
Multi-Omics Research Strategies for Psoriasis and Atopic Dermatitis

Abstract
:1. Introduction
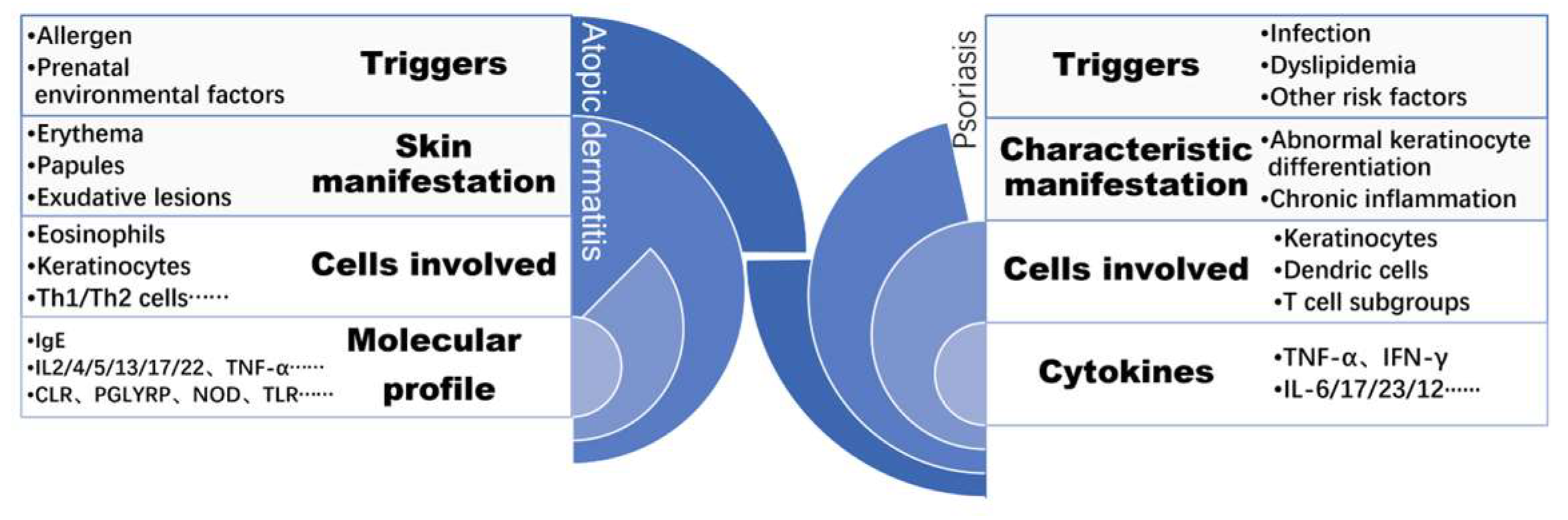
2. The Pathogenesis of Psoriasis and AD
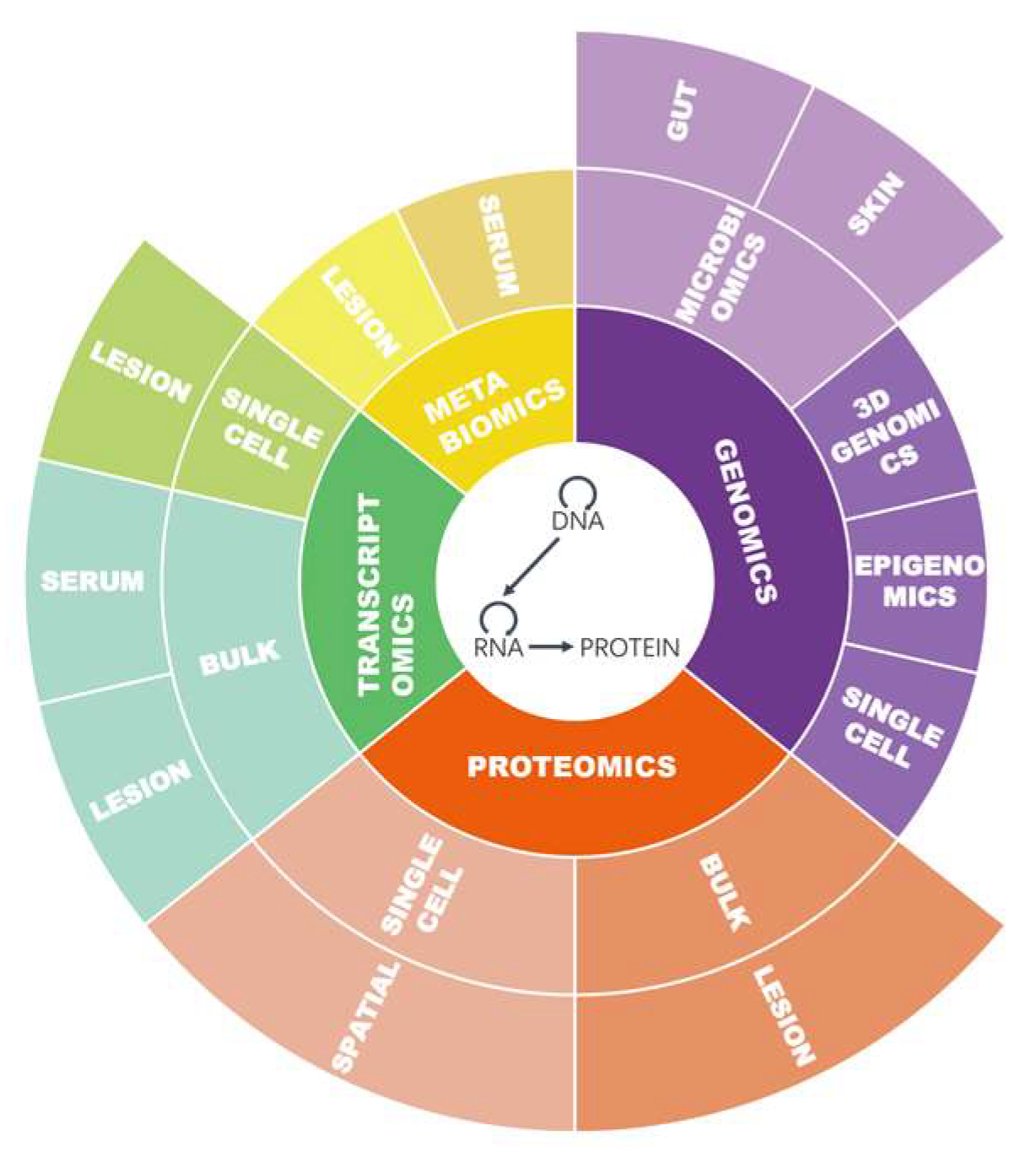
3. Omics Approaches Applied in Psoriasis and AD
3.1. Databases
3.2. Omics Approaches and Biomarkers
3.2.1. Genomics
Genome Analysis
Epigenomics
3D Genomics
3.2.2. Transcriptomics
3.2.3. Proteomics
3.2.4. Metabolomics
3.2.5. Microbiomics
4. Multi-Omics Strategies
4.1. The Omics-First Strategy
4.1.1. Parallel Research for Prioritizing Biomarkers
4.1.2. Subsequent Research for Further Comprehension
Genomics and Transcriptomics
Transcriptomics and Metabolomics
Proteomics and Metabolomics
Microbiomics and Transcriptomics
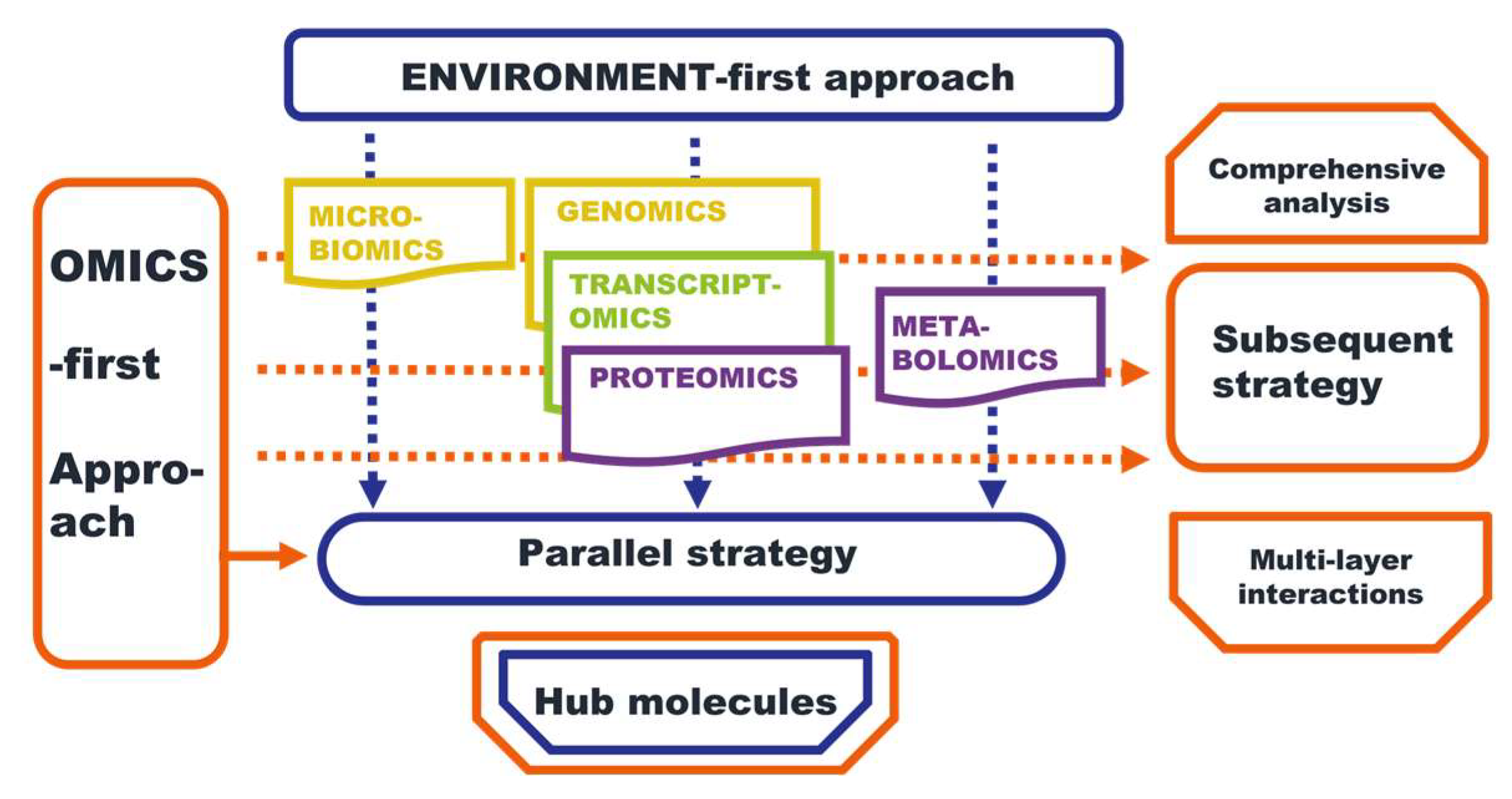
4.2. The Environment-First Approach
5. Tools for Integrating Omics Data
6. Future Perspective
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Griffiths, C.E.M.; Armstrong, A.W.; Gudjonsson, J.E.; Barker, J.N.W.N. Psoriasis. Lancet 2021, 397, 1301–1315. [Google Scholar] [CrossRef]
- Ständer, S. Atopic Dermatitis. N. Engl. J. Med. 2021, 384, 1136–1143. [Google Scholar] [CrossRef] [PubMed]
- Griffiths, C.E.; Barker, J.N. Pathogenesis and clinical features of psoriasis. Lancet 2007, 370, 263–271. [Google Scholar] [CrossRef] [PubMed]
- Nickoloff, B.J. Cracking the cytokine code in psoriasis. Nat. Med. 2007, 13, 242–244. [Google Scholar] [CrossRef] [PubMed]
- Crick, F. Central dogma of molecular biology. Nature 1970, 227, 561–563. [Google Scholar] [CrossRef] [PubMed]
- Blackstock, W.P.; Weir, M.P. Proteomics: Quantitative and physical mapping of cellular proteins. Trends Biotechnol. 1999, 17, 121–127. [Google Scholar]
- Venkatesh, A.; Patel, S.K.; Ray, S.; Shastri, J.; Chatterjee, G.; Kochar, S.K.; Patankar, S.; Srivastava, S. Proteomics of Plasmodium vivax malaria: New insights, progress and potential. Expert Rev. Proteom. 2016, 13, 771–782. [Google Scholar] [CrossRef]
- Kennedy-Crispin, M.; Billick, E.; Mitsui, H.; Gulati, N.; Fujita, H.; Gilleaudeau, P.; Sullivan-Whalen, M.; Johnson-Huang, L.M.; Suárez-Fariñas, M.; Krueger, J.G. Human keratinocytes’ response to injury upregulates CCL20 and other genes linking innate and adaptive immunity. J. Investig. Dermatol. 2012, 132, 105–113. [Google Scholar] [CrossRef]
- Buckley, L.H.; Xiao, R.; Perman, M.J.; Grossman, A.B.; Weiss, P.F. Psoriasis Associated With Tumor Necrosis Factor Inhibitors in Children With Inflammatory Diseases. Arthritis Care Res. 2021, 73, 215–220. [Google Scholar] [CrossRef]
- Renne, J.; Schäfer, V.; Werfel, T.; Wittmann, M. Interleukin-1 from epithelial cells fosters T cell-dependent skin inflammation. Br. J. Dermatol. 2010, 162, 1198–1205. [Google Scholar] [CrossRef]
- Hänsel, A.; Günther, C.; Ingwersen, J.; Starke, J.; Schmitz, M.; Bachmann, M.; Meurer, M.; Rieber, E.P.; Schäkel, K. Human slan (6-sulfo LacNAc) dendritic cells are inflammatory dermal dendritic cells in psoriasis and drive strong TH17/TH1 T-cell responses. J. Allergy Clin. Immunol. 2011, 127, 787–794.e9. [Google Scholar] [CrossRef] [PubMed]
- Nestle, F.O.; Turka, L.A.; Nickoloff, B.J. Characterization of dermal dendritic cells in psoriasis. Autostimulation of T lymphocytes and induction of Th1 type cytokines. J. Clin. Investig. 1994, 94, 202–209. [Google Scholar] [CrossRef] [PubMed]
- Rendon, A.; Schäkel, K. Psoriasis Pathogenesis and Treatment. Int. J. Mol. Sci. 2019, 20, 1475. [Google Scholar] [CrossRef]
- David Boothe, W.; Tarbox, J.A.; Tarbox, M.B. Atopic Dermatitis: Pathophysiology. Adv. Exp. Med. Biol. 2017, 1027, 21–37. [Google Scholar] [CrossRef]
- Sroka-Tomaszewska, J.; Trzeciak, M. Molecular Mechanisms of Atopic Dermatitis Pathogenesis. Int. J. Mol. Sci. 2021, 22, 4130. [Google Scholar] [CrossRef] [PubMed]
- Matsunaga, M.C.; Yamauchi, P.S. IL-4 and IL-13 Inhibition in Atopic Dermatitis. J. Drugs Dermatol. 2016, 15, 925–929. [Google Scholar]
- Gene Expression Omnibus Repository of National Library of Medicine. Available online: https://www.ncbi.nlm.nih.gov/geo/ (accessed on 7 April 2023).
- Human Gene Mutation Database. Available online: https://www.hgmd.cf.ac.uk/ac/index.php (accessed on 7 April 2023).
- Ensembl. Available online: http://ensembl.org/index.html (accessed on 7 April 2023).
- ClinVar. Available online: https://www.ncbi.nlm.nih.gov/clinvar/ (accessed on 7 April 2023).
- The Epigenetic Equivalent of FORGE, Using EWAS Rather than GWAS Data. Available online: https://eforge.altiusinstitute.org/ (accessed on 7 April 2023).
- The Human microRNA Disease Database. Available online: http://www.cuilab.cn/hmdd (accessed on 7 April 2023).
- circRNADisease. Available online: http://cgga.org.cn:9091/circRNADisease/ (accessed on 7 April 2023).
- lncRNADisease. Available online: http://www.rnanut.net/lncrnadisease/ (accessed on 7 April 2023).
- Uniprot. Available online: https://www.uniprot.org/ (accessed on 7 April 2023).
- Simple Modular Architecture Research Tool. Available online: http://smart.embl-heidelberg.de/ (accessed on 7 April 2023).
- Post Translational Modification Database. Available online: http://ptmd.biocuckoo.org/ (accessed on 7 April 2023).
- The Human Metabolome Database. Available online: https://hmdb.ca/ (accessed on 7 April 2023).
- Pathogen Host Interactions Database. Available online: http://www.phi-base.org/ (accessed on 7 April 2023).
- Human Microbiome Project Data Portal. Available online: https://portal.hmpdacc.org/ (accessed on 7 April 2023).
- Stenson, P.D.; Ball, E.V.; Mort, M.; Phillips, A.D.; Shiel, J.A.; Thomas, N.S.T.; Abeysinghe, S.; Krawczak, M.; Cooper, D.N. Human Gene Mutation Database (HGMD): 2003 update. Hum. Mutat. 2003, 21, 577–581. [Google Scholar] [CrossRef]
- Rappaport, N.; Twik, M.; Plaschkes, I.; Nudel, R.; Iny Stein, T.; Levitt, J.; Gershoni, M.; Morrey, C.P.; Safran, M.; Lancet, D. MalaCards: An amalgamated human disease compendium with diverse clinical and genetic annotation and structured search. Nucleic Acids Res. 2017, 45, D877–D887. [Google Scholar] [CrossRef]
- National Center for Biotechnology Information (NCBI) [Internet]. Bethesda (MD): National Library of Medicine (US), National Center for Biotechnology Information. Available online: https://www.ncbi.nlm.nih.gov/ (accessed on 7 April 2023).
- The UniProt Consortium UniProt: The Universal Protein Knowledgebase in 2023. Available online: https://www.uniprot.org/uniprotkb?query=psoriasis (accessed on 28 February 2023).
- Wishart, D.S.; Guo, A.; Oler, E.; Wang, F.; Anjum, A.; Peters, H.; Dizon, R.; Sayeeda, Z.; Tian, S.; Lee, B.L.; et al. HMDB 5.0: The Human Metabolome Database for 2022. Nucleic Acids Res. 2022, 50, D622–D631. [Google Scholar] [CrossRef]
- Farag, A.; Shoaib, M.; Labeeb, A.; Sleem, A.; Hussien, H.; Elshaib, M.; Hanout, H. S100A8 (rs3806232) gene polymorphism and S100A8 serum level in psoriasis vulgaris patients: A preliminary study. J. Cosmet. Dermatol. 2022, 21, 4974–4982. [Google Scholar] [CrossRef]
- Wang, L.; Yu, X.; Wu, C.; Zhu, T.; Wang, W.; Zheng, X.; Jin, H. RNA sequencing-based longitudinal transcriptomic profiling gives novel insights into the disease mechanism of generalized pustular psoriasis. BMC Med. Genom. 2018, 11, 52. [Google Scholar] [CrossRef] [PubMed]
- Tsoi, L.C.; Rodriguez, E.; Stölzl, D.; Wehkamp, U.; Sun, J.; Gerdes, S.; Sarkar, M.K.; Hübenthal, M.; Zeng, C.; Uppala, R.; et al. Progression of acute-to-chronic atopic dermatitis is associated with quantitative rather than qualitative changes in cytokine responses. J. Allergy Clin. Immunol. 2020, 145, 1406–1415. [Google Scholar] [CrossRef] [PubMed]
- Gittler, J.K.; Shemer, A.; Suárez-Fariñas, M.; Fuentes-Duculan, J.; Gulewicz, K.J.; Wang, C.Q.F.; Mitsui, H.; Cardinale, I.; de Guzman Strong, C.; Krueger, J.G.; et al. Progressive activation of T(H)2/T(H)22 cytokines and selective epidermal proteins characterizes acute and chronic atopic dermatitis. J. Allergy Clin. Immunol. 2012, 130, 1344–1354. [Google Scholar] [CrossRef] [PubMed]
- Schonthaler, H.B.; Guinea-Viniegra, J.; Wculek, S.K.; Ruppen, I.; Ximénez-Embún, P.; Guío-Carrión, A.; Navarro, R.; Hogg, N.; Ashman, K.; Wagner, E.F. S100A8-S100A9 protein complex mediates psoriasis by regulating the expression of complement factor C3. Immunity 2013, 39, 1171–1181. [Google Scholar] [CrossRef]
- Babić, Ž.; Sabolić Pipinić, I.; Varnai, V.M.; Kežić, S.; Macan, J. Associations of TNFα-308G>A, TNFα-238G>A, IL-1α-889C>T and IL-10 -1082G>A Genetic Polymorphisms with Atopic Diseases: Asthma, Rhinitis and Dermatitis. Int. Arch. Allergy Immunol. 2016, 169, 231–240. [Google Scholar] [CrossRef]
- Lowes, M.A.; Suárez-Fariñas, M.; Krueger, J.G. Immunology of psoriasis. Annu. Rev. Immunol. 2014, 32, 227–255. [Google Scholar] [CrossRef]
- Pasquali, L.; Srivastava, A.; Meisgen, F.; Das Mahapatra, K.; Xia, P.; Xu Landén, N.; Pivarcsi, A.; Sonkoly, E. The Keratinocyte Transcriptome in Psoriasis: Pathways Related to Immune Responses, Cell Cycle and Keratinization. Acta Derm. Venereol. 2019, 99, 196–205. [Google Scholar] [CrossRef]
- Möbus, L.; Rodriguez, E.; Harder, I.; Stölzl, D.; Boraczynski, N.; Gerdes, S.; Kleinheinz, A.; Abraham, S.; Heratizadeh, A.; Handrick, C.; et al. Atopic dermatitis displays stable and dynamic skin transcriptome signatures. J. Allergy Clin. Immunol. 2021, 147, 213–223. [Google Scholar] [CrossRef]
- Rodrigues de Souza, I.; Savio de Araujo-Souza, P.; Morais Leme, D. Genetic variants affecting chemical mediated skin immunotoxicity. J. Toxicol. Environ. Health Part B Crit. Rev. 2022, 25, 43–95. [Google Scholar] [CrossRef]
- Hirota, T.; Takahashi, A.; Kubo, M.; Tsunoda, T.; Tomita, K.; Sakashita, M.; Yamada, T.; Fujieda, S.; Tanaka, S.; Doi, S.; et al. Genome-wide association study identifies eight new susceptibility loci for atopic dermatitis in the Japanese population. Nat. Genet. 2012, 44, 1222–1226. [Google Scholar] [CrossRef]
- Nanda, H.; Ponnusamy, N.; Odumpatta, R.; Jeyakanthan, J.; Mohanapriya, A. Exploring genetic targets of psoriasis using genome wide association studies (GWAS) for drug repurposing. 3 Biotech 2020, 10, 43. [Google Scholar] [CrossRef] [PubMed]
- Oetjen, L.K.; Mack, M.R.; Feng, J.; Whelan, T.M.; Niu, H.; Guo, C.J.; Chen, S.; Trier, A.M.; Xu, A.Z.; Tripathi, S.V.; et al. Sensory Neurons Co-opt Classical Immune Signaling Pathways to Mediate Chronic Itch. Cell 2017, 171, 217–228.e13. [Google Scholar] [CrossRef] [PubMed]
- Beck, L.A.; Thaçi, D.; Hamilton, J.D.; Graham, N.M.; Bieber, T.; Rocklin, R.; Ming, J.E.; Ren, H.; Kao, R.; Simpson, E.; et al. Dupilumab treatment in adults with moderate-to-severe atopic dermatitis. N. Engl. J. Med. 2014, 371, 130–139. [Google Scholar] [CrossRef] [PubMed]
- Brunner, P.M.; Suárez-Fariñas, M.; He, H.; Malik, K.; Wen, H.-C.; Gonzalez, J.; Chan, T.C.-C.; Estrada, Y.; Zheng, X.; Khattri, S.; et al. The atopic dermatitis blood signature is characterized by increases in inflammatory and cardiovascular risk proteins. Sci. Rep. 2017, 7, 8707. [Google Scholar] [CrossRef]
- Kõks, S.; Kingo, K.; Vabrit, K.; Rätsep, R.; Karelson, M.; Silm, H.; Vasar, E. Possible relations between the polymorphisms of the cytokines IL-19, IL-20 and IL-24 and plaque-type psoriasis. Genes Immun. 2005, 6, 407–415. [Google Scholar] [CrossRef]
- Vu, Y.H.; Hashimoto-Hachiya, A.; Takemura, M.; Yumine, A.; Mitamura, Y.; Nakahara, T.; Furue, M.; Tsuji, G. IL-24 Negatively Regulates Keratinocyte Differentiation Induced by Tapinarof, an Aryl Hydrocarbon Receptor Modulator: Implication in the Treatment of Atopic Dermatitis. Int. J. Mol. Sci. 2020, 21, 9412. [Google Scholar] [CrossRef]
- Méhul, B.; Laffet, G.; Séraïdaris, A.; Russo, L.; Fogel, P.; Carlavan, I.; Pernin, C.; Andres, P.; Queille-Roussel, C.; Voegel, J.J. Noninvasive proteome analysis of psoriatic stratum corneum reflects pathophysiological pathways and is useful for drug profiling. Br. J. Dermatol. 2017, 177, 470–488. [Google Scholar] [CrossRef]
- He, H.; Olesen, C.M.; Pavel, A.B.; Clausen, M.-L.; Wu, J.; Estrada, Y.; Zhang, N.; Agner, T.; Guttman-Yassky, E. Tape-Strip Proteomic Profiling of Atopic Dermatitis on Dupilumab Identifies Minimally Invasive Biomarkers. Front. Immunol. 2020, 11, 1768. [Google Scholar] [CrossRef]
- Traks, T.; Keermann, M.; Prans, E.; Karelson, M.; Loite, U.; Kõks, G.; Silm, H.; Kõks, S.; Kingo, K. Polymorphisms in IL36G gene are associated with plaque psoriasis. BMC Med. Genet. 2019, 20, 10. [Google Scholar] [CrossRef]
- Chiricozzi, A.; Guttman-Yassky, E.; Suárez-Fariñas, M.; Nograles, K.E.; Tian, S.; Cardinale, I.; Chimenti, S.; Krueger, J.G. Integrative responses to IL-17 and TNF-α in human keratinocytes account for key inflammatory pathogenic circuits in psoriasis. J. Investig. Dermatol. 2011, 131, 677–687. [Google Scholar] [CrossRef]
- Suárez-Fariñas, M.; Ungar, B.; Correa da Rosa, J.; Ewald, D.A.; Rozenblit, M.; Gonzalez, J.; Xu, H.; Zheng, X.; Peng, X.; Estrada, Y.D.; et al. RNA sequencing atopic dermatitis transcriptome profiling provides insights into novel disease mechanisms with potential therapeutic implications. J. Allergy Clin. Immunol. 2015, 135, 1218–1227. [Google Scholar] [CrossRef] [PubMed]
- Puig, L.; Julià, A.; Marsal, S. The pathogenesis and genetics of psoriasis. Actas Dermo Sifiliográficas 2014, 105, 535–545. [Google Scholar] [CrossRef] [PubMed]
- Capon, F.; Di Meglio, P.; Szaub, J.; Prescott, N.J.; Dunster, C.; Baumber, L.; Timms, K.; Gutin, A.; Abkevic, V.; Burden, A.D.; et al. Sequence variants in the genes for the interleukin-23 receptor (IL23R) and its ligand (IL12B) confer protection against psoriasis. Hum. Genet. 2007, 122, 201–206. [Google Scholar] [CrossRef] [PubMed]
- Capon, F.; Bijlmakers, M.-J.; Wolf, N.; Quaranta, M.; Huffmeier, U.; Allen, M.; Timms, K.; Abkevich, V.; Gutin, A.; Smith, R.; et al. Identification of ZNF313/RNF114 as a novel psoriasis susceptibility gene. Hum. Mol. Genet. 2008, 17, 1938–1945. [Google Scholar] [CrossRef] [PubMed]
- Hüffmeier, U.; Uebe, S.; Ekici, A.B.; Bowes, J.; Giardina, E.; Korendowych, E.; Juneblad, K.; Apel, M.; McManus, R.; Ho, P.; et al. Common variants at TRAF3IP2 are associated with susceptibility to psoriatic arthritis and psoriasis. Nat. Genet. 2010, 42, 996–999. [Google Scholar] [CrossRef]
- Wu, Y.; Zhang, Z.; Tao, L.; Chen, G.; Liu, F.; Wang, T.; Xue, F.; Chen, Y.; He, L.; Zheng, J.; et al. A high copy number of FCGR3B is associated with psoriasis vulgaris in Han Chinese. Dermatology 2014, 229, 70–75. [Google Scholar] [CrossRef]
- Prans, E.; Kingo, K.; Traks, T.; Silm, H.; Vasar, E.; Kõks, S. Copy number variations in IL22 gene are associated with Psoriasis vulgaris. Hum. Immunol. 2013, 74, 792–795. [Google Scholar] [CrossRef]
- Paternoster, L.; Standl, M.; Waage, J.; Baurecht, H.; Hotze, M.; Strachan, D.P.; Curtin, J.A.; Bønnelykke, K.; Tian, C.; Takahashi, A.; et al. Multi-ancestry genome-wide association study of 21,000 cases and 95,000 controls identifies new risk loci for atopic dermatitis. Nat. Genet. 2015, 47, 1449–1456. [Google Scholar] [CrossRef]
- Nedoszytko, B.; Reszka, E.; Gutowska-Owsiak, D.; Trzeciak, M.; Lange, M.; Jarczak, J.; Niedoszytko, M.; Jablonska, E.; Romantowski, J.; Strapagiel, D.; et al. Genetic and Epigenetic Aspects of Atopic Dermatitis. Int. J. Mol. Sci. 2020, 21, 6484. [Google Scholar] [CrossRef]
- Ruchusatsawat, K.; Wongpiyabovorn, J.; Shuangshoti, S.; Hirankarn, N.; Mutirangura, A. SHP-1 promoter 2 methylation in normal epithelial tissues and demethylation in psoriasis. J. Mol. Med. 2006, 84, 175–182. [Google Scholar] [CrossRef]
- Roberson, E.D.O.; Liu, Y.; Ryan, C.; Joyce, C.E.; Duan, S.; Cao, L.; Martin, A.; Liao, W.; Menter, A.; Bowcock, A.M. A subset of methylated CpG sites differentiate psoriatic from normal skin. J. Investig. Dermatol. 2012, 132, 583–592. [Google Scholar] [CrossRef] [PubMed]
- Zhang, P.; Zhao, M.; Liang, G.; Yin, G.; Huang, D.; Su, F.; Zhai, H.; Wang, L.; Su, Y.; Lu, Q. Whole-genome DNA methylation in skin lesions from patients with psoriasis vulgaris. J. Autoimmun. 2013, 41, 17–24. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez, E.; Baurecht, H.; Wahn, A.F.; Kretschmer, A.; Hotze, M.; Zeilinger, S.; Klopp, N.; Illig, T.; Schramm, K.; Prokisch, H.; et al. An integrated epigenetic and transcriptomic analysis reveals distinct tissue-specific patterns of DNA methylation associated with atopic dermatitis. J. Investig. Dermatol. 2014, 134, 1873–1883. [Google Scholar] [CrossRef]
- Zhang, P.; Su, Y.; Zhao, M.; Huang, W.; Lu, Q. Abnormal histone modifications in PBMCs from patients with psoriasis vulgaris. Eur. J. Dermatol. 2011, 21, 552–557. [Google Scholar] [CrossRef] [PubMed]
- Mu, Z.; Zhang, J. The Role of Genetics, the Environment, and Epigenetics in Atopic Dermatitis. Adv. Exp. Med. Biol. 2020, 1253, 107–140. [Google Scholar] [CrossRef] [PubMed]
- Kempfer, R.; Pombo, A. Methods for mapping 3D chromosome architecture. Nat. Rev. Genet. 2020, 21, 207–226. [Google Scholar] [CrossRef] [PubMed]
- Croft, J.A.; Bridger, J.M.; Boyle, S.; Perry, P.; Teague, P.; Bickmore, W.A. Differences in the localization and morphology of chromosomes in the human nucleus. J. Cell Biol. 1999, 145, 1119–1131. [Google Scholar] [CrossRef] [PubMed]
- Yasen, A.; Aini, A.; Wang, H.; Li, W.; Zhang, C.; Ran, B.; Tuxun, T.; Maimaitinijiati, Y.; Shao, Y.; Aji, T.; et al. Progress and applications of single-cell sequencing techniques. Infect. Genet. Evol. 2020, 80, 104198. [Google Scholar] [CrossRef]
- Gawad, C.; Koh, W.; Quake, S.R. Single-cell genome sequencing: Current state of the science. Nat. Rev. Genet. 2016, 17, 175–188. [Google Scholar] [CrossRef]
- Rao, S.S.P.; Huntley, M.H.; Durand, N.C.; Stamenova, E.K.; Bochkov, I.D.; Robinson, J.T.; Sanborn, A.L.; Machol, I.; Omer, A.D.; Lander, E.S.; et al. A 3D map of the human genome at kilobase resolution reveals principles of chromatin looping. Cell 2014, 159, 1665–1680. [Google Scholar] [CrossRef]
- Ray-Jones, H.; Duffus, K.; McGovern, A.; Martin, P.; Shi, C.; Hankinson, J.; Gough, O.; Yarwood, A.; Morris, A.P.; Adamson, A.; et al. Mapping DNA interaction landscapes in psoriasis susceptibility loci highlights KLF4 as a target gene in 9q31. BMC Biol. 2020, 18, 47. [Google Scholar] [CrossRef] [PubMed]
- Sahlén, P.; Spalinskas, R.; Asad, S.; Mahapatra, K.D.; Höjer, P.; Anil, A.; Eisfeldt, J.; Srivastava, A.; Nikamo, P.; Mukherjee, A.; et al. Chromatin interactions in differentiating keratinocytes reveal novel atopic dermatitis- and psoriasis-associated genes. J Allergy Clin. Immunol. 2021, 147, 1742–1752. [Google Scholar] [CrossRef] [PubMed]
- Gudjonsson, J.E.; Ding, J.; Johnston, A.; Tejasvi, T.; Guzman, A.M.; Nair, R.P.; Voorhees, J.J.; Abecasis, G.R.; Elder, J.T. Assessment of the psoriatic transcriptome in a large sample: Additional regulated genes and comparisons with in vitro models. J. Investig. Dermatol. 2010, 130, 1829–1840. [Google Scholar] [CrossRef] [PubMed]
- Keermann, M.; Kõks, S.; Reimann, E.; Prans, E.; Abram, K.; Kingo, K. Transcriptional landscape of psoriasis identifies the involvement of IL36 and IL36RN. BMC Genom. 2015, 16, 322. [Google Scholar] [CrossRef] [PubMed]
- Oestreicher, J.L.; Walters, I.B.; Kikuchi, T.; Gilleaudeau, P.; Surette, J.; Schwertschlag, U.; Dorner, A.J.; Krueger, J.G.; Trepicchio, W.L. Molecular classification of psoriasis disease-associated genes through pharmacogenomic expression profiling. The Pharm. J. 2001, 1, 272–287. [Google Scholar] [CrossRef]
- Xu, Y.; Ji, Y.; Lan, X.; Gao, X.; Chen, H.-D.; Geng, L. miR-203 contributes to IL-17-induced VEGF secretion by targeting SOCS3 in keratinocytes. Mol. Med. Rep. 2017, 16, 8989–8996. [Google Scholar] [CrossRef]
- Joyce, C.E.; Zhou, X.; Xia, J.; Ryan, C.; Thrash, B.; Menter, A.; Zhang, W.; Bowcock, A.M. Deep sequencing of small RNAs from human skin reveals major alterations in the psoriasis miRNAome. Hum. Mol. Genet. 2011, 20, 4025–4040. [Google Scholar] [CrossRef]
- Schwingen, J.; Kaplan, M.; Kurschus, F.C. Review-Current Concepts in Inflammatory Skin Diseases Evolved by Transcriptome Analysis: In-Depth Analysis of Atopic Dermatitis and Psoriasis. Int. J. Mol. Sci. 2020, 21, 699. [Google Scholar] [CrossRef]
- Quinn, S.R.; Mangan, N.E.; Caffrey, B.E.; Gantier, M.P.; Williams, B.R.G.; Hertzog, P.J.; McCoy, C.E.; O’Neill, L.A.J. The role of Ets2 transcription factor in the induction of microRNA-155 (miR-155) by lipopolysaccharide and its targeting by interleukin-10. J. Biol. Chem. 2014, 289, 4316–4325. [Google Scholar] [CrossRef]
- Nakamizo, S.; Dutertre, C.-A.; Khalilnezhad, A.; Zhang, X.M.; Lim, S.; Lum, J.; Koh, G.; Foong, C.; Yong, P.J.A.; Tan, K.J.; et al. Single-cell analysis of human skin identifies CD14+ type 3 dendritic cells co-producing IL1B and IL23A in psoriasis. J. Exp. Med. 2021, 218, e20202345. [Google Scholar] [CrossRef]
- Kim, J.; Lee, J.; Kim, H.J.; Kameyama, N.; Nazarian, R.; Der, E.; Cohen, S.; Guttman-Yassky, E.; Putterman, C.; Krueger, J.G. Single-cell transcriptomics applied to emigrating cells from psoriasis elucidate pathogenic versus regulatory immune cell subsets. J. Allergy Clin. Immunol. 2021, 148, 1281–1292. [Google Scholar] [CrossRef] [PubMed]
- Qie, C.; Jiang, J.; Liu, W.; Hu, X.; Chen, W.; Xie, X.; Liu, J. Single-cell RNA-Seq reveals the transcriptional landscape and heterogeneity of skin macrophages in Vsir-/- murine psoriasis. Theranostics 2020, 10, 10483–10497. [Google Scholar] [CrossRef] [PubMed]
- Reschke, R.; Shapiro, J.W.; Yu, J.; Rouhani, S.J.; Olson, D.J.; Zha, Y.; Gajewski, T.F. Checkpoint Blockade-Induced Dermatitis and Colitis Are Dominated by Tissue-Resident Memory T Cells and Th1/Tc1 Cytokines. Cancer Immunol. Res. 2022, 10, 1167–1174. [Google Scholar] [CrossRef] [PubMed]
- Huang, T.; Gao, Z.; Zhang, Y.; Fan, K.; Wang, F.; Li, Y.; Zhong, J.; Fan, H.Y.; Cao, Q.; Zhou, J.; et al. CRL4DCAF2 negatively regulates IL-23 production in dendritic cells and limits the development of psoriasis. J. Exp. Med. 2018, 215, 1999–2017. [Google Scholar] [CrossRef]
- Gong, X.; Wang, W. Profiles of Innate Immune Cell Infiltration and Related Core Genes in Psoriasis. BioMed Res. Int. 2021, 2021, 6656622. [Google Scholar] [CrossRef]
- Li, C.; Lu, Y.; Han, X. Identification of Effective Diagnostic Biomarkers and Immune Cell Infiltration in Atopic Dermatitis by Comprehensive Bioinformatics Analysis. Front. Mol. Biosci. 2022, 9, 917077. [Google Scholar] [CrossRef]
- Zeng, F.; Liu, H.; Lu, D.; Liu, Q.; Chen, H.; Zheng, F. Integrated analysis of gene expression profiles identifies transcription factors potentially involved in psoriasis pathogenesis. J. Cell. Biochem. 2019, 120, 12582–12594. [Google Scholar] [CrossRef]
- Williamson, J.C.; Scheipers, P.; Schwämmle, V.; Zibert, J.R.; Beck, H.C.; Jensen, O.N. A proteomics approach to the identification of biomarkers for psoriasis utilising keratome biopsy. J. Proteom. 2013, 94, 176–185. [Google Scholar] [CrossRef]
- Zhou, X.; Chen, Y.; Cui, L.; Shi, Y.; Guo, C. Advances in the pathogenesis of psoriasis: From keratinocyte perspective. Cell Death Dis. 2022, 13, 81. [Google Scholar] [CrossRef]
- Nicholson, J.K.; Lindon, J.C.; Holmes, E. ‘Metabonomics’: Understanding the metabolic responses of living systems to pathophysiological stimuli via multivariate statistical analysis of biological NMR spectroscopic data. Xenobiotica 1999, 29, 1181–1189. [Google Scholar] [CrossRef]
- Ilves, L.; Ottas, A.; Kaldvee, B.; Abram, K.; Soomets, U.; Zilmer, M.; Jaks, V.; Kingo, K. Metabolomic Differences between the Skin and Blood Sera of Atopic Dermatitis and Psoriasis. Int. J. Mol. Sci. 2022, 23, 13001. [Google Scholar] [CrossRef] [PubMed]
- Dutkiewicz, E.P.; Hsieh, K.-T.; Wang, Y.-S.; Chiu, H.-Y.; Urban, P.L. Hydrogel Micropatch and Mass Spectrometry-Assisted Screening for Psoriasis-Related Skin Metabolites. Clin. Chem. 2016, 62, 1120–1128. [Google Scholar] [CrossRef]
- Kang, H.; Li, X.; Zhou, Q.; Quan, C.; Xue, F.; Zheng, J.; Yu, Y. Exploration of candidate biomarkers for human psoriasis based on gas chromatography-mass spectrometry serum metabolomics. Br. J. Dermatol. 2017, 176, 713–722. [Google Scholar] [CrossRef] [PubMed]
- Alonso, A.; Julià, A.; Vinaixa, M.; Domènech, E.; Fernández-Nebro, A.; Cañete, J.D.; Ferrándiz, C.; Tornero, J.; Gisbert, J.P.; Nos, P.; et al. Urine metabolome profiling of immune-mediated inflammatory diseases. BMC Med. 2016, 14, 133. [Google Scholar] [CrossRef]
- Klindworth, A.; Pruesse, E.; Schweer, T.; Peplies, J.; Quast, C.; Horn, M.; Glöckner, F.O. Evaluation of general 16S ribosomal RNA gene PCR primers for classical and next-generation sequencing-based diversity studies. Nucleic Acids Res. 2013, 41, e1. [Google Scholar] [CrossRef]
- Alekseyenko, A.V.; Perez-Perez, G.I.; De Souza, A.; Strober, B.; Gao, Z.; Bihan, M.; Li, K.; Methé, B.A.; Blaser, M.J. Community differentiation of the cutaneous microbiota in psoriasis. Microbiome 2013, 1, 31. [Google Scholar] [CrossRef]
- Gao, Z.; Tseng, C.-h.; Strober, B.E.; Pei, Z.; Blaser, M.J. Substantial alterations of the cutaneous bacterial biota in psoriatic lesions. PLoS ONE 2008, 3, e2719. [Google Scholar] [CrossRef]
- Kong, H.H.; Oh, J.; Deming, C.; Conlan, S.; Grice, E.A.; Beatson, M.A.; Nomicos, E.; Polley, E.C.; Komarow, H.D.; Murray, P.R.; et al. Temporal shifts in the skin microbiome associated with disease flares and treatment in children with atopic dermatitis. Genome Res. 2012, 22, 850–859. [Google Scholar] [CrossRef]
- Chng, K.R.; Tay, A.S.L.; Li, C.; Ng, A.H.Q.; Wang, J.; Suri, B.K.; Matta, S.A.; McGovern, N.; Janela, B.; Wong, X.F.C.C.; et al. Whole metagenome profiling reveals skin microbiome-dependent susceptibility to atopic dermatitis flare. Nat. Microbiol. 2016, 1, 16106. [Google Scholar] [CrossRef]
- Choi, J.-Y.; Kim, H.; Koo, H.-Y.-R.; You, J.; Yu, D.-S.; Lee, Y.-B.; Lee, M. Severe Scalp Psoriasis Microbiome Has Increased Biodiversity and Relative Abundance of Pseudomonas Compared to Mild Scalp Psoriasis. J. Clin. Med. 2022, 11, 7133. [Google Scholar] [CrossRef]
- Zhang, X.; Shi, L.; Sun, T.; Guo, K.; Geng, S. Dysbiosis of gut microbiota and its correlation with dysregulation of cytokines in psoriasis patients. BMC Microbiol. 2021, 21, 78. [Google Scholar] [CrossRef] [PubMed]
- Todberg, T.; Egeberg, A.; Zachariae, C.; Sørensen, N.; Pedersen, O.; Skov, L. Patients with psoriasis have a dysbiotic taxonomic and functional gut microbiota. Br. J. Dermatol. 2022, 187, 89–98. [Google Scholar] [CrossRef] [PubMed]
- Hasin, Y.; Seldin, M.; Lusis, A. Multi-omics approaches to disease. Genome Biol. 2017, 18, 83. [Google Scholar] [CrossRef]
- Harden, J.L.; Lewis, S.M.; Pierson, K.C.; Suárez-Fariñas, M.; Lentini, T.; Ortenzio, F.S.; Zaba, L.C.; Goldbach-Mansky, R.; Bowcock, A.M.; Lowes, M.A. CARD14 expression in dermal endothelial cells in psoriasis. PLoS ONE 2014, 9, e111255. [Google Scholar] [CrossRef] [PubMed]
- Elnabawi, Y.A.; Garshick, M.S.; Tawil, M.; Barrett, T.J.; Fisher, E.A.; Lo Sicco, K.; Neimann, A.L.; Scher, J.U.; Krueger, J.; Berger, J.S. CCL20 in psoriasis: A potential biomarker of disease severity, inflammation, and impaired vascular health. J. Am. Acad. Dermatol. 2021, 84, 913–920. [Google Scholar] [CrossRef]
- Federico, A.; Pavel, A.; Möbus, L.; McKean, D.; Del Giudice, G.; Fortino, V.; Niehues, H.; Rastrick, J.; Eyerich, K.; Eyerich, S.; et al. The integration of large-scale public data and network analysis uncovers molecular characteristics of psoriasis. Hum. Genom. 2022, 16, 62. [Google Scholar] [CrossRef] [PubMed]
- Ghosh, D.; Bernstein, J.A.; Khurana Hershey, G.K.; Rothenberg, M.E.; Mersha, T.B. Leveraging Multilayered "Omics" Data for Atopic Dermatitis: A Road Map to Precision Medicine. Front. Immunol. 2018, 9, 2727. [Google Scholar] [CrossRef]
- Piruzian, E.; Bruskin, S.; Ishkin, A.; Abdeev, R.; Moshkovskii, S.; Melnik, S.; Nikolsky, Y.; Nikolskaya, T. Integrated network analysis of transcriptomic and proteomic data in psoriasis. BMC Syst. Biol. 2010, 4, 41. [Google Scholar] [CrossRef]
- Wang, X.; Kaiser, H.; Kvist-Hansen, A.; McCauley, B.D.; Skov, L.; Hansen, P.R.; Becker, C. IL-17 Pathway Members as Potential Biomarkers of Effective Systemic Treatment and Cardiovascular Disease in Patients with Moderate-to-Severe Psoriasis. Int. J. Mol. Sci. 2022, 23, 555. [Google Scholar] [CrossRef]
- Khunsriraksakul, C.; McGuire, D.; Sauteraud, R.; Chen, F.; Yang, L.; Wang, L.; Hughey, J.; Eckert, S.; Dylan Weissenkampen, J.; Shenoy, G.; et al. Integrating 3D genomic and epigenomic data to enhance target gene discovery and drug repurposing in transcriptome-wide association studies. Nat. Commun. 2022, 13, 3258. [Google Scholar] [CrossRef]
- Töröcsik, D.; Weise, C.; Gericke, J.; Szegedi, A.; Lucas, R.; Mihaly, J.; Worm, M.; Rühl, R. Transcriptomic and lipidomic profiling of eicosanoid/docosanoid signalling in affected and non-affected skin of human atopic dermatitis patients. Exp. Dermatol. 2019, 28, 177–189. [Google Scholar] [CrossRef]
- Łuczaj, W.; Gęgotek, A.; Skrzydlewska, E. Analytical approaches to assess metabolic changes in psoriasis. J. Pharm. Biomed. Anal. 2021, 205, 114359. [Google Scholar] [CrossRef] [PubMed]
- Fyhrquist, N.; Muirhead, G.; Prast-Nielsen, S.; Jeanmougin, M.; Olah, P.; Skoog, T.; Jules-Clement, G.; Feld, M.; Barrientos-Somarribas, M.; Sinkko, H.; et al. Microbe-host interplay in atopic dermatitis and psoriasis. Nat. Commun. 2019, 10, 4703. [Google Scholar] [CrossRef] [PubMed]
- Hu, X.; Qi, C.; Feng, F.; Wang, Y.; Di, T.; Meng, Y.; Wang, Y.; Zhao, N.; Zhang, X.; Li, P.; et al. Combining network pharmacology, RNA-seq, and metabolomics strategies to reveal the mechanism of Cimicifugae Rhizoma—Smilax glabra Roxb herb pair for the treatment of psoriasis. Phytomedicine 2022, 105, 154384. [Google Scholar] [CrossRef] [PubMed]
- Qiu, Q.; Deng, J.; Deng, H.; Yao, D.; Yan, Y.; Ye, S.; Shang, X.; Deng, Y.; Han, L.; Zheng, G.; et al. Association of the characteristics of the blood metabolome and gut microbiome with the outcome of methotrexate therapy in psoriasis. Front. Immunol. 2022, 13, 937539. [Google Scholar] [CrossRef] [PubMed]
- Yang, G.; Zhou, S.; He, H.; Shen, Z.; Liu, Y.; Hu, J.; Wang, J. Exploring the “gene-protein-metabolite” network of coronary heart disease with phlegm and blood stasis syndrome by integrated multi-omics strategy. Front. Immunol. 2022, 13, 1022627. [Google Scholar] [CrossRef]
- Zhao, Y.; Jhamb, D.; Shu, L.; Arneson, D.; Rajpal, D.K.; Yang, X. Multi-omics integration reveals molecular networks and regulators of psoriasis. BMC Syst. Biol. 2019, 13, 8. [Google Scholar] [CrossRef]
- Agamah, F.E.; Bayjanov, J.R.; Niehues, A.; Njoku, K.F.; Skelton, M.; Mazandu, G.K.; Ederveen, T.H.A.; Mulder, N.; Chimusa, E.R.; ‘t Hoen, P.A.C. Computational approaches for network-based integrative multi-omics analysis. Front. Mol. Biosci. 2022, 9, 967205. [Google Scholar] [CrossRef]
- Srivastava, P.; Talwar, M.; Yadav, A.; Choudhary, A.; Mohanty, S.; Bharti, S.; Narad, P.; Sengupta, A. VIRdb 2.0: Interactive analysis of comorbidity conditions associated with vitiligo pathogenesis using co-expression network-based approach. F1000Research 2020, 9, 1055. [Google Scholar] [CrossRef]
- Ding, J.; Blencowe, M.; Nghiem, T.; Ha, S.-M.; Chen, Y.-W.; Li, G.; Yang, X. Mergeomics 2.0: A web server for multi-omics data integration to elucidate disease networks and predict therapeutics. Nucleic Acids Res. 2021, 49, W375–W387. [Google Scholar] [CrossRef]
- Cisek, K.; Krochmal, M.; Klein, J.; Mischak, H. The application of multi-omics and systems biology to identify therapeutic targets in chronic kidney disease. Nephrol. Dial. Transplant. 2016, 31, 2003–2011. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Database Name | Database Full Name | Application | Ref. |
|---|---|---|---|
| GEO | Gene Expression Omnibus repository of the National Library of Medicine | All | [17] |
| HGMD | Human Gene Mutation Database | Genomics | [18] |
| Ensembl | Ensembl | Genomics | [19] |
| ClinVar | ClinVar | Genomics | [20] |
| eFORGE | the epigenetic equivalent of FORGE, using EWAS rather than GWAS data | Epigenomics | [21] |
| HMDD | the Human microRNA Disease Database | MicroRNA | [22] |
| circRNADisease | circRNADisease | CircRNA | [23] |
| lncRNADisease | lncRNADisease | lncRNA | [24] |
| Uniprot | Uniprot | Proteomics | [25] |
| SMART | Simple Modular Architecture Research Tool | Proteomics | [26] |
| PTMD | Post-Translational Modification Database | Proteomics | [27] |
| HMDB | The Human Metabolome Database | Metabiomics | [28] |
| PHI | Pathogen–Host Interactions Database | Microbiomics | [29] |
| HMP | Human Microbiome Project Data Portal | Microbiomics | [30] |
| Pathways | Marker Name | Omics Type | Disease | Ref. |
|---|---|---|---|---|
| Epidermal differentiation pathway | S100A8(SNP) | genomics | Psoriasis | [36] |
| S100A8 | transcriptomics | Psoriasis | [37] | |
| S100A8 | transcriptomics | AD | [38] | |
| S100A8(protein) | proteomics | AD | [39] | |
| S100A8(protein) | proteomics | Psoriasis | [40] | |
| IL-17-mediated inflammatory Pathway | IL-1a(SNP) | genomics | AD | [41] |
| IL-1a | transcriptomics | Psoriasis | [42] | |
| IL-31 Pathway | IL-31 | transcriptomics | Psoriasis | [43] |
| IL-31 | transcriptomics | AD | [44] | |
| Calcium-permeable cation TRPs channels | TRPV1(SNP) | genomics | AD | [45] |
| TRPV1 | transcriptomics | Psoriasis | [46] | |
| TRPV1 | transcriptomics | AD | [44] | |
| TH1 cytokine | IL-13(SNP) | genomics | AD | [46] |
| IL-13 | genomics | Psoriasis | [47] | |
| IL-13 | transcriptomics | Psoriasis | [43] | |
| IL-13 | transcriptomics | AD | [48] | |
| IL-13 | transcriptomics | AD | [49] | |
| IL-13(protein) | proteomics | AD | [50] | |
| IL-20 family cytokine | IL-24(haplotype) | genomics | Psoriasis | [51] |
| IL-24 | transcriptomics | Psoriasis | [43] | |
| IL-24 | transcriptomics | AD | [52] | |
| Immunomodulatory chemokines | CXCL1 | transcriptomics | Psoriasis | [42] |
| CXCL1 | transcriptomics | AD | [38] | |
| CXCL1 | proteomics | Psoriasis | [53] | |
| CXCL1 | proteomics | AD | [54] | |
| IL-1 pathways | IL-36(SNP) | genomics | psoriasis | [55] |
| IL-36 | transcriptomics | Psoriasis | [56] | |
| IL-36 | transcriptomics | AD | [57] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Guo, Y.; Luo, L.; Zhu, J.; Li, C. Multi-Omics Research Strategies for Psoriasis and Atopic Dermatitis. Int. J. Mol. Sci. 2023, 24, 8018. https://doi.org/10.3390/ijms24098018
Guo Y, Luo L, Zhu J, Li C. Multi-Omics Research Strategies for Psoriasis and Atopic Dermatitis. International Journal of Molecular Sciences. 2023; 24(9):8018. https://doi.org/10.3390/ijms24098018
Chicago/Turabian StyleGuo, Youming, Lingling Luo, Jing Zhu, and Chengrang Li. 2023. "Multi-Omics Research Strategies for Psoriasis and Atopic Dermatitis" International Journal of Molecular Sciences 24, no. 9: 8018. https://doi.org/10.3390/ijms24098018