
Proteomic and Metabolomic Changes in Psoriasis Preclinical and Clinical Aspects
 ,
,
Abstract
:1. Introduction
2. Rodents as a Model for Psoriasis
2.1. Spontaneous Mouse Models
2.2. Induced Modulation of the Skin Environment
2.3. Psoriatic Human Skin Xenograft Model
2.4. Transgenic Models
2.5. T-Cell Transfer Rodents Model
2.6. Genome Editing-Models Based on CRISPR/Cas9 Technology
3. Proteomic and Metabolomic in Biomarker Discovery

3.1. Post-Translational Modifications (PTMs) of Proteins
3.2. Biological Material Selection for Proteomics/Metabolomics Studies
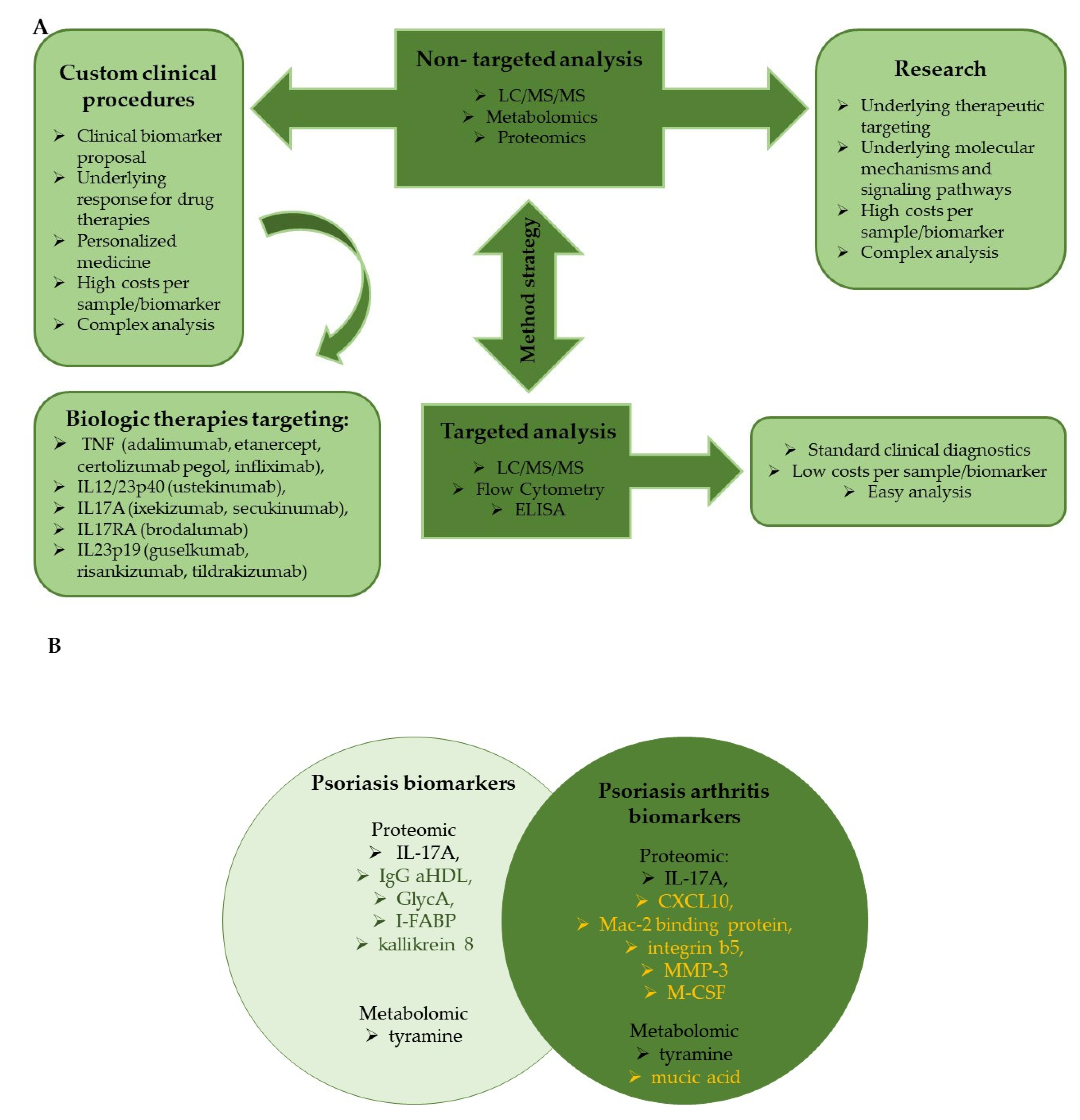
4. Clinical Trials in Proteomic and Metabolomic Biomarkers Discovery
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Yan, D.; Gudjonsson, J.E.; Le, S.; Maverakis, E.; Plazyo, O.; Ritchlin, C.; Scher, J.U.; Singh, R.; Ward, N.L.; Bell, S.; et al. New Frontiers in Psoriatic Disease Research, Part I: Genetics, Environmental Triggers, Immunology, Pathophysiology, and Precision Medicine. J. Investig. Dermatol. 2021, 141, 2112–2122.e3. [Google Scholar] [CrossRef] [PubMed]
- Parisi, R.; Iskandar, I.Y.K.; Kontopantelis, E.; Augustin, M.; Griffiths, C.E.M.; Ashcroft, D.M. National, Regional, and Worldwide Epidemiology of Psoriasis: Systematic Analysis and Modelling Study. BMJ 2020, 369, m1590. [Google Scholar] [CrossRef] [PubMed]
- Borzęcki, A.; Koncewicz, A.; Raszewska-Famielec, M.; Dudra-Jastrzębska, M. Epidemiology of Psoriasis in the Years 2008–2015 in Poland. Przegl. Dermatol. 2018, 105, 693–700. [Google Scholar] [CrossRef]
- Purzycka-Bohdan, D.; Kisielnicka, A.; Zabłotna, M.; Nedoszytko, B.; Nowicki, R.J.; Reich, A.; Samotij, D.; Szczęch, J.; Krasowska, D.; Bartosińska, J.; et al. Chronic Plaque Psoriasis in Poland: Disease Severity, Prevalence of Comorbidities, and Quality of Life. J. Clin. Med. 2022, 11, 1254. [Google Scholar] [CrossRef]
- Michalek, I.M.; Loring, B.; John, S.M. A Systematic Review of Worldwide Epidemiology of Psoriasis. J. Eur. Acad. Dermatol. Venereol. 2017, 31, 205–212. [Google Scholar] [CrossRef]
- AlQassimi, S.; AlBrashdi, S.; Galadari, H.; Hashim, M.J. Global Burden of Psoriasis—Comparison of Regional and Global Epidemiology, 1990 to 2017. Int. J. Dermatol. 2020, 59, 566–571. [Google Scholar] [CrossRef]
- Rendon, A.; Schäkel, K. Psoriasis Pathogenesis and Treatment. Int. J. Mol. Sci. 2019, 20, 1475. [Google Scholar] [CrossRef]
- Zachariae, H. Prevalence of Joint Disease in Patients with Psoriasis: Implications for Therapy. Am. J. Clin. Dermatol. 2003, 4, 441–447. [Google Scholar] [CrossRef]
- Bu, J.; Ding, R.; Zhou, L.; Chen, X.; Shen, E. Epidemiology of Psoriasis and Comorbid Diseases: A Narrative Review. Front. Immunol. 2022, 13, 880201. [Google Scholar] [CrossRef]
- Griffiths, C.E.M.; Armstrong, A.W.; Gudjonsson, J.E.; Barker, J.N.W.N. Psoriasis. Lancet 2021, 397, 1301–1315. [Google Scholar] [CrossRef]
- Sanchez, D.P.; Sonthalia, S. Koebner Phenomenon; StatPearls Publishing: Treasure Island, FL, USA, 2023. [Google Scholar]
- Rønholt, K.; Iversen, L. Old and New Biological Therapies for Psoriasis. Int. J. Mol. Sci. 2017, 18, 2297. [Google Scholar] [CrossRef]
- Gates, A.H.; Karasek, M. Hereditary Absence of Sebaceous Glands in the Mouse. Science 1965, 148, 1471–1473. [Google Scholar] [CrossRef]
- Sundberg, J.P.; Dunstan, R.W.; Roop, D.R.; Beamer, W.G. Full-Thickness Skin Grafts from Flaky Skin Mice to Nude Mice: Maintenance of the Psoriasiform Phenotype. J. Investig. Dermatol. 1994, 102, 781–788. [Google Scholar] [CrossRef]
- Sundberg, J.P.; Beamer, W.G.; Shultz, L.D.; Dunstan, R.W. Inherited Mouse Mutations as Models of Human Adnexal, Cornification, and Papulosquamous Dermatoses. J. Investig. Dermatol. 1990, 95, 62S–63S. [Google Scholar] [CrossRef]
- Sundberga, J.P.; Francea, M.; Boggessa, D.; Sundberga, B.A.; Jenson’, A.B.; Beamera, W.G.; Shultza, L.D.; Words, K. Development and Progression of Psoriasiform Dermatitis and Systemic Lesions in the Flaky Skin (Fsn) Mouse Mutant. Pathobiology 1997, 65, 271–286. [Google Scholar] [CrossRef]
- Brown, W.R.; Hardy, M.H. A Hypothesis on the Cause of Chronic Epidermal Hyperproliferation in Asebia Mice. Clin. Exp. Dermatol. 1988, 13, 74–77. [Google Scholar] [CrossRef]
- Fallon, P.G.; Sasaki, T.; Sandilands, A.; Campbell, L.E.; Saunders, S.P.; Mangan, N.E.; Callanan, J.J.; Kawasaki, H.; Shiohama, A.; Kubo, A.; et al. A Homozygous Frameshift Mutation in the Mouse Flg Gene Facilitates Enhanced Percutaneous Allergen Priming. Nat. Genet. 2009, 41, 602–608. [Google Scholar] [CrossRef]
- Wilkinson, D.I.; Karasek, M.A. Skin Lipids of a Normal and Mutant (Asebic) Mouse Strain. J. Investig. Dermatol. 1966, 47, 449–455. [Google Scholar] [CrossRef]
- Vandeghinste, N.; Klattig, J.; Jagerschmidt, C.; Lavazais, S.; Marsais, F.; Haas, J.D.; Auberval, M.; Lauffer, F.; Moran, T.; Ongenaert, M.; et al. Neutralization of IL-17C Reduces Skin Inflammation in Mouse Models of Psoriasis and Atopic Dermatitis. J. Investig. Dermatol. 2018, 138, 1555–1563. [Google Scholar] [CrossRef]
- Ho, A.W.; Gaffen, S.L. IL-17RC: A Partner in IL-17 Signaling and Beyond. Semin. Immunopathol. 2010, 32, 33–42. [Google Scholar] [CrossRef]
- Nies, J.F.; Panzer, U. IL-17C/IL-17RE: Emergence of a Unique Axis in TH17 Biology. Front. Immunol. 2020, 11, 341. [Google Scholar] [CrossRef] [PubMed]
- Chan, T.C.; Hawkes, J.E.; Krueger, J.G. Interleukin 23 in the Skin: Role in Psoriasis Pathogenesis and Selective Interleukin 23 Blockade as Treatment. Ther. Adv. Chronic Dis. 2018, 9, 111–119. [Google Scholar] [CrossRef] [PubMed]
- van der Fits, L.; Mourits, S.; Voerman, J.S.A.; Kant, M.; Boon, L.; Laman, J.D.; Cornelissen, F.; Mus, A.-M.; Florencia, E.; Prens, E.P.; et al. Imiquimod-Induced Psoriasis-Like Skin Inflammation in Mice Is Mediated via the IL-23/IL-17 Axis. J. Immunol. 2009, 182, 5836–5845. [Google Scholar] [CrossRef] [PubMed]
- Nakahara, T.; Kido-Nakahara, M.; Ulzii, D.; Miake, S.; Fujishima, K.; Sakai, S.; Chiba, T.; Tsuji, G.; Furue, M. Topical Application of Endothelin Receptor a Antagonist Attenuates Imiquimod-Induced Psoriasiform Skin Inflammation. Sci. Rep. 2020, 10, 9510. [Google Scholar] [CrossRef]
- Mohammed, S.S.; Kadhim, H.M.; Al-Sudani, I.M.; Musatafa, W.W. Anti-Inflammatory Effects of Topically Applied Azilsartan in a Mouse Model of Imiquimod-Induced Psoriasis. Int. J. Drug Deliv. Technol. 2022, 12, 1249–1255. [Google Scholar] [CrossRef]
- Yang, Y.; Zhao, Y.; Lai, R.; Xian, L.; Lei, Q.; Xu, J.; Guo, M.; Xian, D.; Zhong, J. An Emerging Role of Proanthocyanidins on Psoriasis: Evidence from a Psoriasis-Like Mouse Model. Oxid. Med. Cell. Longev. 2022, 2022, 5800586. [Google Scholar] [CrossRef]
- Schafer, P.H.; Chen, P.; Fang, L.; Wang, A.; Chopra, R. The Pharmacodynamic Impact of Apremilast, an Oral Phosphodiesterase 4 Inhibitor, on Circulating Levels of Inflammatory Biomarkers in Patients with Psoriatic Arthritis: Substudy Results from a Phase III, Randomized, Placebo-Controlled Trial (PALACE 1). J. Immunol. Res. 2015, 2015, 906349. [Google Scholar] [CrossRef]
- Zhou, Y.; Wang, P.; Yan, B.X.; Chen, X.Y.; Landeck, L.; Wang, Z.Y.; Li, X.X.; Zhang, J.; Zheng, M.; Man, X.Y. Quantitative Proteomic Profile of Psoriatic Epidermis Identifies OAS2 as a Novel Biomarker for Disease Activity. Front. Immunol. 2020, 11, 1432. [Google Scholar] [CrossRef]
- Bugaut, H.; Aractingi, S. Major Role of the IL17/23 Axis in Psoriasis Supports the Development of New Targeted Therapies. Front. Immunol. 2021, 12, 621956. [Google Scholar] [CrossRef]
- Bromley, S.K.; Larson, R.P.; Ziegler, S.F.; Luster, A.D. IL-23 Induces Atopic Dermatitis-Like Inflammation Instead of Psoriasis-Like Inflammation in CCR2-Deficient Mice. PLoS ONE 2013, 8, e58196. [Google Scholar] [CrossRef]
- Lindroos, J.; Svensson, L.; Norsgaard, H.; Lovato, P.; Moller, K.; Hagedorn, P.H.; Olsen, G.M.; Labuda, T. IL-23-Mediated Epidermal Hyperplasia Is Dependent on IL-6. J. Investig. Dermatol. 2011, 131, 1110–1118. [Google Scholar] [CrossRef]
- Chan, J.R.; Blumenschein, W.; Murphy, E.; Diveu, C.; Wiekowski, M.; Abbondanzo, S.; Lucian, L.; Geissler, R.; Brodie, S.; Kimball, A.B.; et al. IL-23 Stimulates Epidermal Hyperplasia via TNF and IL-20R2-Dependent Mechanisms with Implications for Psoriasis Pathogenesis. J. Exp. Med. 2006, 203, 2577–2587. [Google Scholar] [CrossRef]
- Austin, L.M.; Ozawa, M.; Kikuchi, T.; Walters, I.B.; Krueger, J.G. The Majority of Epidermal T Cells in Psoriasis Vulgaris Lesions Can Produce Type 1 Cytokines, Interferon-γ, Interleukin-2, and Tumor Necrosis Factor-α, Defining TC1 (Cytotoxic T Lymphocyte) and TH1 Effector Populations: 1 a Type 1 Differentiation Bias Is Also Measured in Circulating Blood T Cells in Psoriatic Patients. J. Investig. Dermatol. 1999, 113, 752–759. [Google Scholar]
- Svensson, L.; Røpke, M.A.; Norsgaard, H. Psoriasis Drug Discovery: Methods for Evaluation of Potential Drug Candidates. Expert Opin. Drug Discov. 2012, 7, 49–61. [Google Scholar] [CrossRef]
- Haftek, M.; Ortonne, J.P.; Staquet, M.J.; Viac, J.; Thivolet, J. Normal and Psoriatic Human Skin Grafts on “nude” Mice: Morphological and Immunochemical Studies. J. Investig. Dermatol. 1981, 76, 48–52. [Google Scholar] [CrossRef]
- Raychaudhuri, S.; Raychaudhuri, S. Scid Mouse Model of Psoriasis: A Unique Tool for Drug Development of Autoreactive T-Cell and TH-17 Cell-Mediated Autoimmune Diseases. Indian J. Dermatol. 2010, 55, 157. [Google Scholar] [CrossRef]
- Di Domizio, J.; Conrad, C.; Gilliet, M. Xenotransplantation Model of Psoriasis. In Inflammation: Methods in Molecular Biology; Clausen, B., Laman, J., Eds.; Humana Press: New York, NY, USA, 2017; Volume 1559, pp. 83–90. [Google Scholar] [CrossRef]
- Tiirikainen, M.L.; Woetmann, A.; Norsgaard, H.; Santamaria-Babí, L.F.; Lovato, P. Ex Vivo Culture of Lesional Psoriasis Skin for Pharmacological Testing. J. Dermatol. Sci. 2020, 97, 109–116. [Google Scholar] [CrossRef]
- Norsgaard, H.; Svensson, L.; Hagedorn, P.H.; Moller, K.; Olsen, G.M.; Labuda, T. Translating Clinical Activity and Gene Expression Signatures of Etanercept and Ciclosporin to the Psoriasis Xenograft SCID Mouse Model. Br. J. Dermatol. 2012, 166, 649–652. [Google Scholar] [CrossRef]
- Gourlay, W.A.; Chambers, W.H.; Monaco, A.P.; Maki, T. Importance of natural killer cells in the rejection of hamster skin xenografts. Transplantation 1998, 65, 727–734. [Google Scholar] [CrossRef]
- Ashkar, A.A.; Di Santo, J.P.; Croy, B.A. Interferon γ Contributes to Initiation of Uterine Vascular Modification, Decidual Integrity, and Uterine Natural Killer Cell Maturation during Normal Murine Pregnancy. J. Exp. Med. 2000, 192, 259–270. [Google Scholar] [CrossRef]
- Boyman, O.; Hefti, H.P.; Conrad, C.; Nickoloff, B.J.; Suter, M.; Nestle, F.O. Spontaneous Development of Psoriasis in a New Animal Model Shows an Essential Role for Resident T Cells and Tumor Necrosis Factor-α. J. Exp. Med. 2004, 199, 731–736. [Google Scholar] [CrossRef] [PubMed]
- Wagner, C.L.; Visvanathan, S.; Elashoff, M.; McInnes, I.B.; Mease, P.J.; Krueger, G.G.; Murphy, F.T.; Papp, K.; Gomez-Reino, J.J.; Mack, M.; et al. Markers of Inflammation and Bone Remodelling Associated with Improvement in Clinical Response Measures in Psoriatic Arthritis Patients Treated with Golimumab. Ann. Rheum. Dis. 2013, 72, 83–88. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Deshpande, M.; Grisotto, M.; Smaldini, P.; Garcia, R.; He, Z.; Gulko, P.S.; Lira, S.A.; Furtado, G.C. Skin Expression of IL-23 Drives the Development of Psoriasis and Psoriatic Arthritis in Mice. Sci. Rep. 2020, 10, 8259. [Google Scholar] [CrossRef] [PubMed]
- Van Nuffel, E.; Staal, J.; Baudelet, G.; Haegman, M.; Driege, Y.; Hochepied, T.; Afonina, I.S.; Beyaert, R. MALT 1 Targeting Suppresses CARD 14-induced Psoriatic Dermatitis in Mice. EMBO Rep. 2020, 21, e49237. [Google Scholar] [CrossRef]
- Schonthaler, H.B.; Huggenberger, R.; Wculek, S.K.; Detmar, M.; Wagner, E.F.; Karin, M. Systemic Anti-VEGF Treatment Strongly Reduces Skin Inflammation in a Mouse Model of Psoriasis. Proc. Natl. Acad. Sci. USA 2009, 106, 21264–21269. [Google Scholar] [CrossRef]
- Retser, E.; Schied, T.; Skryabin, B.V.; Vogl, T.; Kanczler, J.M.; Hamann, N.; Niehoff, A.; Hermann, S.; Eisenblätter, M.; Wachsmuth, L.; et al. Doxycycline-Induced Expression of Transgenic Human Tumor Necrosis Factor α in Adult Mice Results in Psoriasis-like Arthritis. Arthritis Rheum. 2013, 65, 2290–2300. [Google Scholar] [CrossRef]
- Voskas, D.; Jones, N.; Van Slyke, P.; Sturk, C.; Chang, W.; Haninec, A.; Olya Babichev, Y.; Tran, J.; Master, Z.; Chen, S.; et al. A Cyclosporine-Sensitive Psoriasis-Like Disease Produced in Tie2 Transgenic Mice. Am. J. Pathol. 2005, 166, 843–855. [Google Scholar] [CrossRef]
- Nakajima, K.; Kanda, T.; Takaishi, M.; Shiga, T.; Miyoshi, K.; Nakajima, H.; Kamijima, R.; Tarutani, M.; Benson, J.M.; Elloso, M.M.; et al. Distinct Roles of IL-23 and IL-17 in the Development of Psoriasis-Like Lesions in a Mouse Model. J. Immunol. 2011, 186, 4481–4489. [Google Scholar] [CrossRef]
- Rizzo, H.L.; Kagami, S.; Phillips, K.G.; Kurtz, S.E.; Jacques, S.L.; Blauvelt, A. IL-23–Mediated Psoriasis-Like Epidermal Hyperplasia Is Dependent on IL-17A. J. Immunol. 2011, 186, 1495–1502. [Google Scholar] [CrossRef]
- Detmar, M.; Brown, L.F.; Schön, M.P.; Elicker, B.M.; Velasco, P.; Richard, L.; Fukumura, D.; Monsky, W.; Claffey, K.P.; Jain, R.K. Increased Microvascular Density and Enhanced Leukocyte Rolling and Adhesion in the Skin of VEGF Transgenic Mice. J. Investig. Dermatol. 1998, 111, 1–6. [Google Scholar] [CrossRef]
- Blessing, M.; Schirmacher, P.; Kaiser, S. Overexpression of Bone Morphogenetic Protein-6 (BMP-6) in the Epidermis of Transgenic Mice: Inhibition or Stimulation of Proliferation Depending on the Pattern of Transgene Expression and Formation of Psoriatic Lesions. J. Cell Biol. 1996, 135, 227–239. [Google Scholar] [CrossRef]
- Billi, A.C.; Ludwig, J.E.; Fritz, Y.; Rozic, R.; Swindell, W.R.; Tsoi, L.C.; Gruzska, D.; Abdollahi-Roodsaz, S.; Xing, X.; Diaconu, D.; et al. KLK6 Expression in Skin Induces PAR1-Mediated Psoriasiform Dermatitis and Inflammatory Joint Disease. J. Clin. Investig. 2020, 130, 3151–3157. [Google Scholar] [CrossRef]
- Johnston, A.; Fritz, Y.; Dawes, S.M.; Diaconu, D.; Al-Attar, P.M.; Guzman, A.M.; Chen, C.S.; Fu, W.; Gudjonsson, J.E.; McCormick, T.S.; et al. Keratinocyte Overexpression of IL-17C Promotes Psoriasiform Skin Inflammation. J. Immunol. 2013, 190, 2252–2262. [Google Scholar] [CrossRef]
- SchÖn, P.; Detmar, M.; Parker’, C.M. Murine Psoriasis-like Disorder Induced by Naive CD4+ T Cells. Nat. Med. 1997, 3, 183–188. [Google Scholar] [CrossRef]
- Davenport, C.M.; Mcadams, H.A.; Kou, J.; Mascioli, K.; Eichman, C.; Healy, L.; Peterson, J.; Murphy, S.; Coppola, D.; Truneh, A. Inhibition of Pro-Inf Lammatory Cytokine Generation by CTLA4-Ig in the Skin and Colon of Mice Adoptively Transplanted with CD45RB Hi CD4 + T Cells Correlates with Suppression of Psoriasis and Colitis. Int. Immunopharmacol. 2002, 2, 653–672. [Google Scholar] [CrossRef]
- Takahashi, H.; Kouno, M.; Nagao, K.; Wada, N.; Hata, T.; Nishimoto, S.; Iwakura, Y.; Yoshimura, A.; Yamada, T.; Kuwana, M.; et al. Desmoglein 3-Specific CD4+ T Cells Induce Pemphigus Vulgaris and Interface Dermatitis in Mice. J. Clin. Investig. 2011, 121, 3677–3688. [Google Scholar] [CrossRef]
- Breban, M.; Fernández-Sueiro, J.L.; Richardson, J.A.; Hadavand, R.R.; Maika, S.D.; Hammer, R.E.; Taurog, J.D. T Cells, but Not Thymic Exposure to HLA-B27, Are Required for the Inflammatory Disease of HLA-B27 Transgenic Rats. J. Immunol. 1996, 156, 794–803. [Google Scholar] [CrossRef]
- Hong, K.; Chu, A.; Lú, B.R.; Berg, E.L.; Ehrhardt, R.O. IL-12, Independently of IFN-gamma, Plays a Crucial Role in the Pathogenesis of a Murine Psoriasis-Like Skin Disorder. J. Immunol. 1999, 162, 7480–7491. [Google Scholar] [CrossRef]
- Nishimoto, S.; Kotani, H.; Tsuruta, S.; Shimizu, N.; Ito, M.; Shichita, T.; Morita, R.; Takahashi, H.; Amagai, M.; Yoshimura, A. Th17 Cells Carrying TCR Recognizing Epidermal Autoantigen Induce Psoriasis-like Skin Inflammation. J. Immunol. 2013, 191, 3065–3072. [Google Scholar] [CrossRef]
- Dort, E.N.; Tanguay, P.; Hamelin, R.C. CRISPR/Cas9 Gene Editing: An Unexplored Frontier for Forest Pathology. Front. Plant Sci. 2020, 11, 1126. [Google Scholar] [CrossRef]
- Strzyz, P. CRISPR–Cas9 Wins Nobel. Nat. Rev. Mol. Cell Biol. 2020, 21, 714. [Google Scholar] [CrossRef] [PubMed]
- Roth-Carter, Q.R.; Godsel, L.; Koetsier, J.L.; Broussard, J.A.; Burks, H.E.; Fitz, G.; Huffine, A.L.; Amagai, S.; Lloyd, S.; Kweon, J.; et al. 225 Desmoglein 1 Deficiency in Knockout Mice Impairs Epidermal Barrier Formation and Results in a Psoriasis-like Gene Signature in E18.5 Embryos. J. Investig. Dermatol. 2020, 140, S26. [Google Scholar] [CrossRef]
- Godsel, L.M.; Roth-Carter, Q.R.; Koetsier, J.L.; Tsoi, L.C.; Broussard, J.A.; Fitz, G.N.; Lloyd, S.M.; Kweon, J.; Huffine, A.L.; Burks, H.E.; et al. Th17-Skewed Inflammation Due to Genetic Deficiency of a Cadherin Stress Sensor. bioRxiv 2020. [Google Scholar] [CrossRef]
- Ippagunta, S.K.; Gangwar, R.; Finkelstein, D.; Vogel, P.; Pelletier, S.; Gingras, S.; Redeckea, V.; Häckera, H. Keratinocytes Contribute Intrinsically to Psoriasis upon Loss of TNIP1 Function. Proc. Natl. Acad. Sci. USA 2016, 113, E6162–E6171. [Google Scholar] [CrossRef]
- Li, H.; Yao, Q.; Mariscal, A.G.; Wu, X.; Hülse, J.; Pedersen, E.; Helin, K.; Waisman, A.; Vinkel, C.; Thomsen, S.F.; et al. Epigenetic Control of IL-23 Expression in Keratinocytes Is Important for Chronic Skin Inflammation. Nat. Commun. 2018, 9, 1420. [Google Scholar] [CrossRef]
- Zomer, H.D.; Trentin, A.G. Skin Wound Healing in Humans and Mice: Challenges in Translational Research. J. Dermatol. Sci. 2018, 90, 3–12. [Google Scholar] [CrossRef]
- Gangwar, R.S.; Gudjonsson, J.E.; Ward, N.L. Mouse Models of Psoriasis: A Comprehensive Review. J. Investig. Dermatol. 2022, 142, 884–897. [Google Scholar] [CrossRef]
- Smajlović, A.; Haverić, A.; Alić, A.; Hadžić, M.; Smajlović, A.; Mujezinović, I.; Lojo-Kadrić, N.; Ramić, J.; Elez-Burnjaković, N.; Haverić, S.; et al. Molecular and Histopathological Profiling of Imiquimod Induced Dermatosis in Swiss Wistar Rats: Contribution to the Rat Model for Novel Anti-Psoriasis Treatments. Mol. Biol. Rep. 2021, 48, 4295–4303. [Google Scholar] [CrossRef]
- Liu, N.; Qin, H.; Cai, Y.; Li, X.; Wang, L.; Xu, Q.; Xue, F.; Chen, L.; Ding, C.; Hu, X.; et al. Dynamic Trafficking Patterns of IL-17-Producing Γδ T Cells Are Linked to the Recurrence of Skin Inflammation in Psoriasis-like Dermatitis. EBioMedicine 2022, 82, 104136. [Google Scholar] [CrossRef]
- Christensen, P.K.F.; Hansen, A.K.; Skov, S.; Engkilde, K.; Larsen, J.; Høyer-Hansen, M.H.; Koch, J. Sustaining the T-Cell Activity in Xenografted Psoriasis Skin. PLoS ONE 2023, 18, e0278390. [Google Scholar] [CrossRef]
- Kowalczyk, T.; Ciborowski, M.; Kisluk, J.; Kretowski, A.; Barbas, C. Mass Spectrometry Based Proteomics and Metabolomics in Personalized Oncology. Biochim. Biophys. Acta (BBA)—Mol. Basis Dis. 2020, 1866, 165690. [Google Scholar] [CrossRef]
- Rozanova, S.; Barkovits, K.; Nikolov, M.; Schmidt, C.; Urlaub, H.; Marcus, K. Quantitative Mass Spectrometry-Based Proteomics: An Overview. In Methods in Molecular Biology; Humana: New York, NY, USA, 2021; Volume 2228, pp. 85–116. [Google Scholar] [CrossRef]
- Schonthaler, H.B.; Guinea-Viniegra, J.; Wculek, S.K.; Ruppen, I.; Ximénez-Embún, P.; Guío-Carrión, A.; Navarro, R.; Hogg, N.; Ashman, K.; Wagner, E.F. S100A8-S100A9 Protein Complex Mediates Psoriasis by Regulating the Expression of Complement Factor C3. Immunity 2013, 39, 1171–1181. [Google Scholar] [CrossRef]
- Yan, K.X.; Meng, Q.; He, H.; Zhu, H.W.; Wang, Z.C.; Han, L.; Huang, Q.; Zhang, Z.H.; Yawalkar, N.; Zhou, H.; et al. ITRAQ-Based Quantitative Proteomics Reveals Biomarkers/Pathways in Psoriasis That Can Predict the Efficacy of Methotrexate. J. Eur. Acad. Dermatol. Venereol. 2022, 36, 1784–1795. [Google Scholar] [CrossRef]
- Li, Y.; Lin, P.; Wang, S.; Li, S.; Wang, R.; Yang, L.; Wang, H. Quantitative Analysis of Differentially Expressed Proteins in Psoriasis Vulgaris Using Tandem Mass Tags and Parallel Reaction Monitoring. Clin. Proteom. 2020, 17, 30. [Google Scholar] [CrossRef]
- Gęgotek, A.; Domingues, P.; Wroński, A.; Wójcik, P.; Skrzydlewska, E. Proteomic Plasma Profile of Psoriatic Patients. J. Pharm. Biomed. Anal. 2018, 155, 185–193. [Google Scholar] [CrossRef]
- Gęgotek, A.; Domingues, P.; Wroński, A.; Ambrożewicz, E.; Skrzydlewska, E. The Proteomic Profile of Keratinocytes and Lymphocytes in Psoriatic Patients. Proteom. Clin. Appl. 2019, 13, 1800119. [Google Scholar] [CrossRef]
- Gęgotek, A.; Domingues, P.; Wroński, A.; Skrzydlewska, E. Changes in Proteome of Fibroblasts Isolated from Psoriatic Skin Lesions. Int. J. Mol. Sci. 2020, 21, 5363. [Google Scholar] [CrossRef]
- Xu, M.; Deng, J.; Xu, K.; Zhu, T.; Han, L.; Yan, Y.; Yao, D.; Deng, H.; Wang, D.; Sun, Y.; et al. In-Depth Serum Proteomics Reveals Biomarkers of Psoriasis Severity and Response to Traditional Chinese Medicine. Theranostics 2019, 9, 2475–2488. [Google Scholar] [CrossRef]
- Reindl, J.; Pesek, J.; Krüger, T.; Wendler, S.; Nemitz, S.; Muckova, P.; Büchler, R.; Opitz, S.; Krieg, N.; Norgauer, J.; et al. Proteomic Biomarkers for Psoriasis and Psoriasis Arthritis. J. Proteom. 2016, 140, 55–61. [Google Scholar] [CrossRef]
- Plavina, T.; Hincapie, M.; Wakshull, E.; Subramanyam, M.; Hancock, S.W. Increased Plasma Concentrations of Cytoskeletal and Aa2+-Binding Proteins and Their Peptides in Psoriasis Patients. Clin. Chem. 2008, 54, 1805–1814. [Google Scholar] [CrossRef]
- Matsuura, T.; Sato, M.; Nagai, K.; Sato, T.; Arito, M.; Omoteyama, K.; Suematsu, N.; Okamoto, K.; Kato, T.; Soma, Y.; et al. Serum Peptides as Putative Modulators of Inflammation in Psoriasis. J. Dermatol. Sci. 2017, 87, 36–49. [Google Scholar] [CrossRef] [PubMed]
- Łuczaj, W.; Gęgotek, A.; Skrzydlewska, E. Analytical Approaches to Assess Metabolic Changes in Psoriasis. J. Pharm. Biomed. Anal. 2021, 205, 114359. [Google Scholar] [CrossRef] [PubMed]
- Eissa, A.; Cretu, D.; Soosaipillai, A.; Thavaneswaran, A.; Pellett, F.; Diamandis, A.; Cevikbas, F.; Steinhoff, M.; Diamandis, E.P.; Gladman, D.; et al. Serum Kallikrein-8 Correlates with Skin Activity, but Not Psoriatic Arthritis, in Patients with Psoriatic Disease. Clin. Chem. Lab. Med. 2013, 51, 317–325. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Xu, Q.; Li, B.; Li, H.; Shen, S.; Wu, J.; Ge, H.; Zhang, H.; Chen, S.; Chen, W.; et al. Proteomic Analysis of Psoriatic Skin Lesions in a Chinese Population. J. Proteom. 2021, 240, 104207. [Google Scholar] [CrossRef]
- Li, S.S.; Liu, Y.; Li, H.; Wang, L.-P.; Xue, L.-F.; Yin, G.-S.; Wu, X.-S. Identification of Psoriasis Vulgaris Biomarkers in Human Plasma by Non-Targeted Metabolomics Based on UPLC-Q-TOF/MS. Eur. Rev. Med. Pharmacol. Sci. 2019, 23, 3940–3950. [Google Scholar]
- Kishikawa, T.; Arase, N.; Tsuji, S.; Maeda, Y.; Nii, T.; Hirata, J.; Suzuki, K.; Yamamoto, K.; Masuda, T.; Ogawa, K.; et al. Large-Scale Plasma-Metabolome Analysis Identifies Potential Biomarkers of Psoriasis and Its Clinical Subtypes. J. Dermatol. Sci. 2021, 102, 78–84. [Google Scholar] [CrossRef]
- Sun, L.; Guo, X.; Qin, Y.; Li, P.; Yu, C.; Gao, X.; Xie, X.; Xu, X. Serum Intestinal Metabolites Are Raised in Patients with Psoriasis and Metabolic Syndrome. Clin. Cosmet. Investig. Dermatol. 2022, 15, 879–886. [Google Scholar] [CrossRef]
- Armstrong, A.W.; Wu, J.; Johnson, M.A.; Grapov, D.; Azizi, B.; Dhillon, J.; Fiehn, O. Metabolomics in Psoriatic Disease: Pilot Study Reveals Metabolite Differences in Psoriasis and Psoriatic Arthritis. F1000Research 2014, 3, 248. [Google Scholar] [CrossRef]
- Kang, H.; Li, X.; Zhou, Q.; Quan, C.; Xue, F.; Zheng, J.; Yu, Y. Exploration of Candidate Biomarkers for Human Psoriasis Based on Gas Chromatography-Mass Spectrometry Serum Metabolomics. Br. J. Dermatol. 2017, 176, 713–722. [Google Scholar] [CrossRef]
- Looby, N.; Roszkowska, A.; Reyes-Garcés, N.; Yu, M.; Bączek, T.; Kulasingam, V.; Pawliszyn, J.; Chandran, V. Serum Metabolic Fingerprinting of Psoriasis and Psoriatic Arthritis Patients Using Solid-Phase Microextraction—Liquid Chromatography—High-Resolution Mass Spectrometry. Metabolomics 2021, 17, 59. [Google Scholar] [CrossRef]
- Mysliwiec, H.; Harasim-Symbor, E.; Baran, A.; Szterling-Jaworowska, M.; Milewska, A.J.; Chabowski, A.; Flisiak, I. Abnormal Serum Fatty Acid Profile in Psoriatic Arthritis. Arch. Med. Sci. 2019, 15, 1407–1414. [Google Scholar] [CrossRef]
- Ambrożewicz, E.; Wójcik, P.; Wroński, A.; Łuczaj, W.; Jastrzab, A.; Žarković, N.; Skrzydlewska, E. Pathophysiological Alterations of Redox Signaling and Endocannabinoid System in Granulocytes and Plasma of Psoriatic Patients. Cells 2018, 7, 159. [Google Scholar] [CrossRef]
- Tsoukalas, D.; Fragoulakis, V.; Sarandi, E.; Docea, A.O.; Papakonstaninou, E.; Tsilimidos, G.; Anamaterou, C.; Fragkiadaki, P.; Aschner, M.; Tsatsakis, A.; et al. Targeted Metabolomic Analysis of Serum Fatty Acids for the Prediction of Autoimmune Diseases. Front. Mol. Biosci. 2019, 6, 120. [Google Scholar] [CrossRef]
- Bilgiç, Ö.; Altınyazar, H.C.; Baran, H.; Ünlü, A. Serum Homocysteine, Asymmetric Dimethyl Arginine (ADMA) and Other Arginine–NO Pathway Metabolite Levels in Patients with Psoriasis. Arch. Dermatol. Res. 2015, 307, 439–444. [Google Scholar] [CrossRef]
- Sikora, M.; Kiss, N.; Stec, A.; Giebultowicz, J.; Samborowska, E.; Jazwiec, R.; Dadlez, M.; Olszewska, M.; Rudnicka, L. Trimethylamine N-Oxide, a Gut Microbiota-Derived Metabolite, Is Associated with Cardiovascular Risk in Psoriasis: A Cross-Sectional Pilot Study. Dermatol. Ther. 2021, 11, 1277–1289. [Google Scholar] [CrossRef]
- Chen, C.; Hou, G.; Zeng, C.; Ren, Y.; Chen, X.; Peng, C. Metabolomic Profiling Reveals Amino Acid and Carnitine Alterations as Metabolic Signatures in Psoriasis. Theranostics 2020, 11, 754–767. [Google Scholar] [CrossRef]
- Ramessur, R.; Corbett, M.; Marshall, D.; Acencio, M.L.; Barbosa, I.A.; Dand, N.; Di Meglio, P.; Haddad, S.; Jensen, A.H.M.; Koopmann, W.; et al. Biomarkers of Disease Progression in People with Psoriasis: A Scoping Review. Br. J. Dermatol. 2022, 187, 481–493. [Google Scholar] [CrossRef]
- Ziehfreund, S.; Tizek, L.; Hangel, N.; Fritzsche, M.-C.; Weidinger, S.; Smith, C.; Bryce, P.J.; Greco, D.; Van Den Bogaard, E.H.; Flohr, C.; et al. Requirements and Expectations of High-Quality Biomarkers for Atopic Dermatitis and Psoriasis in 2021—A Two-Round Delphi Survey among International Experts. J. Eur. Acad. Dermatol. Venereol. 2022, 36, 1467–1476. [Google Scholar] [CrossRef]
- Shelef, M.A.; Sokolove, J.; Lahey, L.J.; Wagner, C.A.; Sackmann, E.K.; Warner, T.F.; Wang, Y.; Beebe, D.J.; Robinson, W.H.; Huttenlocher, A. Peptidylarginine Deiminase 4 Contributes to Tumor Necrosis Factor α-Induced Inflammatory Arthritis. Arthritis Rheumatol. 2014, 66, 1482–1491. [Google Scholar] [CrossRef]
- Kiss, B.; Szántó, M.; Hegedűs, C.; Antal, D.; Szödényi, A.; Márton, J.; Méhes, G.; Virág, L.; Szegedi, A.; Bai, P. Poly(ADP-Ribose) Polymerase-1 Depletion Enhances the Severity of Inflammation in an Imiquimod-Induced Model of Psoriasis. Exp. Dermatol. 2020, 29, 79–85. [Google Scholar] [CrossRef]
- Sestito, R.; Madonna, S.; Scarponi, C.; Cianfarani, F.; Failla, C.M.; Cavani, A.; Girolomoni, G.; Albanesi, C. STAT3-dependent Effects of IL-22 in Human Keratinocytes Are Counterregulated by Sirtuin 1 through a Direct Inhibition of STAT3 Acetylation. FASEB J. 2011, 25, 916–927. [Google Scholar] [CrossRef] [PubMed]
- Fan, X.; Yan, K.; Meng, Q.; Sun, R.; Yang, X.; Yuan, D.; Li, F.; Deng, H. Abnormal Expression of SIRTs in Psoriasis: Decreased Expression of SIRT 1-5 and Increased Expression of SIRT 6 and 7. Int. J. Mol. Med. 2019, 44, 157–171. [Google Scholar] [CrossRef] [PubMed]
- Bohio, A.A.; Sattout, A.; Wang, R.; Wang, K.; Sah, R.K.; Guo, X.; Zeng, X.; Ke, Y.; Boldogh, I.; Ba, X. C-Abl–Mediated Tyrosine Phosphorylation of PARP1 Is Crucial for Expression of Proinflammatory Genes. J. Immunol. 2019, 203, 1521–1531. [Google Scholar] [CrossRef] [PubMed]
- Kelel, M.; Yang, R.B.; Tsai, T.F.; Liang, P.H.; Wu, F.Y.; Huang, Y.T.; Yang, M.F.; Hsiao, Y.P.; Wang, L.F.; Tu, C.F.; et al. FUT8 Remodeling of EGFR Regulates Epidermal Keratinocyte Proliferation during Psoriasis Development. J. Investig. Dermatol. 2021, 141, 512–522. [Google Scholar] [CrossRef]
- Zou, C.; Huang, C.; Yan, L.; Li, X.; Xing, M.; Li, B.; Gao, C.; Wang, H. Serum N-Glycan Profiling as a Diagnostic Biomarker for the Identification and Assessment of Psoriasis. J. Clin. Lab. Anal. 2021, 35, e23711. [Google Scholar] [CrossRef]
- Ge, H.; Li, B.; Chen, W.; Xu, Q.; Chen, S.; Zhang, H.; Wu, J.; Zhen, Q.; Li, Y.; Yong, L.; et al. Differential Occurrence of Lysine 2-Hydroxyisobutyrylation in Psoriasis Skin Lesions. J. Proteom. 2019, 205, 103420. [Google Scholar] [CrossRef]
- Zhou, B.; Yang, W.; Li, W.; He, L.; Lu, L.; Zhang, L.; Liu, Z.; Wang, Y.; Chao, T.; Huang, R.; et al. Zdhhc2 Is Essential for Plasmacytoid Dendritic Cells Mediated Inflammatory Response in Psoriasis. Front. Immunol. 2021, 11, 607442. [Google Scholar] [CrossRef]
- Yang, L.; Yan, Y. Emerging Roles of Post-Translational Modifications in Skin Diseases: Current Knowledge, Challenges and Future Perspectives. J. Inflamm. Res. 2022, 15, 965–975. [Google Scholar] [CrossRef]
- Lundberg, K.C.; Fritz, Y.; Johnston, A.; Foster, A.M.; Baliwag, J.; Gudjonsson, J.E.; Schlatzer, D.; Gokulrangan, G.; McCormick, T.S.; Chance, M.R.; et al. Proteomics of Skin Proteins in Psoriasis: From Discovery and Verification in a Mouse Model to Confirmation in Humans. Mol. Cell. Proteom. 2015, 14, 109–119. [Google Scholar] [CrossRef]
- Swindell, W.R.; Remmer, H.A.; Sarkar, M.K.; Xing, X.; Barnes, D.H.; Wolterink, L.; Voorhees, J.J.; Nair, R.P.; Johnston, A.; Elder, J.T.; et al. Proteogenomic Analysis of Psoriasis Reveals Discordant and Concordant Changes in MRNA and Protein Abundance. Genome Med. 2015, 7, 86. [Google Scholar] [CrossRef]
- Ryu, J.; Park, S.G.; Park, B.C.; Choe, M.; Lee, K.S.; Cho, J.W. Proteomic Analysis of Psoriatic Skin Tissue for Identification of Differentially Expressed Proteins: Up-Regulation of GSTP1, SFN and PRDX2 in Psoriatic Skin. Int. J. Mol. Med. 2011, 28, 785–792. [Google Scholar] [CrossRef]
- Piruzian, E.; Bruskin, S.; Ishkin, A.; Abdeev, R.; Moshkovskii, S.; Melnik, S.; Nikolsky, Y.; Nikolskaya, T. Integrated Network Analysis of Transcriptomic and Proteomic Data in Psoriasis. BMC Syst. Biol. 2010, 4, 41. [Google Scholar] [CrossRef]
- Méhul, B.; Ménigot, C.; Fogel, P.; Seraidaris, A.; Genette, A.; Pascual, T.; Duvic, M.; Voegel, J.J. Proteomic Analysis of Stratum Corneum in Cutaneous T-Cell Lymphomas and Psoriasis. Exp. Dermatol. 2019, 28, 317–321. [Google Scholar] [CrossRef]
- Méhul, B.; Laffet, G.; Séraïdaris, A.; Russo, L.; Fogel, P.; Carlavan, I.; Pernin, C.; Andres, P.; Queille-Roussel, C.; Voegel, J.J. Noninvasive Proteome Analysis of Psoriatic Stratum Corneum Reflects Pathophysiological Pathways and Is Useful for Drug Profiling. Br. J. Dermatol. 2017, 177, 470–488. [Google Scholar] [CrossRef]
- Kapoor, S.R.; Filer, A.; Fitzpatrick, M.A.; Fisher, B.A.; Taylor, P.C.; Buckley, C.D.; McInnes, I.B.; Raza, K.; Young, S.P. Metabolic Profiling Predicts Response to Anti-Tumor Necrosis Factor α Therapy in Patients with Rheumatoid Arthritis. Arthritis Rheum. 2013, 65, 1448–1456. [Google Scholar] [CrossRef]
- Julià, A.; Vinaixa, M.; Domènech, E.; Fernández-Nebro, A.; Cañete, J.D.; Ferrándiz, C.; Tornero, J.; Gisbert, J.P.; Nos, P.; Casbas, A.G.; et al. Urine Metabolome Profiling of Immune-Mediated Inflammatory Diseases. BMC Med. 2016, 14, 133. [Google Scholar] [CrossRef]
- van Swelm, R.P.L.; Laarakkers, C.M.M.; Kooijmans-Otero, M.; de Jong, E.M.G.J.; Masereeuw, R.; Russel, F.G.M. Biomarkers for Methotrexate-Induced Liver Injury: Urinary Protein Profiling of Psoriasis Patients. Toxicol. Lett. 2013, 221, 219–224. [Google Scholar] [CrossRef]
- Owczarczyk-Saczonek, A.; Purzycka-Bohdan, D.; Nedoszytko, B.; Reich, A.; Szczerkowska-Dobosz, A.; Bartosinska, J.; Batycka-Baran, A.; Czajkowski, R.; Dobrucki, I.T.; Dobrucki, L.W.; et al. Pathogenesis of Psoriasis in the “Omic” Era. Part III. Metabolic Disorders, Metabolomics, Nutrigenomics in Psoriasis. Postepy Dermatol. Alergol. 2020, 37, 452–467. [Google Scholar] [CrossRef]
- Boehncke, W.H. Systemic Inflammation and Cardiovascular Comorbidity in Psoriasis Patients: Causes and Consequences. Front. Immunol. 2018, 9, 579. [Google Scholar] [CrossRef]
- Egeberg, A.; Skov, L.; Gislason, G.; Thyssen, J.; Mallbris, L. Incidence and Prevalence of Psoriasis in Denmark. Acta Dermato-Venereol. 2017, 97, 808–812. [Google Scholar] [CrossRef]
- Alidrisi, H.A.; Al Hamdi, K.; Mansour, A.A. Is There Any Association Between Psoriasis and Hashimoto’s Thyroiditis? Cureus 2019, 11, e4269. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
| Model Type | Animal | Most Common | Mechanism of Psoriasis Development | Advantages | Disadvantages | Ref. |
|---|---|---|---|---|---|---|
| Induced modulation of skin environment |
|
| Activation of:
|
|
| [24,26,70,71] |
| Stimulation of production IL-19 and IL-24 |
|
| [23,32] | ||
| Xenograft model |
|
| Xenograft possesses human immune cells which affect recipient and can induce skin lesions. |
|
| [36,38,40,44,72] |
| T-cell transfer |
|
| Shifting the balance of immune cells |
|
| [56,59,60,61] |
| Spontaneous mutation |
|
| mechanism dependent on the specific gene(s) |
|
| [13,14,15,16,19] |
| Transgenic models |
| Gene overexpression of, e.g., TGFα, IL-6, INFγ, VEGF, IL-23, MEK1; KLK6+ Gene inactivation: CD18−/−, IL-1RA−/−, integrin αE−/−, and many others | mechanism dependent on the down/upregulation of the specific gene(s) |
|
| [54] |
| Genome editing |
| CRISPR/Cas9 technology | mechanism dependent on the down/upregulation of the specific gene(s) |
|
| [62,64,67] |
| Sample | Skin | Plasma/Serum | Urine |
|---|---|---|---|
Easy to:
| + +++ +++ | ++ +++ +++ | +++ +++ +++ |
| Non-invasive | + | ++ | +++ |
| Reproducible | + | +++ | ++ |
| Complexity | ++ | +++ | + |
| Preparation | ++ | +++ | ++ |
| Metabolomics | ++ | +++ | + |
| Proteomics | ++ | +++ | + |
| Contamination | hair cosmetic/drugs residues | hemolyzed lipemic | proteins particles |
| Other | Different method for sample collection: biopsy, blister fluid, scraping, epidermis, stratum corneum | - | - |
| Clinical Trial/ NCT Number | Localization | Number of Participants | End-of-Clinical Trial | Biomarkers Examined | Method |
|---|---|---|---|---|---|
| Psoriatic Inflammation Markers Predictive of Response to Adalimumab (IMPRA)/ NCT03389984 | France | 85 | 2019 | mRNA expression in skin; Drug response: Adalimumab | RT-PCR |
| Multiple-dose Regimen Study to Assess Effect of 12 Months of Secukinumab Treatment on Skin Response and Biomarkers in Psoriasis Patients/ NCT01537432 | United States | 36 | 2014 | histological disease reversal score in skin biopsies; Drug response: Secukinumab | histological examination |
| Monocyte Biomarkers in Moderate to Severe Plaque Psoriasis Subjects Treated With Apremilast/ NCT03442088 | United States | 28 | 2018 | Serum markers of activated monocytes, monocyte transcriptome biomarkers, serum MPO, resistin, IL-17, and TF; Drug response: Apremilast | flow cytometry, qPCR |
| Psoriasis Inflammation and Systemic Co Morbidities/ NCT01170715 | United States | 29 | 2013 | IL17A gene expression, inflammatory cytokines, metabolic and vascular markers; Drug response: Etanercept | ECL, RT-PCR, histological examination |
| Identification of New Prognostic Markers in Psoriatic Arthritis/ NCT03455166 | Italy | 100 | 2016 | Serum levels of cytokine referable to Th17 pathway, MMPs, TIMPs, and markers of bone remodeling. | Data not provided |
| An Explorative Psoriasis Biomarker Study/ NCT04394936 | Netherlands | 50 | 2022 | Lipidomic of the stratum corneum, cutaneous and fecal microbiome, immune cell subsets and protein biomarkers, and genotyping Drug response: Guselkumab | LC-MS, ELISA, NGS, flow cytometry |
| Metabolic Biomarkers in Hashimoto’s Thyroiditis and Psoriasis/ NCT04693936 | Greece | 200 | 2024 | Urinary organic acids levels, blood fatty acids | GC-MS |
| Identification of New Biomarkers to Promote Personalized Treatment of Patients With Inflammatory Rheumatic Diseases/ NCT03214263 | Denmark | 20,000 | 2024 | Data not provided | Data not provided |
| Molecular Signatures in Inflammatory Skin Disease (MSID)/ NCT03358693 | Germany | 300 | 2028 | Immune cell composition, transcriptome, proteome, and microbiome signatures | DNA/RNA sequencing, ELISA, MS, flow cytometry |
| Metabolic Profiling of Immune Responses in Immune-mediated diseases/ NCT04864886 | United States | 150 | 2027 | Metabolomics of immune-mediated diseases in peripheral blood cells, serum, skin biopsies, skin tape strips, and skin swabs, metabolomics of the microbiome | RNA sequencing, MS |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Radulska, A.; Pelikant-Małecka, I.; Jendernalik, K.; Dobrucki, I.T.; Kalinowski, L. Proteomic and Metabolomic Changes in Psoriasis Preclinical and Clinical Aspects. Int. J. Mol. Sci. 2023, 24, 9507. https://doi.org/10.3390/ijms24119507
Radulska A, Pelikant-Małecka I, Jendernalik K, Dobrucki IT, Kalinowski L. Proteomic and Metabolomic Changes in Psoriasis Preclinical and Clinical Aspects. International Journal of Molecular Sciences. 2023; 24(11):9507. https://doi.org/10.3390/ijms24119507
Chicago/Turabian StyleRadulska, Adrianna, Iwona Pelikant-Małecka, Kamila Jendernalik, Iwona T. Dobrucki, and Leszek Kalinowski. 2023. "Proteomic and Metabolomic Changes in Psoriasis Preclinical and Clinical Aspects" International Journal of Molecular Sciences 24, no. 11: 9507. https://doi.org/10.3390/ijms24119507