Yang, B.; Li, J.; Wang, Y.; Tao, R.; Yang, F.; Wu, X.; Yan, X.; Ahmad, M.; Shen, J. ABA-responsive ABRE-BINDING FACTOR3 activates DAM3 expression to promote bud dormancy in Asian pear. Plant Cell Environ. 2020, 43, 1360–1375. [Google Scholar] [CrossRef] [PubMed]
- Dong, Y.; Liu, J.; Huang, L.; Yang, L.; Khalil-Ur-Rehman, M.; Zheng, H.; Tao, J. SHORT VEGETATIVE PHASE 3 mediates hormonal signals control of bud dormancy in grape. Environ. Exp. Bot. 2023, 211, 105342. [Google Scholar] [CrossRef]
- Lee, J.; Lee, I. Regulation and function of SOC1, a flowering pathway integrator. J. Exp. Bot. 2010, 61, 2247–2254. [Google Scholar] [CrossRef] [PubMed]
- Yu, H.; Xu, Y.; Tan, E.L.; Kumar, P.P. AGAMOUS-LIKE 24, a dosage-dependent mediator of the flowering signals. Proc. Natl. Acad. Sci. USA 2002, 99, 16336–16341. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Xi, W.; Shen, L.; Tan, C.; Yu, H. Regulation of floral patterning by flowering time genes. Dev. Cell 2009, 16, 711–722. [Google Scholar] [CrossRef]
- Trainin, T.; Bar-Ya’akov, I.; Holland, D. ParSOC1, a MADS-box gene closely related to Arabidopsis AGL20/SOC1, is expressed in apricot leaves in a diurnal manner and is linked with chilling requirements for dormancy break. Tree Genet. Genomes 2013, 9, 753–766. [Google Scholar] [CrossRef]
- Zhang, Y.; Li, Y.; Zhang, Y.; Guan, S.; Liu, C.; Zheng, G.; Gai, S. Isolation and characterization of a SOC1-Like gene from tree peony (Paeonia suffruticosa). Plant Mol. Biol. Report. 2015, 33, 855–866. [Google Scholar] [CrossRef]
- Voogd, C.; Wang, T.; Varkonyi-Gasic, E. Functional and expression analyses of kiwifruit SOC1-like genes suggest that they may not have a role in the transition to flowering but may affect the duration of dormancy. J. Exp. Bot. 2015, 66, 4699–4710. [Google Scholar] [CrossRef]
- Wang, J.; Gao, Z.; Li, H.; Jiu, S.; Qu, Y.; Wang, L.; Ma, C.; Xu, W.; Wang, S.; Zhang, C. Dormancy-associated MADS-Box (DAM) genes influence chilling requirement of sweet cherries and co-regulate flower development with SOC1 gene. Int. J. Mol. Sci. 2020, 21, 921. [Google Scholar] [CrossRef]
- Falavigna, V.; Severing, E.; Estevan, J.; Farrera, I.; Hugouvieux, V.; Revers, L.; Zubieta, C.; Coupland, G.; Costes, E.; Andrés, F. Potential role of apple SOC1-like transcription factors in gene regulatory networks involved in bud dormancy. In I International Symposium on Reproductive Biology of Fruit Tree Species; International Society for Horticultural Science: Montpellier, France, 2021; pp. 41–48. [Google Scholar]
- Ríos, G.; Leida, C.; Conejero, A.; Badenes, M.L. Epigenetic regulation of bud dormancy events in perennial plants. Front. Plant Sci. 2014, 5, 247. [Google Scholar]
- Chen, W.; Tamada, Y.; Yamane, H.; Matsushita, M.; Osako, Y.; Gao-Takai, M.; Luo, Z.; Tao, R. H3K4me3 plays a key role in establishing permissive chromatin states during bud dormancy and bud break in apple. Plant J. 2022, 111, 1015–1031. [Google Scholar] [CrossRef]
- Conde, D.; Perales, M.; Sreedasyam, A.; Tuskan, G.A.; Lloret, A.; Badenes, M.L.; González-Melendi, P.; Ríos, G.; Allona, I. Engineering tree seasonal cycles of growth through chromatin modification. Front. Plant Sci. 2019, 10, 412. [Google Scholar] [CrossRef] [PubMed]
- Leida, C.; Conesa, A.; Llácer, G.; Badenes, M.L.; Ríos, G. Histone modifications and expression of DAM6 gene in peach are modulated during bud dormancy release in a cultivar-dependent manner. New Phytol. 2012, 193, 67–80. [Google Scholar] [CrossRef]
- Zhu, H.; Chen, P.-Y.; Zhong, S.; Dardick, C.; Callahan, A.; An, Y.-Q.; van Knocker, S.; Yang, Y.; Zhong, G.-Y.; Abbott, A. Thermal-responsive genetic and epigenetic regulation of DAM cluster controlling dormancy and chilling requirement in peach floral buds. Hortic. Res. 2020, 7, 114. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Li, Y.; Cao, K.; Fang, W.; Chen, C.; Wang, X.; Wu, J.; Guo, W.; Wang, L. Peculiarity of transcriptional and H3K27me3 dynamics during peach bud dormancy. Hortic. Plant J. 2023, 10, 38–50. [Google Scholar] [CrossRef]
- Lloret, A.; Martínez-Fuentes, A.; Agustí, M.; Badenes, M.L.; Ríos, G. Chromatin-associated regulation of sorbitol synthesis in flower buds of peach. Plant Mol. Biol. 2017, 95, 507–517. [Google Scholar] [CrossRef]
- Anh Tuan, P.; Bai, S.; Saito, T.; Imai, T.; Ito, A.; Moriguchi, T. Involvement of EARLY BUD-BREAK, an AP2/ERF transcription factor gene, in bud break in Japanese pear (Pyrus pyrifolia Nakai) lateral flower buds: Expression, histone modifications and possible target genes. Plant Cell Physiol. 2016, 57, 1038–1047. [Google Scholar] [CrossRef] [PubMed]
- Azeez, A.; Zhao, Y.C.; Singh, R.K.; Yordanov, Y.S.; Dash, M.; Miskolczi, P.; Stojkovič, K.; Strauss, S.H.; Bhalerao, R.P.; Busov, V.B. EARLY BUD-BREAK 1 and EARLY BUD-BREAK 3 control resumption of poplar growth after winter dormancy. Nat. Commun. 2021, 12, 1123. [Google Scholar] [CrossRef]
- Voogd, C.; Brian, L.A.; Wu, R.; Wang, T.; Allan, A.C.; Varkonyi-Gasic, E. A MADS-box gene with similarity to FLC is induced by cold and correlated with epigenetic changes to control budbreak in kiwifruit. New Phytol. 2022, 233, 2111–2126. [Google Scholar] [CrossRef]
- Canton, M.; Forestan, C.; Marconi, G.; Carrera, E.; Bonghi, C.; Varotto, S. Evidence of chromatin and transcriptional dynamics for cold development in peach flower bud. New Phytol. 2022, 236, 974–988. [Google Scholar] [CrossRef] [PubMed]
- Yang, Q.; Wu, X.; Gao, Y.; Ni, J.; Li, J.; Pei, Z.; Bai, S.; Teng, Y. PpyABF3 recruits the COMPASS-like complex to regulate bud dormancy maintenance via integrating ABA signaling and GA catabolism. New Phytol. 2023, 237, 192–203. [Google Scholar] [CrossRef]
- Zhou, H.; Zhang, W.; Sheng, Y.; Qiu, K.; Liao, L.; Shi, P.; Xie, Q.; Pan, H.; Zhang, J.; Han, Y. A large-scale behavior of allelic dropout and imbalance caused by DNA methylation changes in an early-ripening bud sport of peach. New Phytol. 2023, 239, 13–18. [Google Scholar] [CrossRef]
- Conde, D.; Moreno-Cortés, A.; Dervinis, C.; Ramos-Sánchez, J.M.; Kirst, M.; Perales, M.; González-Melendi, P.; Allona, I. Overexpression of DEMETER, a DNA demethylase, promotes early apical bud maturation in poplar. Plant Cell Environ. 2017, 40, 2806–2819. [Google Scholar] [CrossRef] [PubMed]
- Rothkegel, K.; Sandoval, P.; Soto, E.; Ulloa, L.; Riveros, A.; Lillo-Carmona, V.; Cáceres-Molina, J.; Almeida, A.M.; Meneses, C. Dormant but active: Chilling accumulation modulates the epigenome and transcriptome of Prunus avium during bud dormancy. Front. Plant Sci. 2020, 11, 1115. [Google Scholar] [CrossRef] [PubMed]
- Veerabagu, M.; van der Schoot, C.; Turečková, V.; Tarkowská, D.; Strnad, M.; Rinne, P.L. Light on perenniality: Para-dormancy is based on ABA-GA antagonism and endo-dormancy on the shutdown of GA biosynthesis. Plant Cell Environ. 2023, 46, 1785–1804. [Google Scholar] [CrossRef] [PubMed]
- Agustí, M.; Mesejo, C.; Muñoz-Fambuena, N.; Vera-Sirera, F.; de Lucas, M.; Martínez-Fuentes, A.; Reig, C.; Iglesias, D.J.; Primo-Millo, E.; Blázquez, M.A. Fruit-dependent epigenetic regulation of flowering in Citrus. New Phytol. 2020, 225, 376–384. [Google Scholar] [CrossRef]
- Kumar, G.; Rattan, U.K.; Singh, A.K. Chilling-mediated DNA methylation changes during dormancy and its release reveal the importance of epigenetic regulation during winter dormancy in apple (Malus × domestica Borkh.). PLoS ONE 2016, 11, e0149934. [Google Scholar] [CrossRef]
- Prudencio, Á.S.; Werner, O.; Martínez-García, P.J.; Dicenta, F.; Ros, R.M.; Martínez-Gómez, P. DNA methylation analysis of dormancy release in almond (Prunus dulcis) flower buds using epi-genotyping by sequencing. Int. J. Mol. Sci. 2018, 19, 3542. [Google Scholar] [CrossRef]
- Xin, H.; Zhang, Y.; Wang, X.; Liu, C.; Feng, W.; Gai, S. Morphological, anatomical and DNA methylation changes of tree peony buds during chilling induced dormancy release. Plant Physiol. Biochem. 2019, 144, 64–72. [Google Scholar] [CrossRef]
- Zhang, Y.; Zhang, T.; Si, F.; Wang, X.; Liu, C.; Yuan, Y.; Feng, W.; Gai, S. Changes of dna methylation patterns reveal epigenetic modification of dormancy release-related genes is induced by chilling in tree peony. DNA Cell Biol. 2021, 40, 606–617. [Google Scholar] [CrossRef]
- Rothkegel, K.; Sánchez, E.; Montes, C.; Greve, M.; Tapia, S.; Bravo, S.; Prieto, H.; Almeida, A.M. DNA methylation and small interference RNAs participate in the regulation of MADS-box genes involved in dormancy in sweet cherry (Prunus avium L.). Tree Physiol. 2017, 37, 1739–1751. [Google Scholar] [CrossRef]
- Santamaría, M.; Hasbún, R.; Valera, M.; Meijón, M.; Valledor, L.; Rodríguez, J.L.; Toorop, P.E.; Cañal, M.; Rodríguez, R. Acetylated H4 histone and genomic DNA methylation patterns during bud set and bud burst in Castanea sativa. J. Plant Physiol. 2009, 166, 1360–1369. [Google Scholar] [CrossRef]
- Saito, T.; Bai, S.; Imai, T.; Ito, A.; Nakajima, I.; Moriguchi, T. Histone modification and signalling cascade of the dormancy-associated MADS-box gene, PpMADS 13-1, in J apanese pear (Pyrus pyrifolia) during endodormancy. Plant Cell Environ. 2015, 38, 1157–1166. [Google Scholar] [CrossRef]
- Gao, Y.; Yang, Q.; Yan, X.; Wu, X.; Yang, F.; Li, J.; Wei, J.; Ni, J.; Ahmad, M.; Bai, S. High-quality genome assembly of’Cuiguan’pear (Pyrus pyrifolia) as a reference genome for identifying regulatory genes and epigenetic modifications responsible for bud dormancy. Hortic. Res. 2021, 8, 197. [Google Scholar] [CrossRef]
- Li, L.; Liu, J.; Liang, Q.; Zhang, Y.; Kang, K.; Wang, W.; Feng, Y.; Wu, S.; Yang, C.; Li, Y. Genome-wide analysis of long noncoding RNAs affecting floral bud dormancy in pears in response to cold stress. Tree Physiol. 2021, 41, 771–790. [Google Scholar] [CrossRef]
- Yu, J.; Bennett, D.; Dardick, C.; Zhebentyayeva, T.; Abbott, A.G.; Liu, Z.; Staton, M.E. Genome-wide changes of regulatory non-coding RNAs reveal pollen development initiated at ecodormancy in peach. Front. Mol. Biosci. 2021, 8, 612881. [Google Scholar] [CrossRef] [PubMed]
- Ding, Q.; Zeng, J.; He, X.-Q. Deep sequencing on a genome-wide scale reveals diverse stage-specific microRNAs in cambium during dormancy-release induced by chilling in poplar. BMC Plant Biol. 2014, 14, 267. [Google Scholar] [CrossRef] [PubMed]
- Soto, E.; Sanchez, E.; Nuñez, C.; Montes, C.; Rothkegel, K.; Andrade, P.; Prieto, H.; Almeida, A.M. Small RNA Differential Expression Analysis Reveals miRNAs Involved in Dormancy Progression in Sweet Cherry Floral Buds. Plants 2022, 11, 2396. [Google Scholar] [CrossRef] [PubMed]
- Garighan, J.; Dvorak, E.; Estevan, J.; Loridon, K.; Huettel, B.; Sarah, G.; Farrera, I.; Leclercq, J.; Grynberg, P.; Coiti Togawa, R. The identification of small RNAs differentially expressed in apple buds reveals a potential role of the miR159-MYB regulatory module during dormancy. Plants 2021, 10, 2665. [Google Scholar] [CrossRef] [PubMed]
- Smita, S.; Robben, M.; Deuja, A.; Accerbi, M.; Green, P.J.; Subramanian, S.; Fennell, A. Integrative analysis of gene expression and miRNAs reveal biological pathways associated with bud paradormancy and endodormancy in grapevine. Plants 2021, 10, 669. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Wang, Y.; Gao, X.; Liu, C.; Gai, S. Identification and characterization of microRNAs in tree peony during chilling induced dormancy release by high-throughput sequencing. Sci. Rep. 2018, 8, 4537. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Gao, L.; Wang, Y.; Niu, D.; Yuan, Y.; Liu, C.; Zhan, X.; Gai, S. Dual functions of PsmiR172b-PsTOE3 module in dormancy release and flowering in tree peony (Paeonia suffruticosa). Hortic. Res. 2023, 10, uhad033. [Google Scholar] [CrossRef] [PubMed]
- Conde, D.; Le Gac, A.L.; Perales, M.; Dervinis, C.; Kirst, M.; Maury, S.; González-Melendi, P.; Allona, I. Chilling-responsive DEMETER-LIKE DNA demethylase mediates in poplar bud break. Plant Cell Environ. 2017, 40, 2236–2249. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Si, F.; Wang, Y.; Liu, C.; Zhang, T.; Yuan, Y.; Gai, S. Application of 5-azacytidine induces DNA hypomethylation and accelerates dormancy release in buds of tree peony. Plant Physiol. Biochem. 2020, 147, 91–100. [Google Scholar] [CrossRef]
- Bai, S.; Saito, T.; Ito, A.; Tuan, P.A.; Xu, Y.; Teng, Y.; Moriguchi, T. Small RNA and PARE sequencing in flower bud reveal the involvement of sRNAs in endodormancy release of Japanese pear (Pyrus pyrifolia’Kosui’). BMC Genom. 2016, 17, 230. [Google Scholar] [CrossRef]
- Liu, N.; Jiang, Y.; Zhu, T.; Li, Z.; Sui, S. Small RNA and Degradome Sequencing in Floral Bud Reveal Roles of miRNAs in Dormancy Release of Chimonanthus praecox. Int. J. Mol. Sci. 2023, 24, 4210. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Epigenetic Modification | Mechanism of Gene Expression Regulation | References |
|---|---|---|---|
| Prunus persica | DNA methylation | bud early-ripening associates with changes of DNA methylation. | [95] |
| Populus, Prunus avium | DNA methylation | global DNA methylation during dormancy–growth cycle. | [96,97] |
| Populus | DNA methylation | DNA methylation represses GA3ox2 expression. | [98] |
| Citrus | DNA methylation | DNA methylation represses CcMADS19. | [99] |
| Malus domestica, Prunus dulcis, Paeonia sufruticosa | DNA methylation | DNA methylation is linked to chilling responses. | [100,101,102,103] |
| Prunus avium | DNA methylation | DNA methylation and siRNA are involved in the repression of PavMADS1. | [104] |
| Castanea sativa | Histone modifications | H4ac is involved in bud set and bud burst. | [105] |
| Prunus persica | Histone modifications | H3K4me3 and H3ac is related with DAM6 activation. | [86] |
| Prunus persica | Histone modifications | The dormancy release induces down-regulation of PpeS6PDH, which is associated with a low level of H3K4me3 and a high H3K27me3 level. | [89] |
| Prunus persica | Histone modifications | H3K27me3 is involved in the repression DAM1 and DAM5. | [87] |
| Prunus persica | Histone modifications | changes of H3K4me3 and H3K27me3 levels are involved in the regulation of dormancy. | [93] |
| Prunus persica | Histone modifications | H3K27me3 represses PpSVP1, PpDAM1, and PpNCED1. | [88] |
| Pyrus pyrifolia | Histone modifications | reduction of H3K4me3 and loss of H2A.Z are associated with endodormancy. | [106] |
| Pyrus pyrifolia | Histone modifications | H3K4me3 enhances the expression of PpEBB, DAM, PpyGA2OX1, NAC88, and CYCJ18. | [90,94,107] |
| Populus | Histone modifications | H3K27me3 represses EBB3 expression. | [91] |
| Actinidia chinensis, Citrus, Malus domestica, Prunus avium | Histone modifications | H3K4me3 and H3ac enhances the expression of dormancy-related genes (e.g., DAMs). | [60,84,92,99,108] |
| Prunus persica | non-coding RNAs | small RNA accumulation pattern is associated with DAMs expression. | [87] |
| Prunus persica, Pyrus pyrifolia | non-coding RNAs | global identification of noncoding RNAs related to dormancy regulation. | [108,109] |
| Pyrus pyrifolia | non-coding RNAs | miR6390 targets PpDAM and regulates PpFT2 expression. | [68] |
| Populus | non-coding RNAs | identification of dormancy-related small RNAs. | [110] |
| Prunus avium | non-coding RNAs | miR156/SPL, miR172/AP2, and miR166/ATBH15 are involved in dormancy regulation. | [111] |
| Malus domestica | non-coding RNAs | identifcation miR159a-MYB involved in dormancy regulation. | [112] |
| Vitis vinifera | non-coding RNAs | miRNA156, 167, and 1863 are involved in endodormancy | [113] |
| Paeonia sufruticosa | non-coding RNAs | PsmiR172b represses PsTOE3 during bud dormancy release. | [114,115] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chu, X.; Wang, M.; Fan, Z.; Li, J.; Yin, H. Molecular Mechanisms of Seasonal Gene Expression in Trees. Int. J. Mol. Sci. 2024, 25, 1666. https://doi.org/10.3390/ijms25031666
Chu X, Wang M, Fan Z, Li J, Yin H. Molecular Mechanisms of Seasonal Gene Expression in Trees. International Journal of Molecular Sciences. 2024; 25(3):1666. https://doi.org/10.3390/ijms25031666
Chicago/Turabian StyleChu, Xian, Minyan Wang, Zhengqi Fan, Jiyuan Li, and Hengfu Yin. 2024. "Molecular Mechanisms of Seasonal Gene Expression in Trees" International Journal of Molecular Sciences 25, no. 3: 1666. https://doi.org/10.3390/ijms25031666