
Disruptions of Circadian Genes in Cutaneous Melanoma—An In Silico Analysis of Transcriptome Databases


Abstract
:1. Introduction
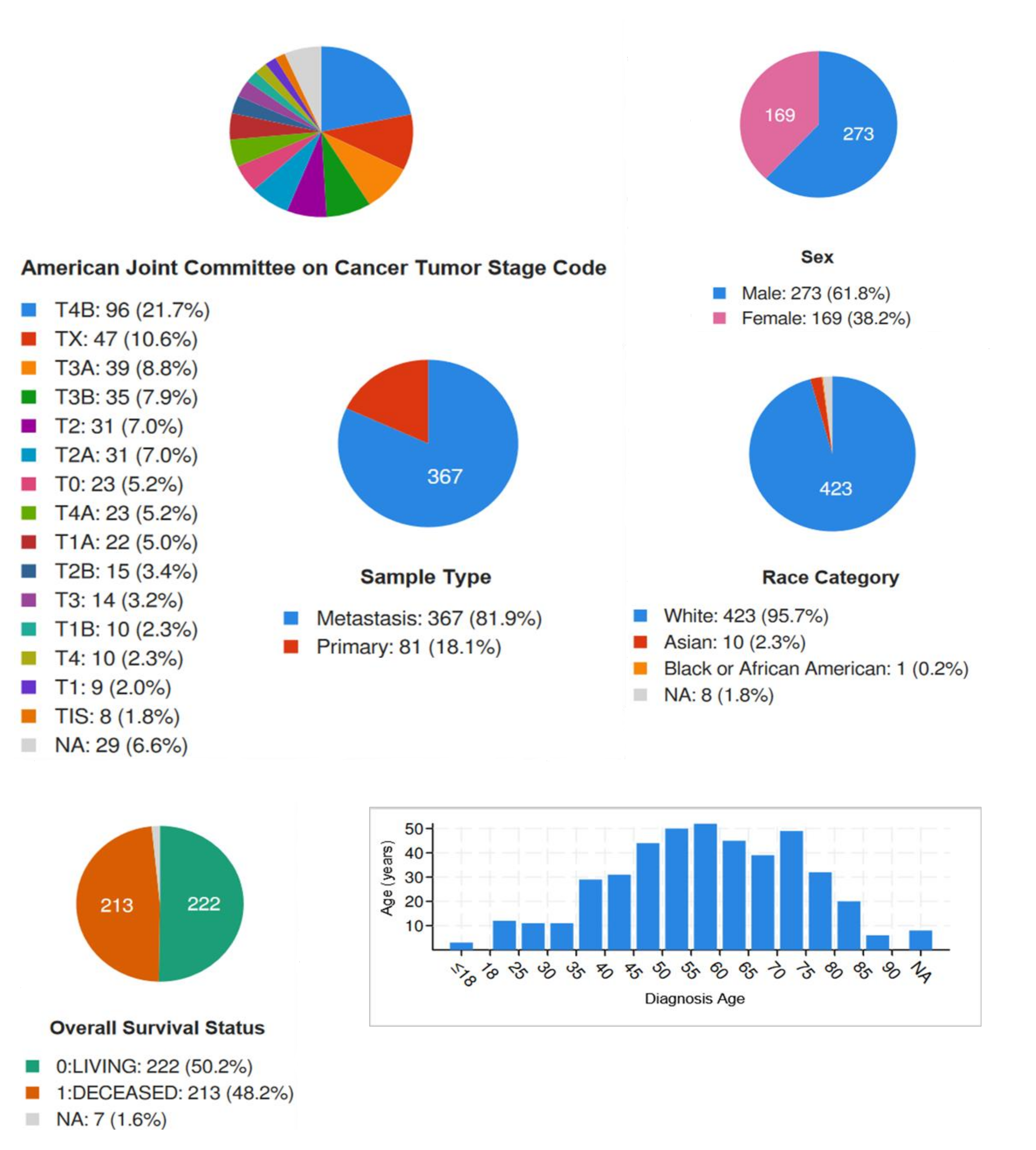
2. Results
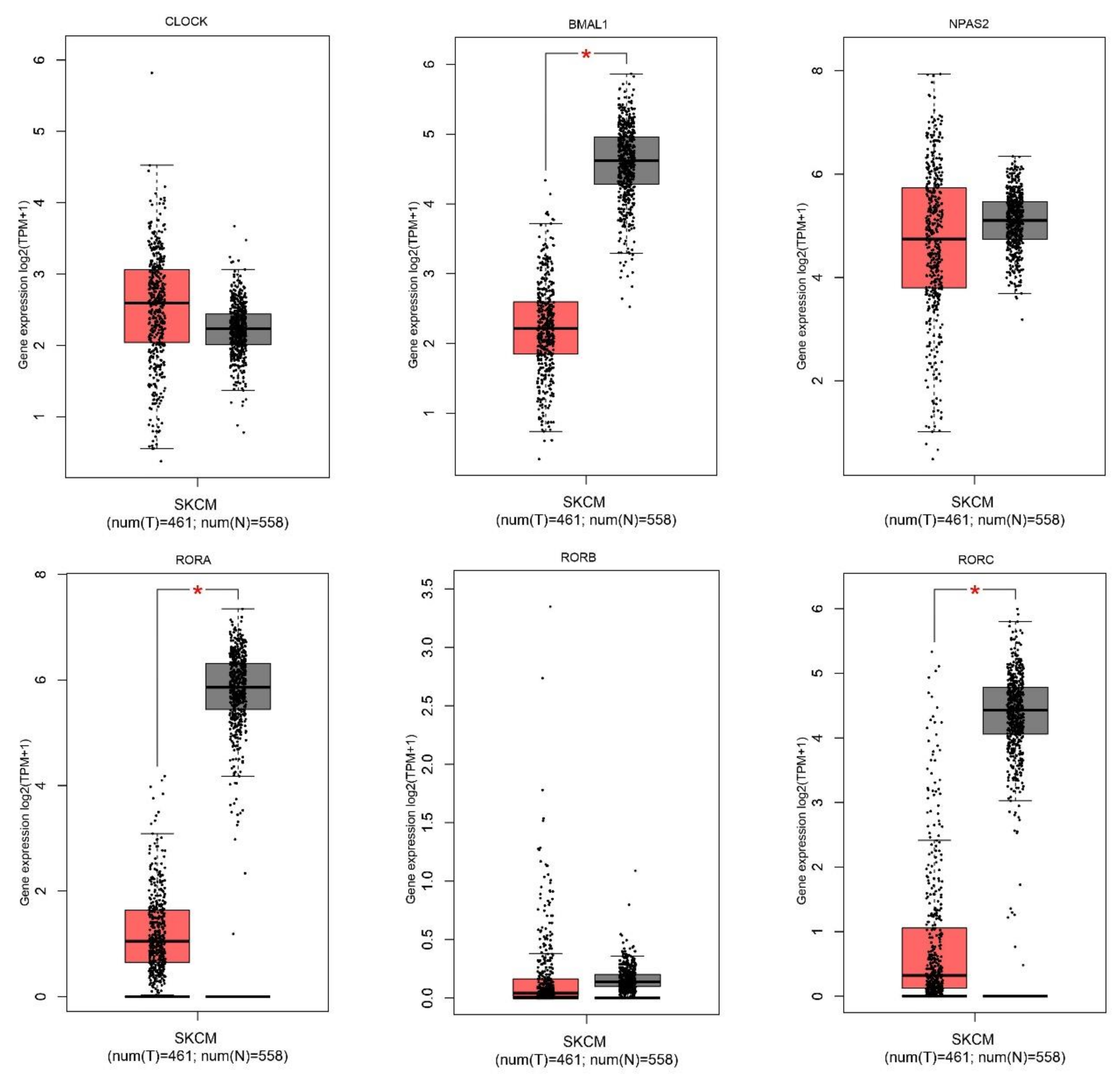
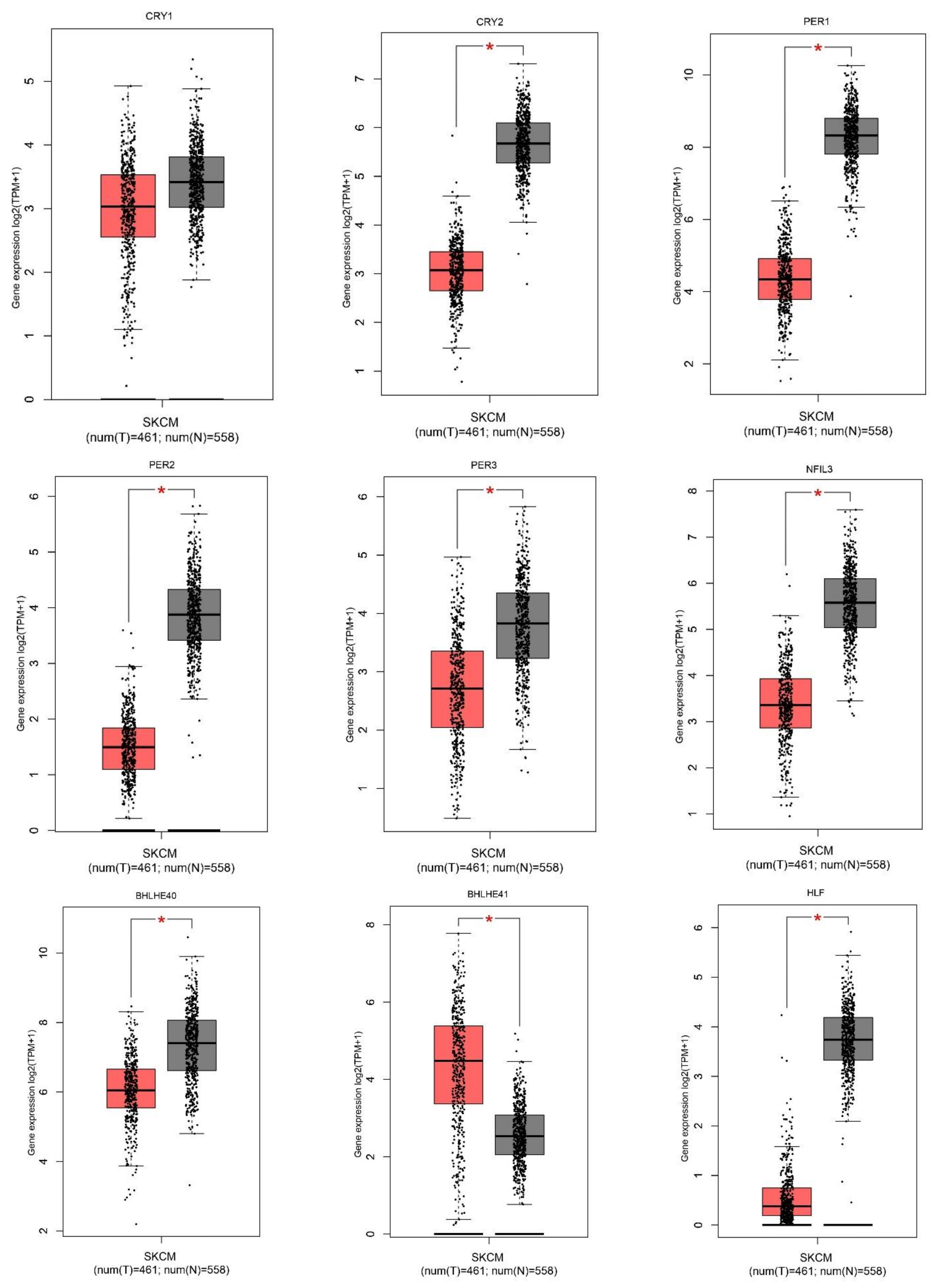
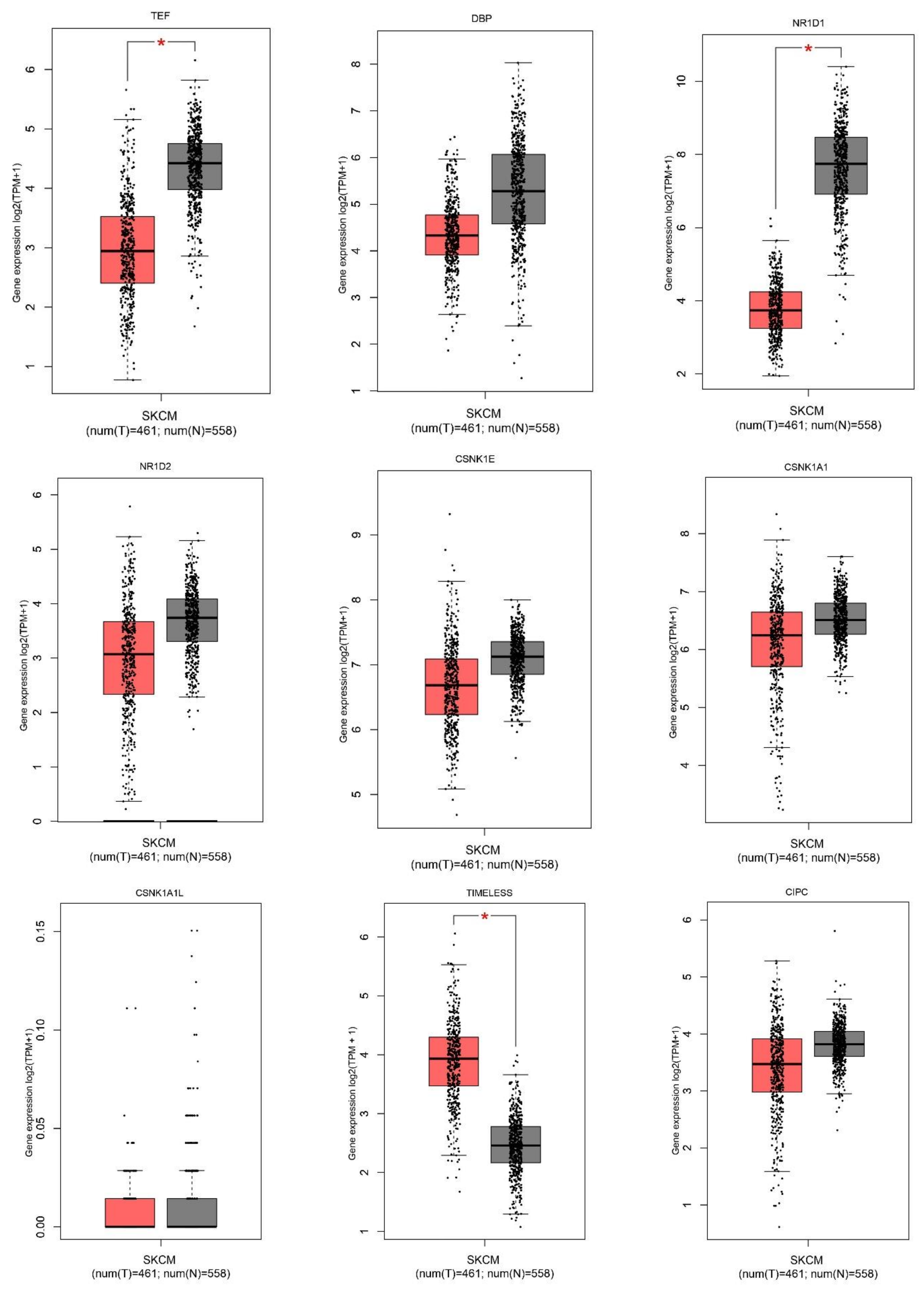
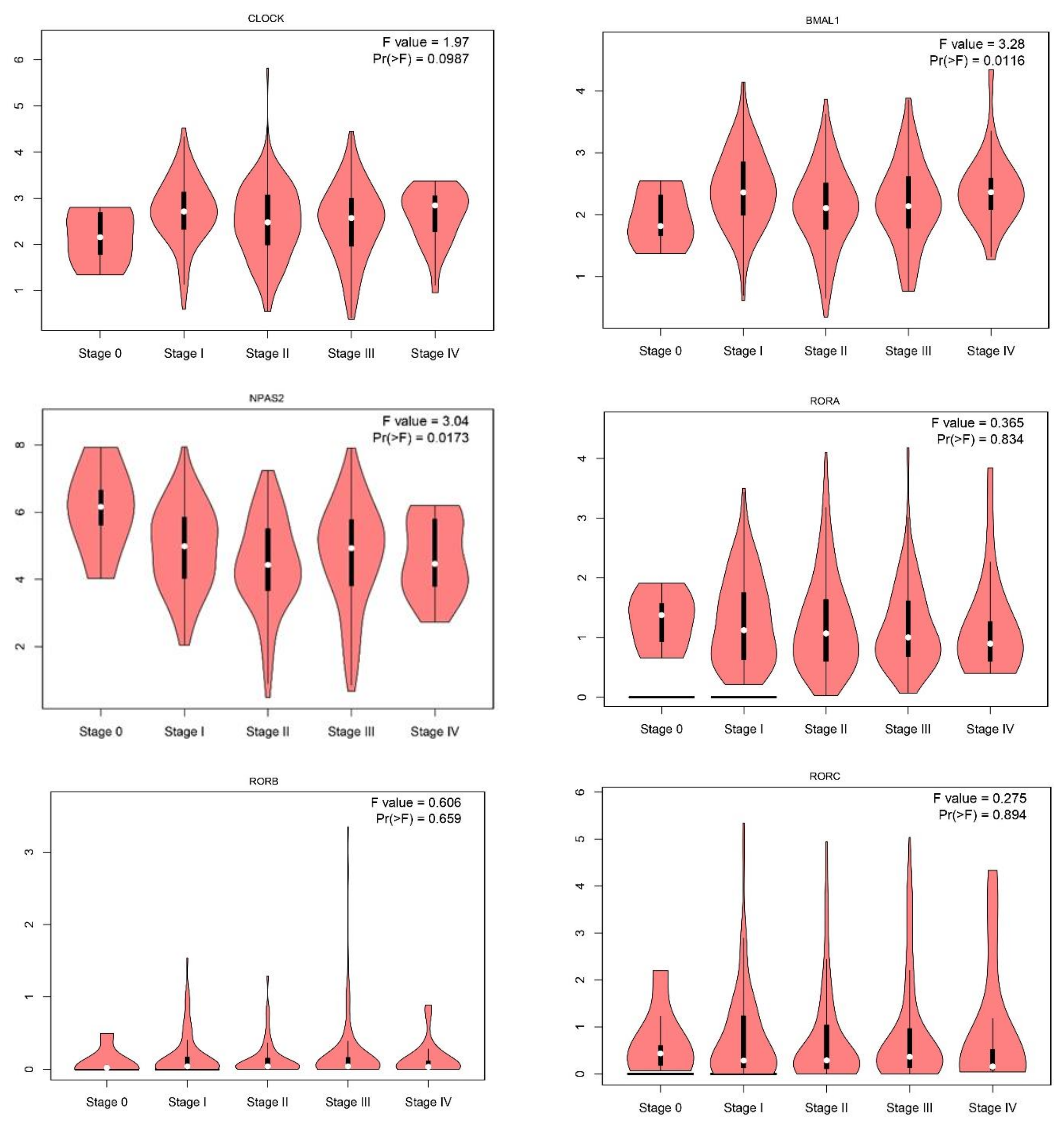
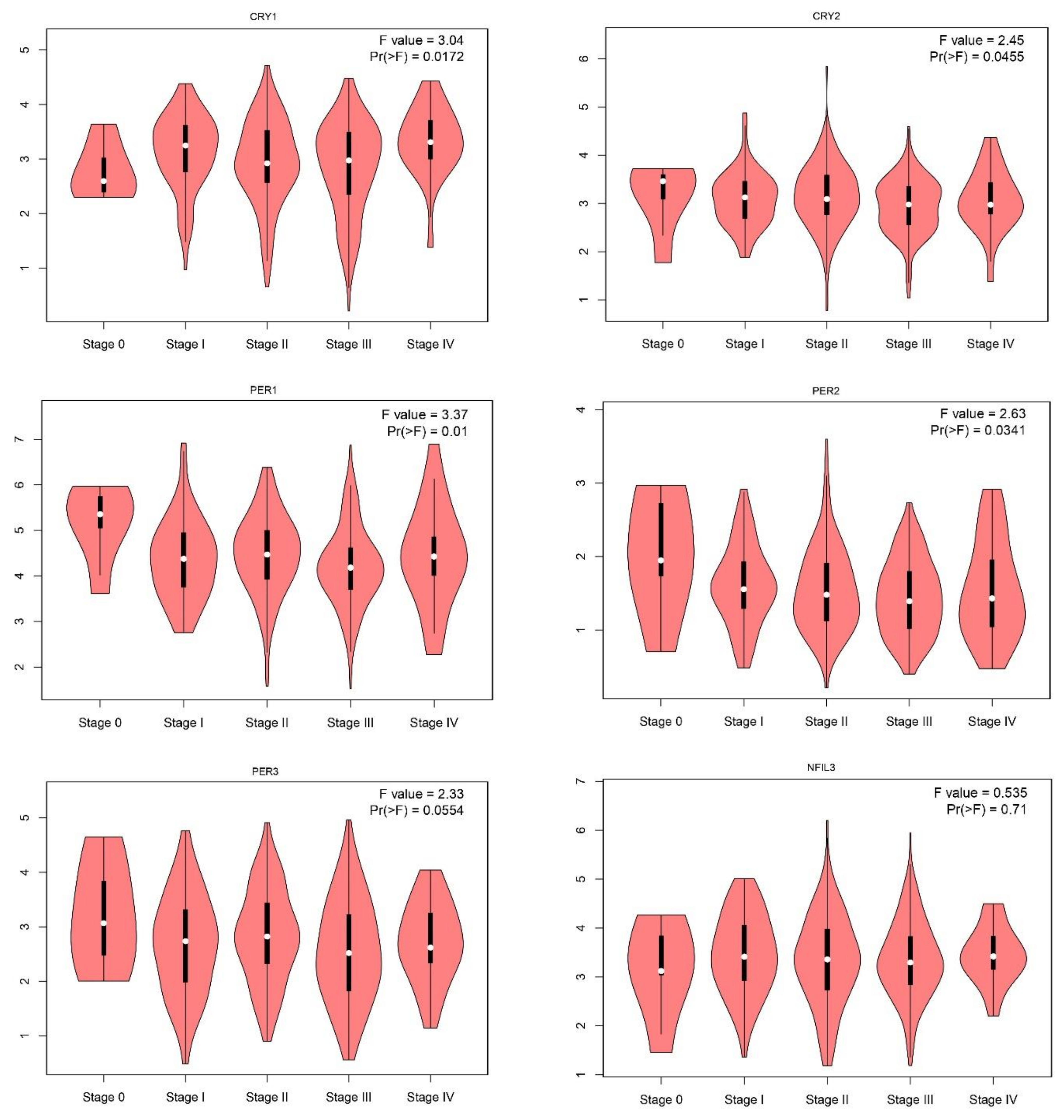
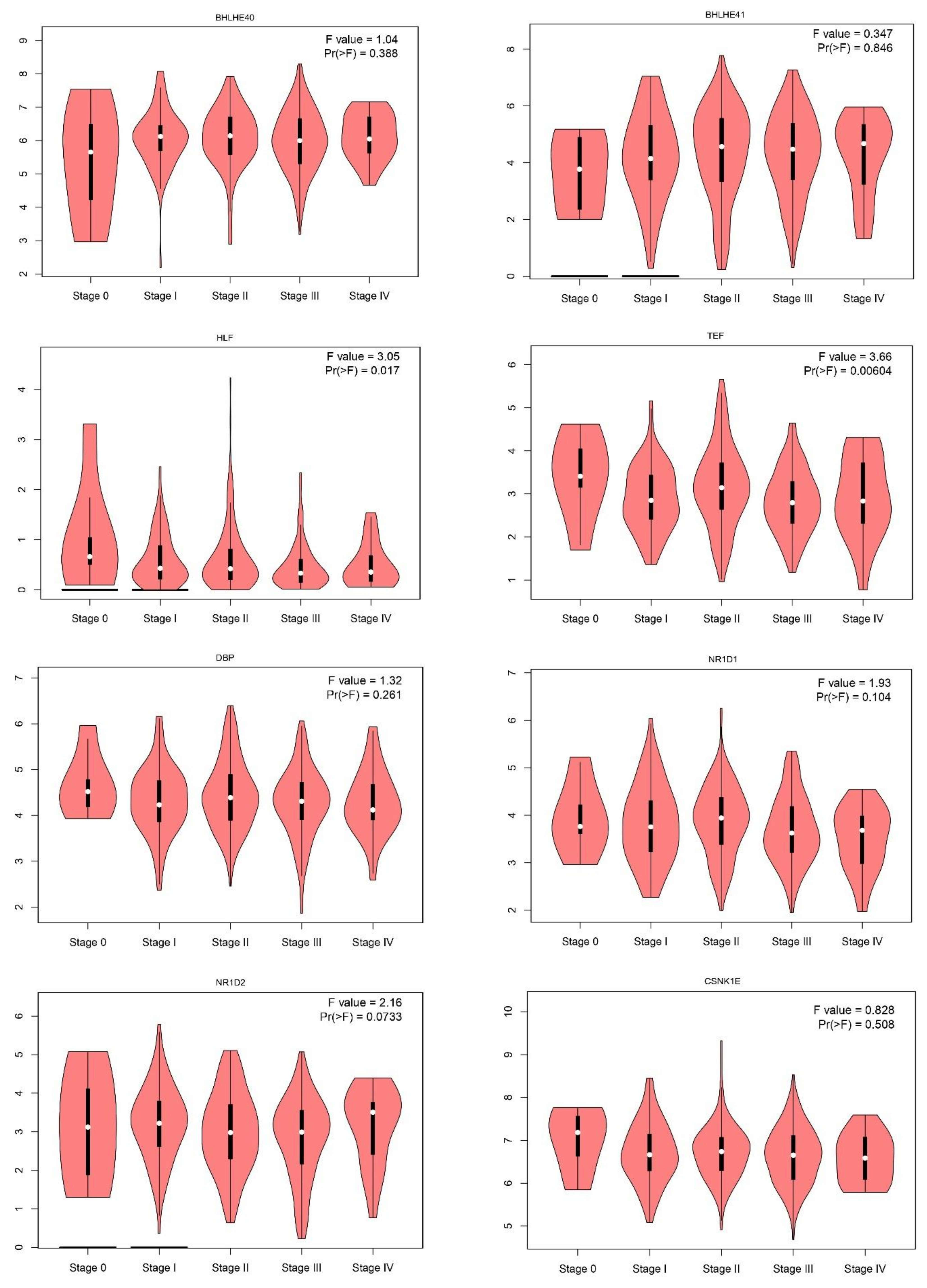
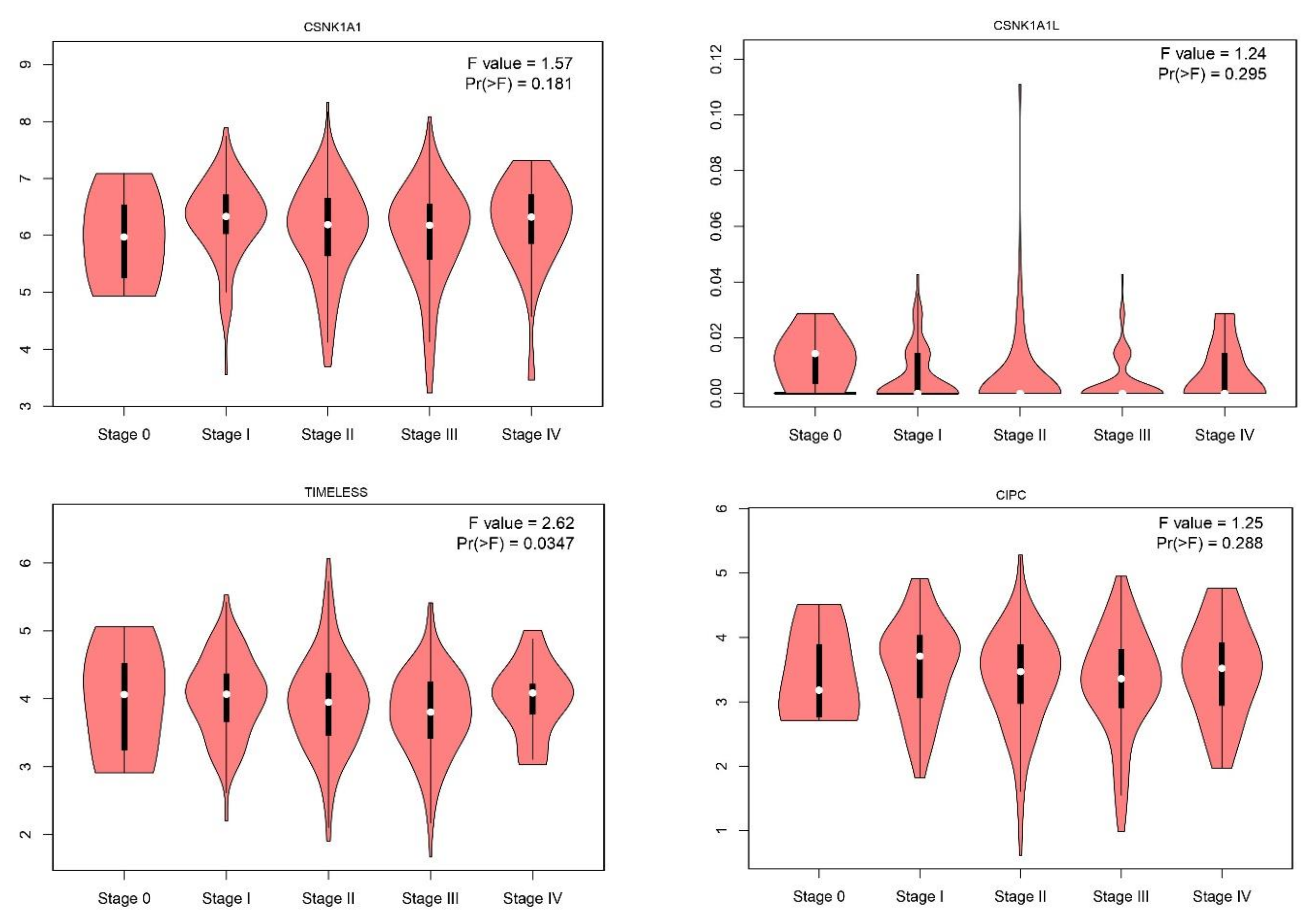
2.1. Gene Expression Level of the Core Circadian Genes in Skin Cutaneous Melanoma
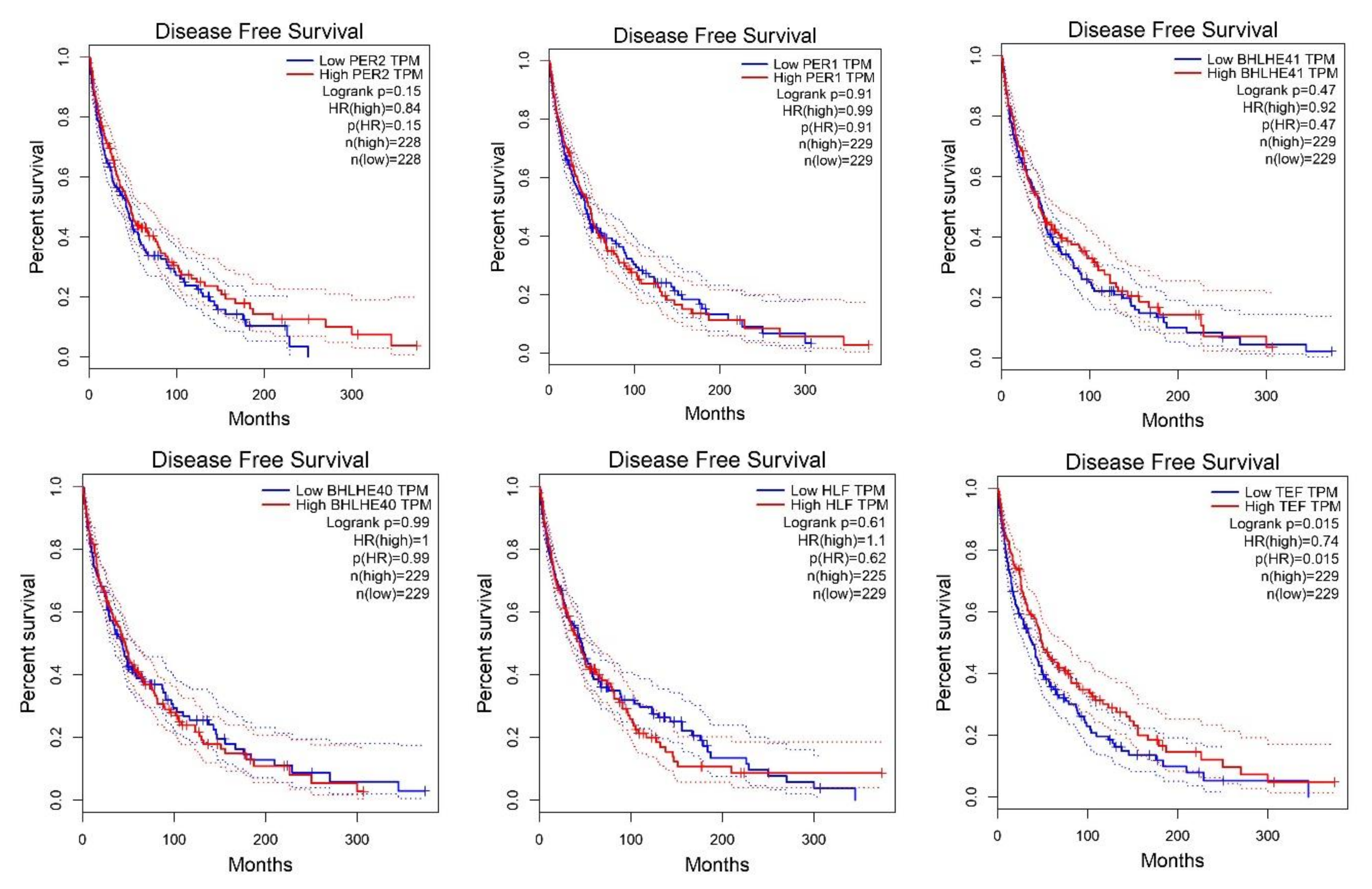
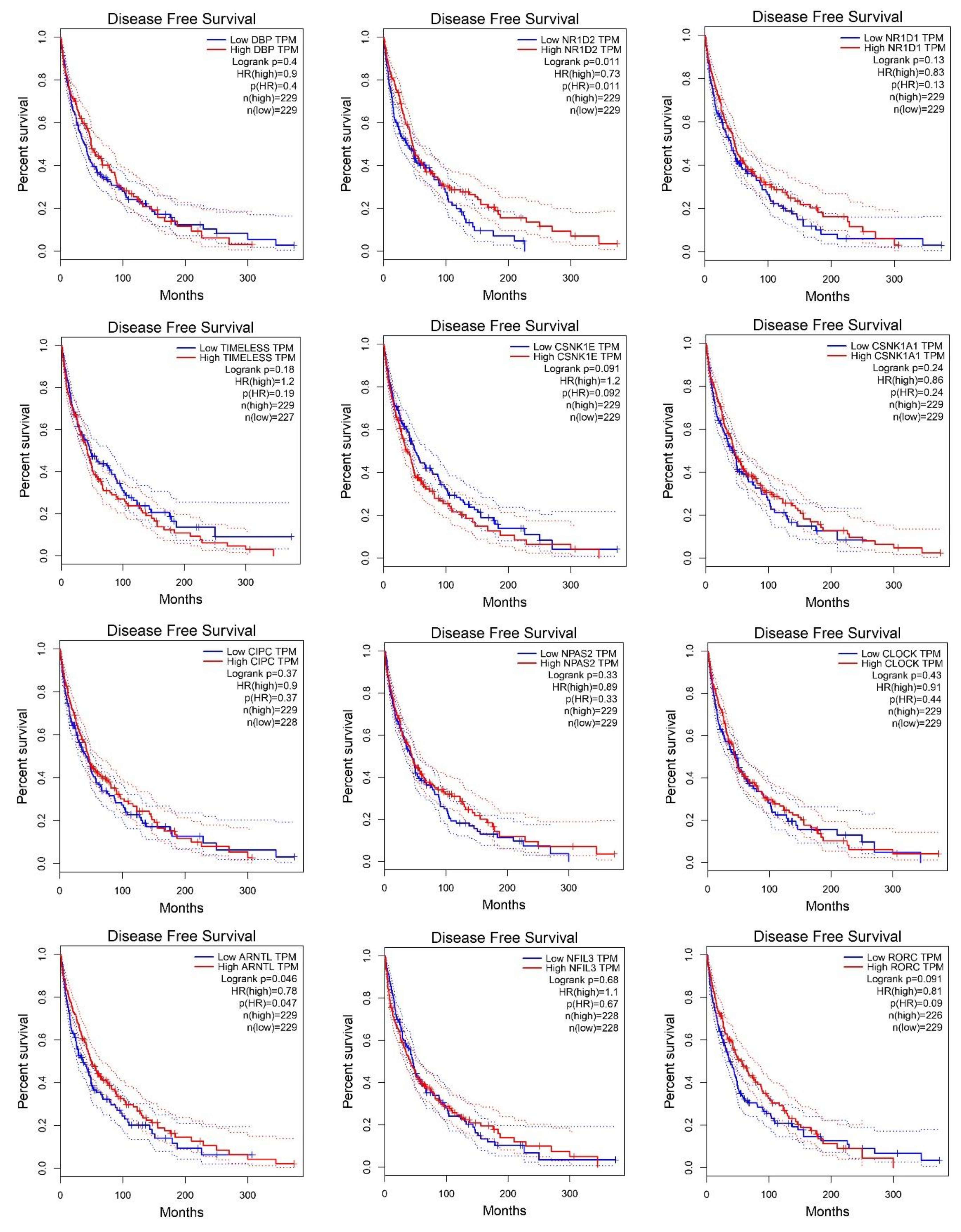
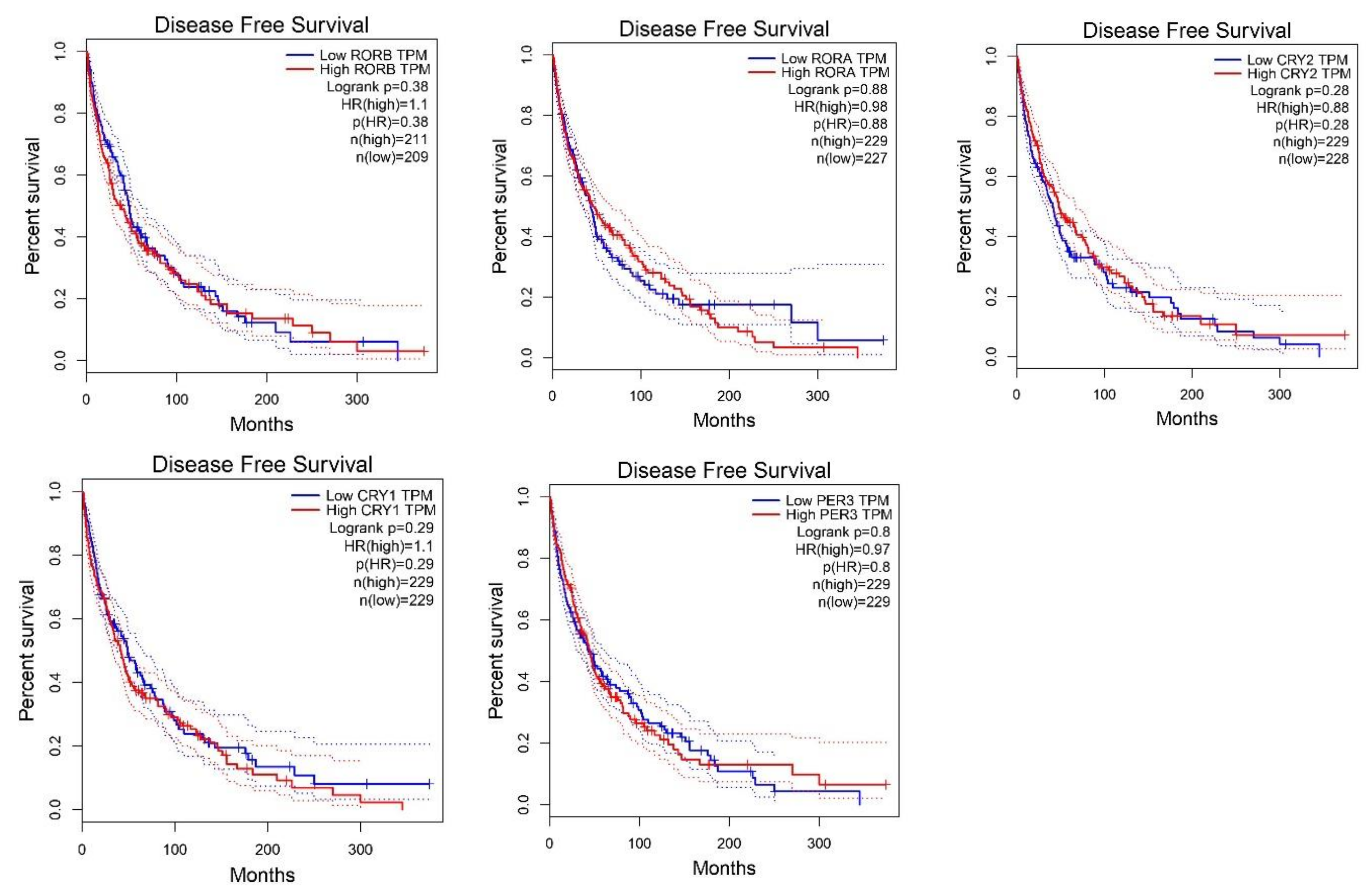
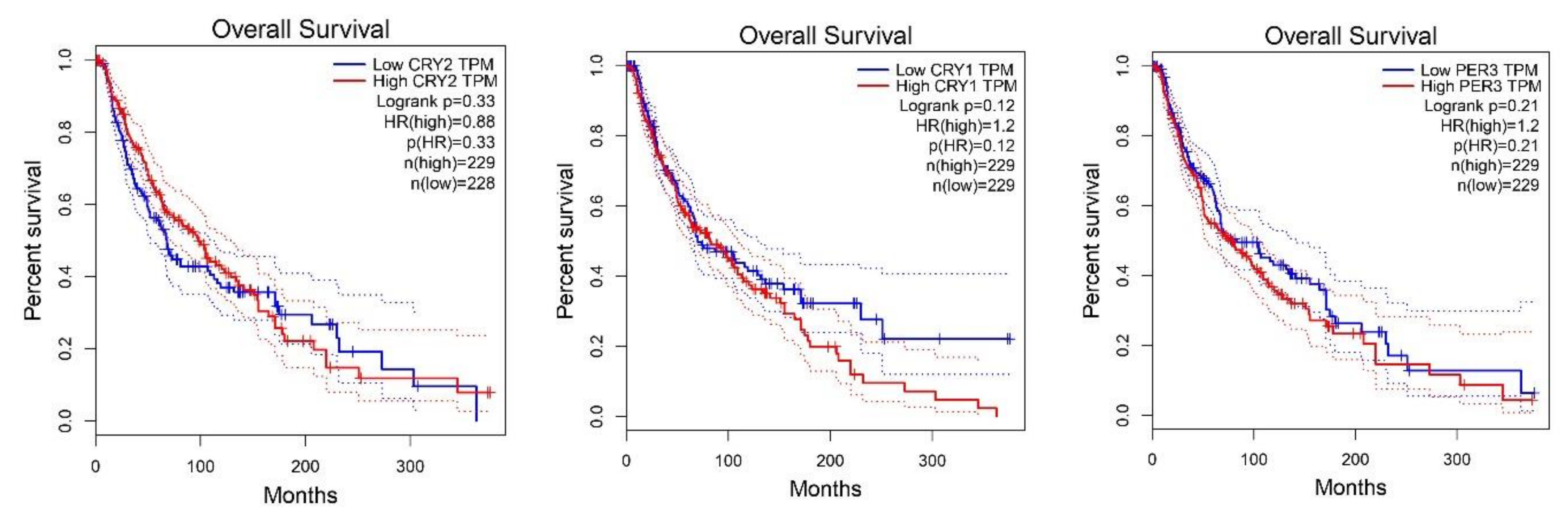
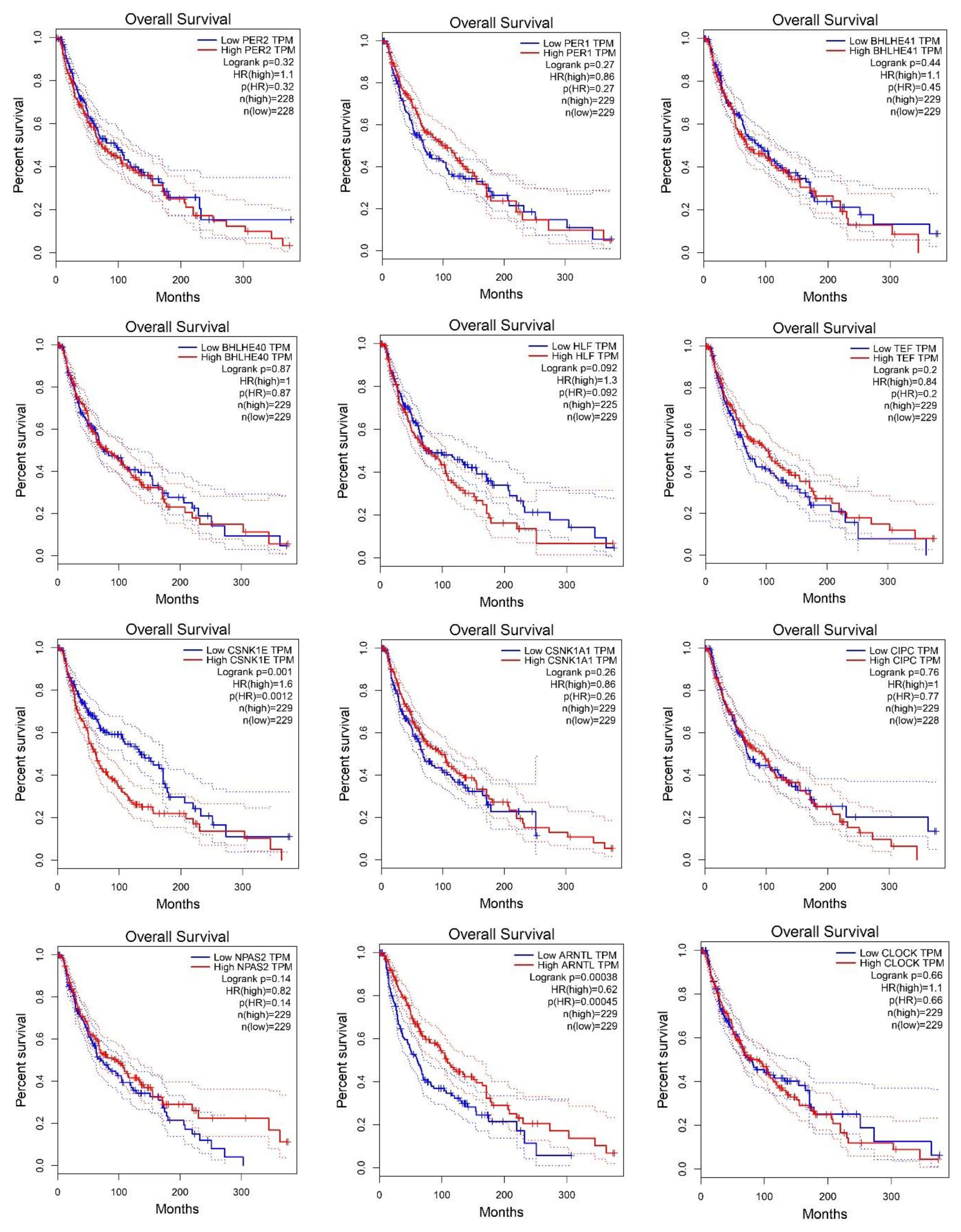
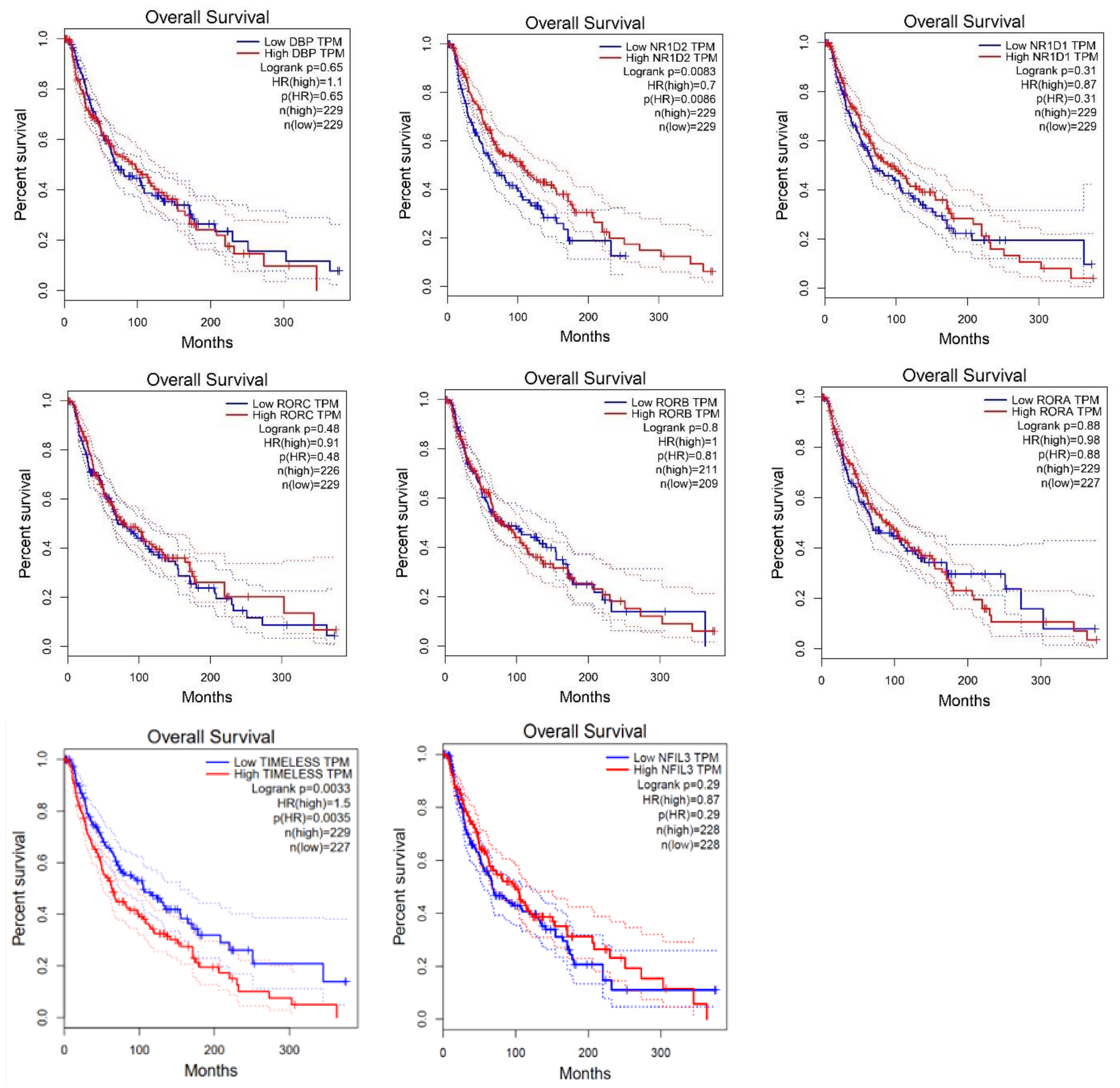
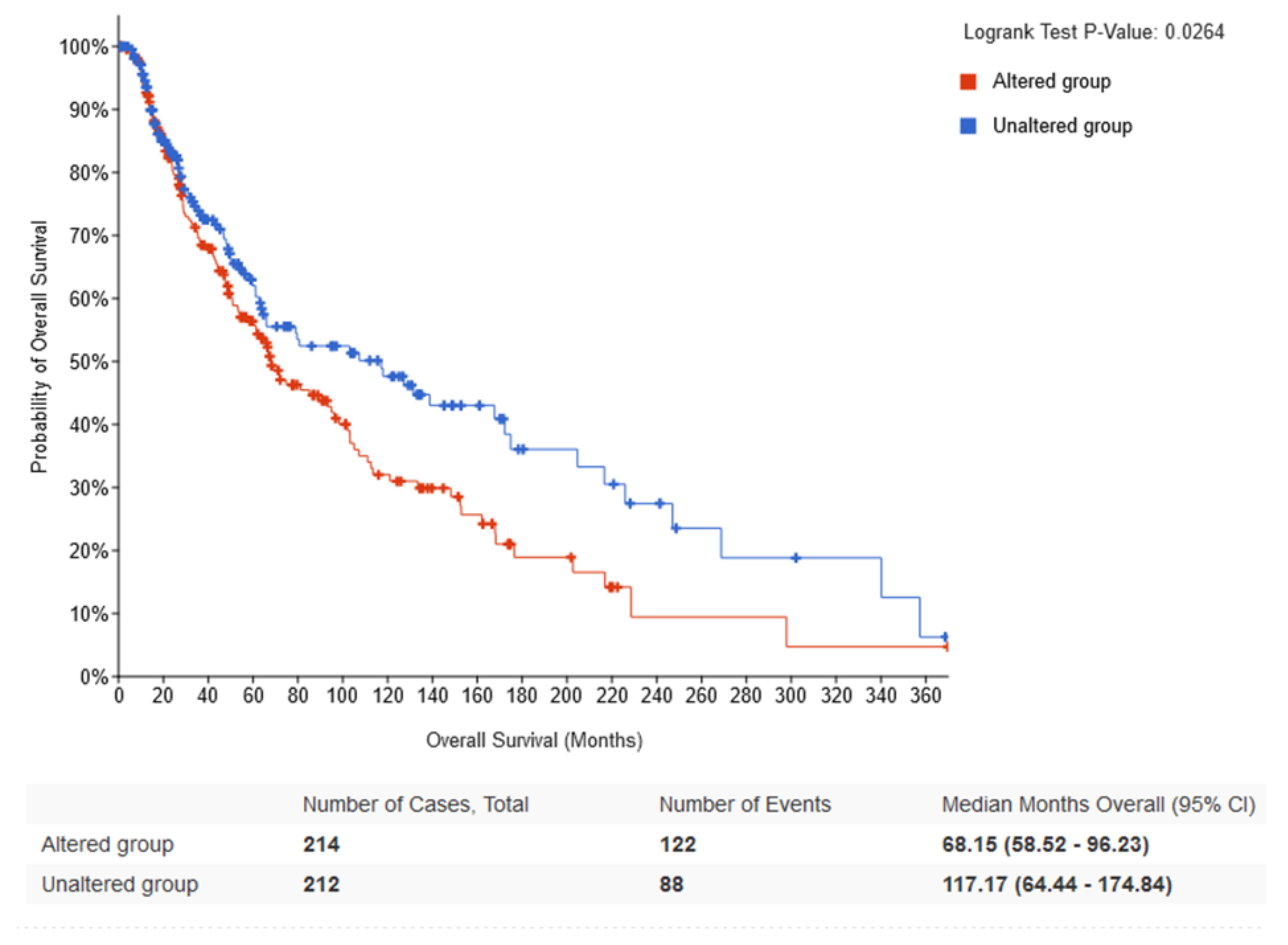
2.2. Survival Analysis According to Gene Expression Level of the Core Circadian Genes in Skin Cutaneous Melanoma
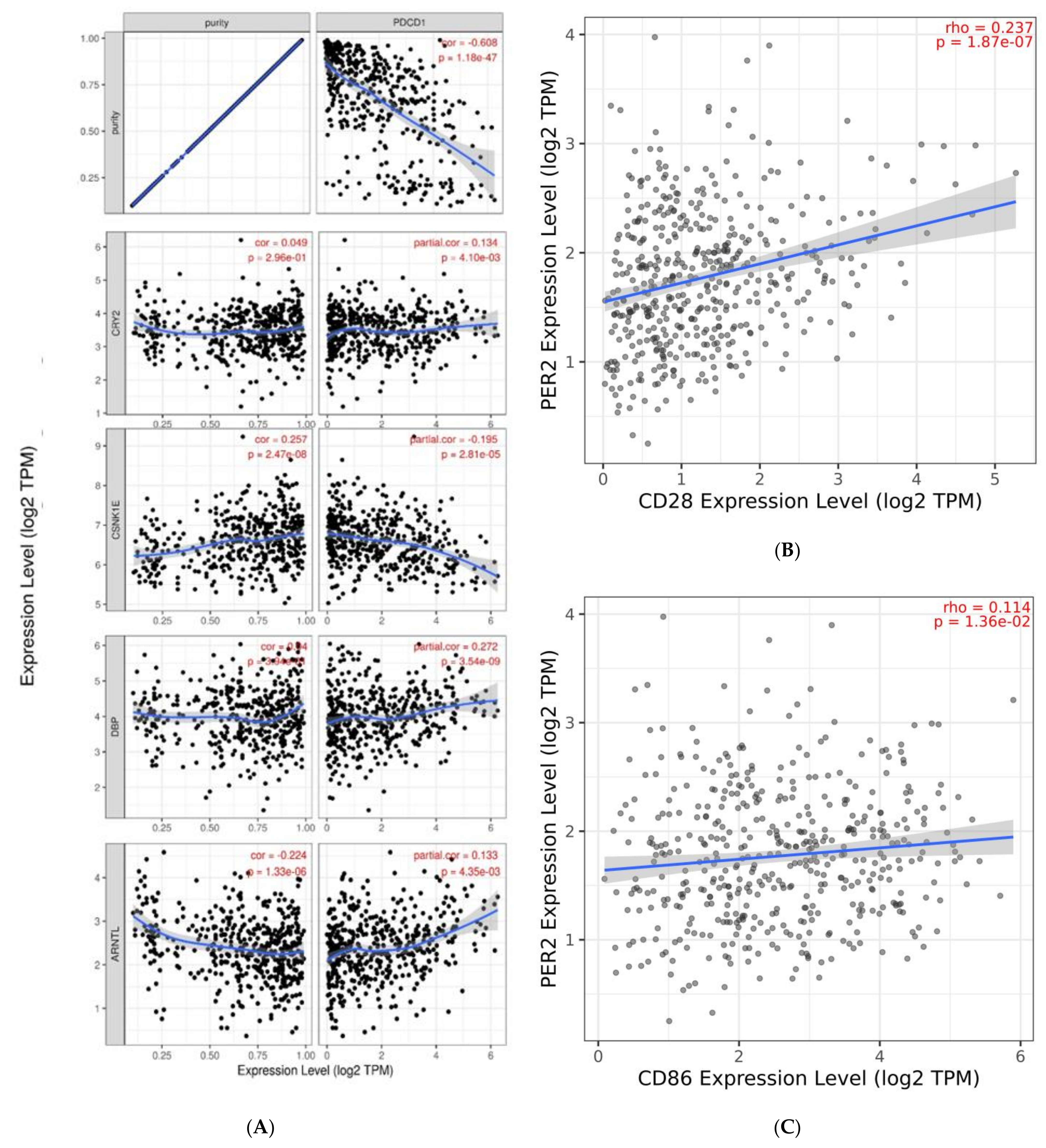
2.3. Infiltration Level
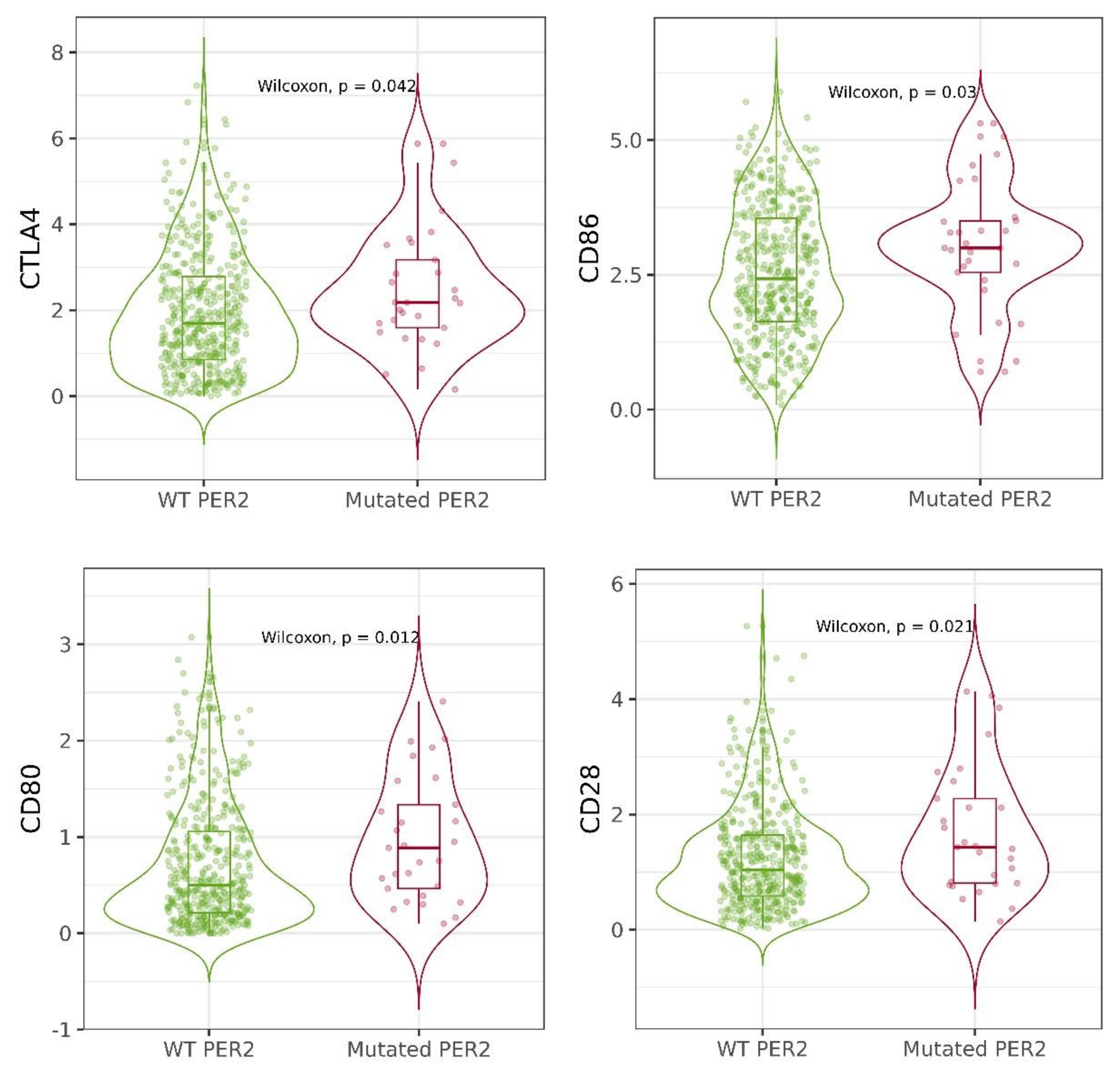
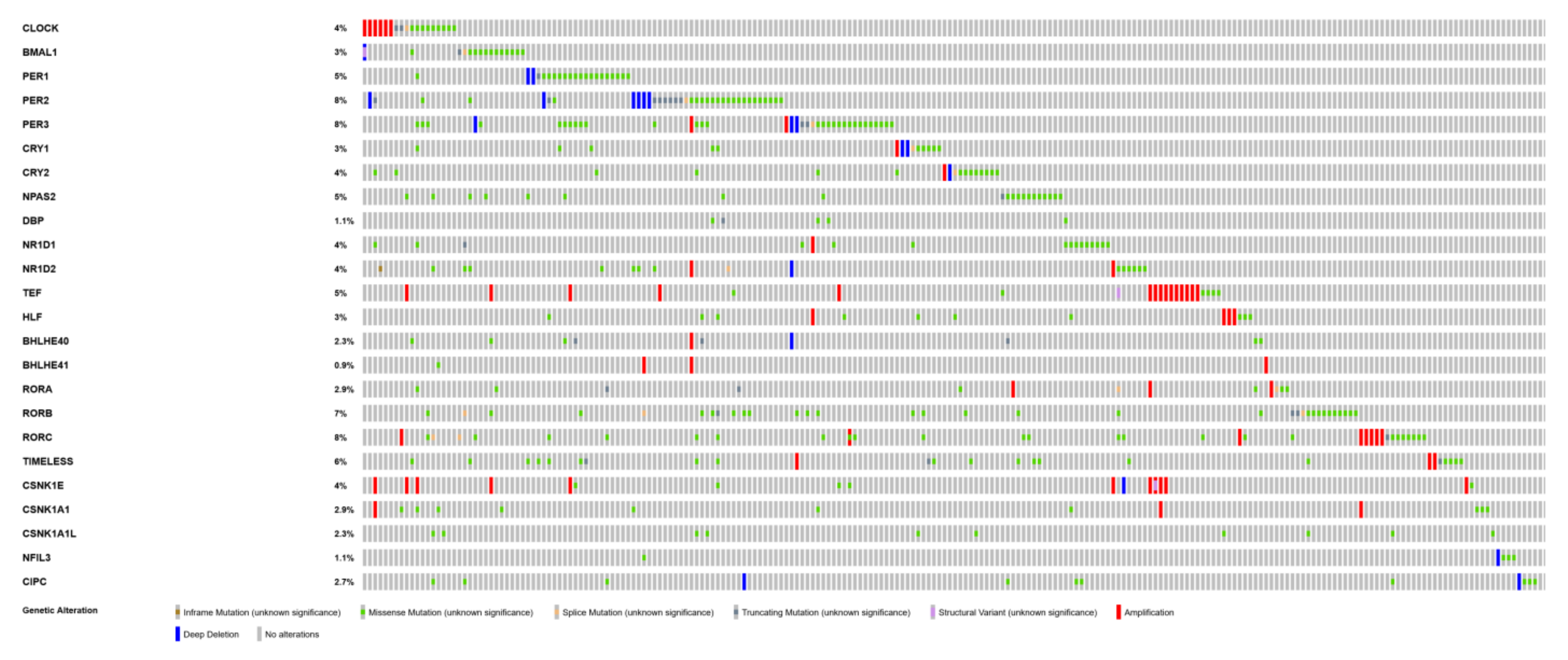
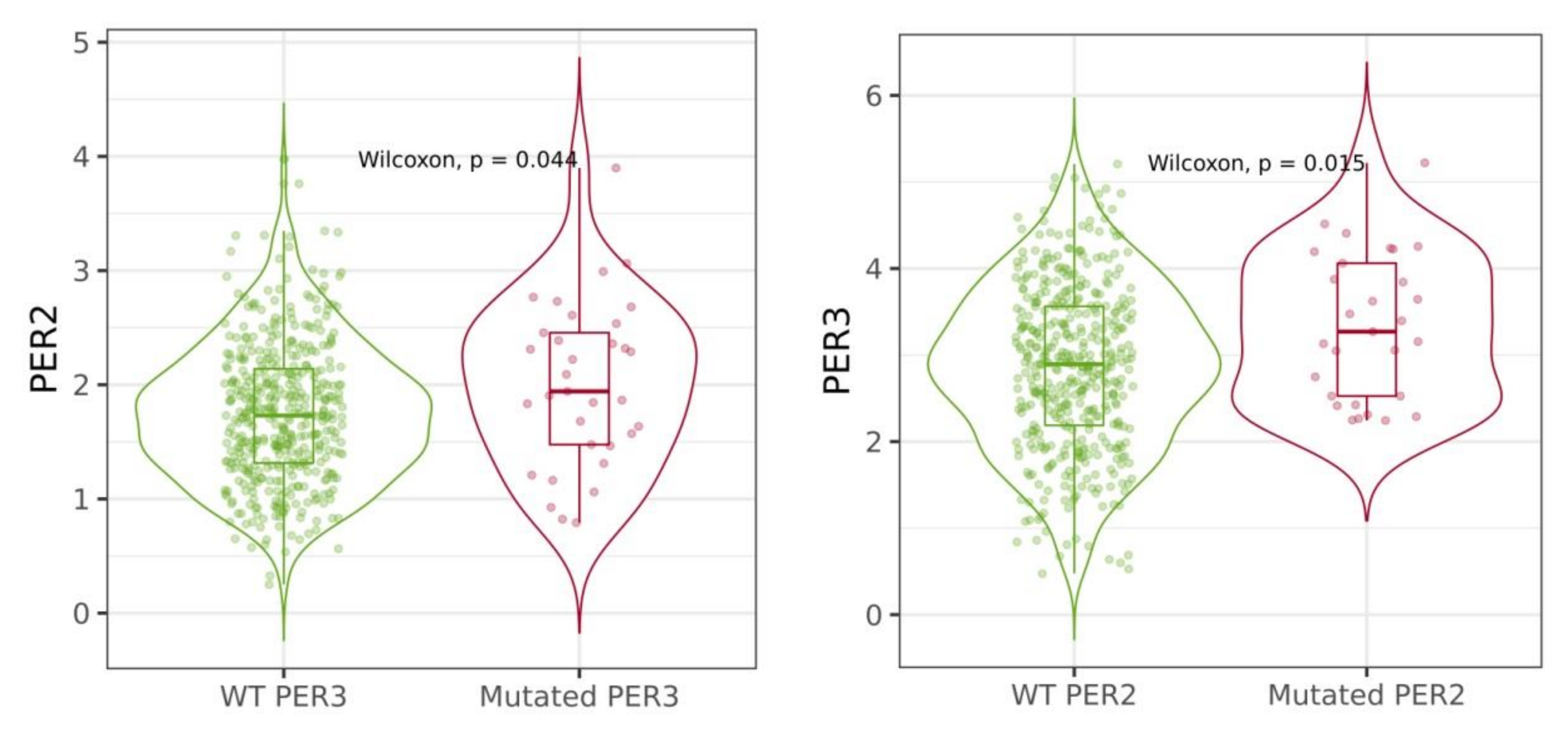
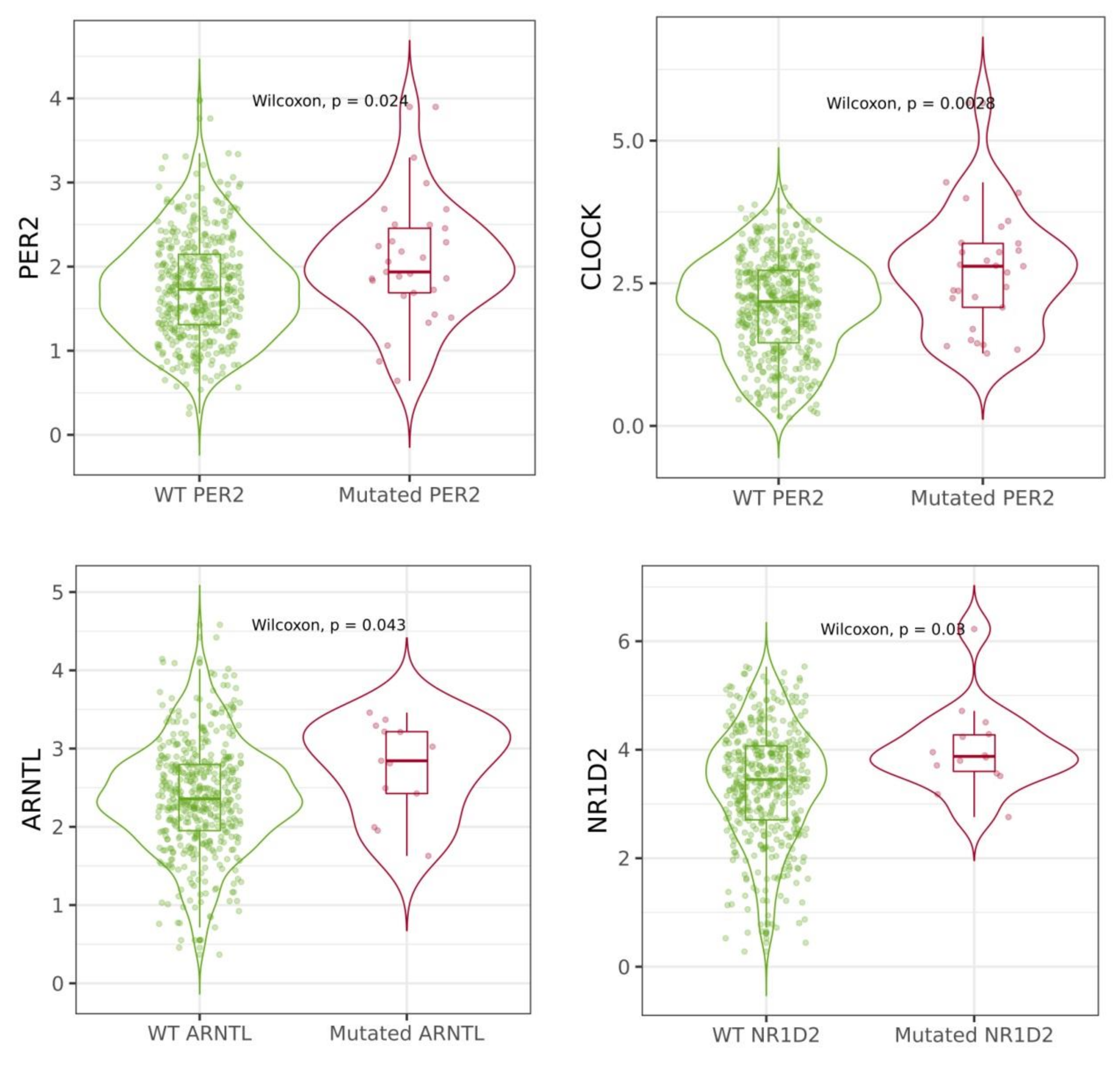
2.4. Mutations of Circadian Genes in Cutaneous Melanoma
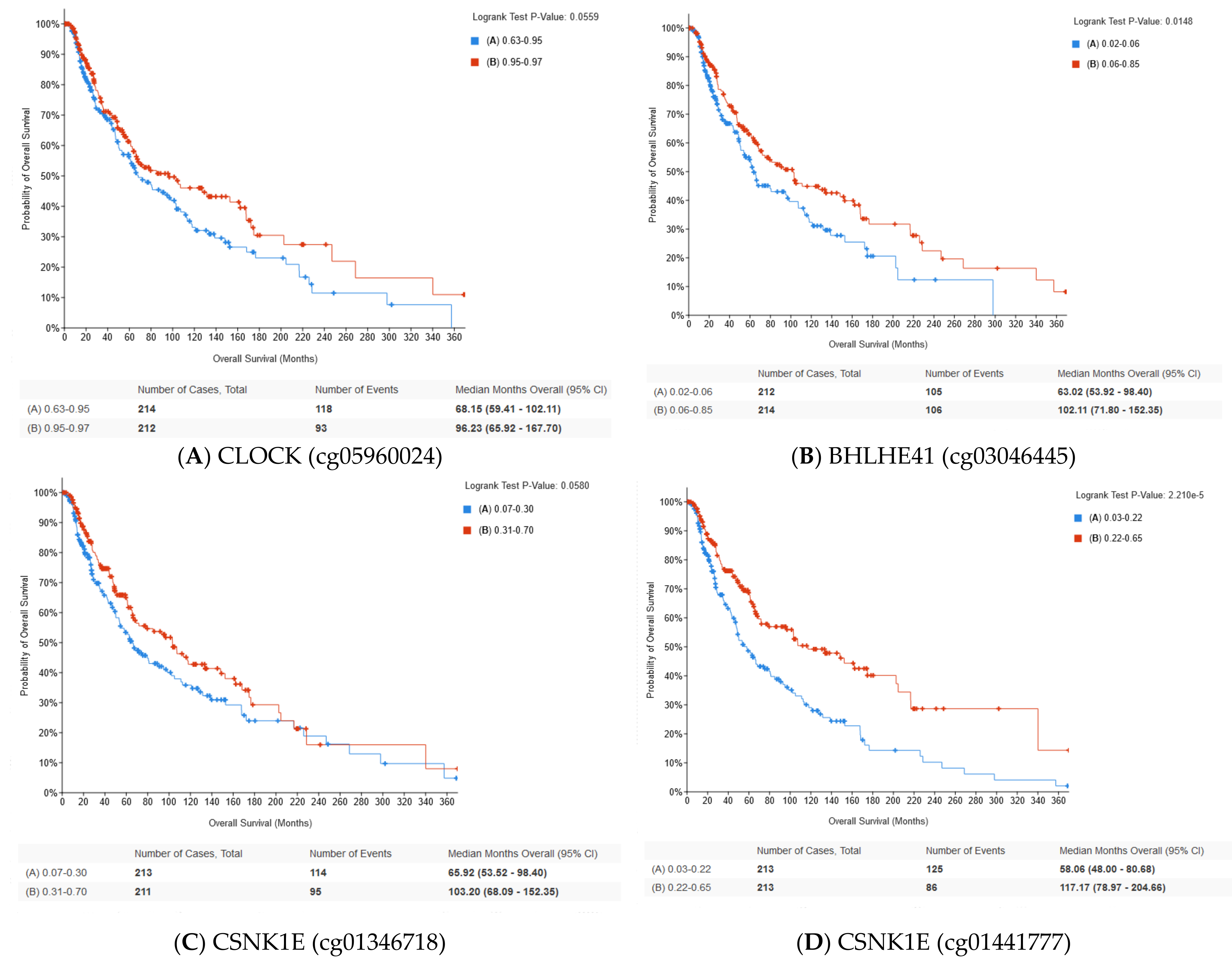
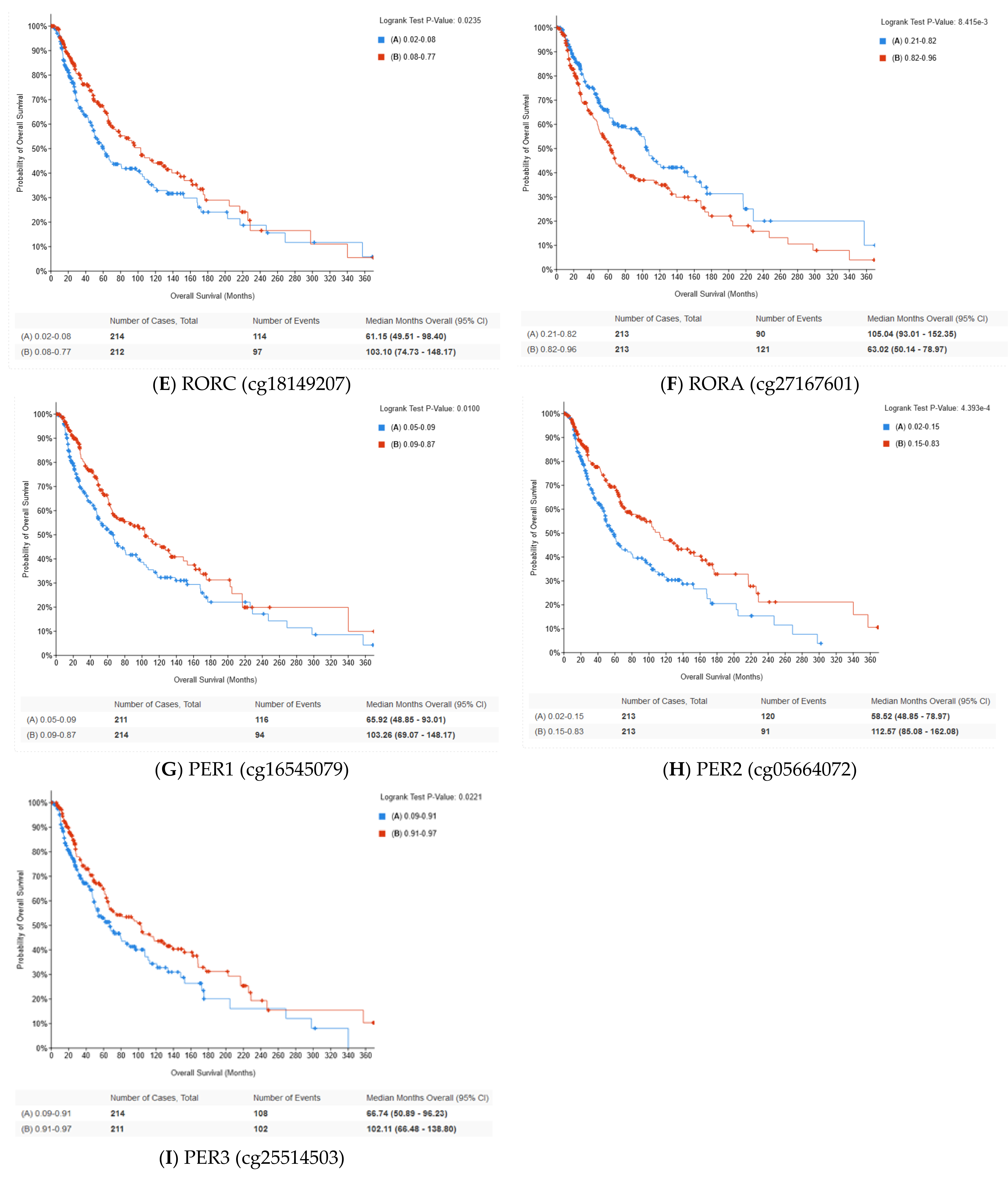
2.5. Methylation of Circadian Genes
3. Discussion
4. Materials and Methods
4.1. GEPIA—Gene Expression Profiling Interactive Analysis Database
4.2. TIMER 2.0—Database
4.3. cBioPortal
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Siegel, R.L.; Miller, K.D.; Wagle, N.S.; Jemal, A. Cancer statistics, 2023. CA Cancer J. Clin. 2023, 73, 17–48. [Google Scholar] [CrossRef] [PubMed]
- Janda, M.; Olsen, C.M.; Mar, J.; Cust, A.E. Early detection of skin cancer in Australia—Current approaches and new opportunities. Public. Health Res. Pract. 2022, 32, 3212204. [Google Scholar] [CrossRef] [PubMed]
- Gutierrez, D.; Arbesman, J. Circadian Dysrhythmias, Physiological Aberrations, and the Link to Skin Cancer. Int. J. Mol. Sci. 2016, 17, 621. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Markova-Car, E.P.; Jurišić, D.; Ilić, N.; Kraljević Pavelić, S. Running for time: Circadian rhythms and melanoma. Tumour Biol. J. Int. Soc. Oncodev. Biol. Med. 2014, 35, 8359–8368. [Google Scholar] [CrossRef] [PubMed]
- Aiello, I.; Mul Fedele, M.L.; Román, F.R.; Golombek, D.A.; Paladino, N. Circadian disruption induced by tumor development in a murine model of melanoma. Chronobiol. Int. 2022, 39, 12–25. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, J.S.; Hong, H.K.; Ko, C.H.; McDearmon, E.L. The genetics of mammalian circadian order and disorder: Implications for physiology and disease. Nat. Rev. Genet. 2008, 9, 764–775. [Google Scholar] [CrossRef]
- Fagiani, F.; Di Marino, D.; Romagnoli, A.; Travelli, C.; Voltan, D.; Di Cesare Mannelli, L.; Racchi, M.; Govoni, S.; Lanni, C. Molecular regulations of circadian rhythm and implications for physiology and diseases. Signal. Transduct. Target. Ther. 2022, 7, 41. [Google Scholar] [CrossRef]
- Rijo-Ferreira, F.; Takahashi, J.S. Genomics of circadian rhythms in health and disease. Genome Med. 2019, 11, 82. [Google Scholar] [CrossRef] [Green Version]
- Lubov, J.E.; Cvammen, W.; Kemp, M.G. The Impact of the Circadian Clock on Skin Physiology and Cancer Development. Int. J. Mol. Sci. 2021, 22, 6112. [Google Scholar] [CrossRef]
- Passarelli, A.; Mannavola, F.; Stucci, L.S.; Tucci, M.; Silvestris, F. Immune system and melanoma biology: A balance between immunosurveillance and immune escape. Oncotarget 2017, 8, 106132–106142. [Google Scholar] [CrossRef]
- Lengyel, Z.; Lovig, C.; Kommedal, S.; Keszthelyi, R.; Szekeres, G.; Battyáni, Z.; Csernus, V.; Nagy, A.D. Altered expression patterns of clock gene mRNAs and clock proteins in human skin tumors. Tumour Biol. J. Int. Soc. Oncodev. Biol. Med. 2013, 34, 811–819. [Google Scholar] [CrossRef]
- De Assis, L.V.M.; Kinker, G.S.; Moraes, M.N.; Markus, R.P.; Fernandes, P.A.; Castrucci, A.M.d.L. Expression of the Circadian Clock Gene BMAL1 Positively Correlates With Antitumor Immunity and Patient Survival in Metastatic Melanoma. Front. Oncol. 2018, 8, 185. [Google Scholar] [CrossRef] [Green Version]
- De Assis, L.V.M.; Moraes, M.N.; Magalhães-Marques, K.K.; Kinker, G.S.; da Silveira Cruz-Machado, S.; Castrucci, A.M.L. Non-Metastatic Cutaneous Melanoma Induces Chronodisruption in Central and Peripheral Circadian Clocks. Int. J. Mol. Sci. 2018, 19, 1065. [Google Scholar] [CrossRef] [Green Version]
- Savvidis, C.; Koutsilieris, M. Circadian rhythm disruption in cancer biology. Mol. Med. 2012, 18, 1249–1260. [Google Scholar] [CrossRef]
- Lee, C.C. Tumor suppression by the mammalian Period genes. Cancer Causes Control. CCC 2006, 17, 525–530. [Google Scholar] [CrossRef]
- Liu, H.; Gong, X.; Yang, K. Overexpression of the clock gene Per2 suppresses oral squamous cell carcinoma progression by activating autophagy via the PI3K/AKT/mTOR pathway. J. Cancer 2020, 11, 3655–3666. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Y.; Wu, Z.; Li, Y.; Zhang, J.; Gao, Y.; Yuan, G.; Han, M. PER3 plays anticancer roles in the oncogenesis and progression of breast cancer via regulating MEK/ERK signaling pathway. J. Chin. Med. Assoc. 2022, 85, 1051–1060. [Google Scholar] [CrossRef] [PubMed]
- Kettner, N.M.; Katchy, C.A.; Fu, L. Circadian gene variants in cancer. Ann. Med. 2014, 46, 208–220. [Google Scholar] [CrossRef] [Green Version]
- De Assis, L.V.M.; Lacerda, J.T.; Moraes, M.N.; Domínguez-Amorocho, O.A.; Kinker, G.S.; Mendes, D.; Silva, M.M.; Menck, C.F.M.; Câmara, N.O.S.; Castrucci, A.M.d.L. Melanopsin (Opn4) is an oncogene in cutaneous melanoma. Commun. Biol. 2022, 5, 461. [Google Scholar] [CrossRef] [PubMed]
- Na, H.; Han, J.; Ka, N.L.; Lee, M.H.; Choi, Y.L.; Shin, Y.K.; Lee, M.O. High expression of NR1D1 is associated with good prognosis in triple-negative breast cancer patients treated with chemotherapy. Breast Cancer Res. BCR 2019, 21, 127. [Google Scholar] [CrossRef] [Green Version]
- Huang, Z.; He, A.; Wang, J.; Lu, H.; Zhang, R.; Wu, L.; Feng, Q. The circadian clock is associated with prognosis and immune infiltration in stomach adenocarcinoma. Aging 2021, 13, 16637–16655. [Google Scholar] [CrossRef]
- Liu, B.; Xu, K.; Jiang, Y.; Li, X. Aberrant expression of Per1, Per2 and Per3 and their prognostic relevance in non-small cell lung cancer. Int. J. Clin. Exp. Pathol. 2014, 7, 7863–7871. [Google Scholar] [PubMed]
- Li, B.; Yang, X.; Li, J.; Yang, Y.; Yan, Z.; Zhang, H.; Mu, J. Elevated expression of CLOCK is associated with poor prognosis in hepatocellular carcinoma. Xi Bao Yu Fen. Zi Mian Yi Xue Za Zhi = Chin. J. Cell. Mol. Immunol. 2018, 34, 163–168. [Google Scholar]
- Falcone, I.; Conciatori, F.; Bazzichetto, C.; Ferretti, G.; Cognetti, F.; Ciuffreda, L.; Milella, M. Tumor Microenvironment: Implications in Melanoma Resistance to Targeted Therapy and Immunotherapy. Cancers 2020, 12, 2870. [Google Scholar] [CrossRef] [PubMed]
- Simiczyjew, A.; Dratkiewicz, E.; Mazurkiewicz, J.; Ziętek, M.; Matkowski, R.; Nowak, D. The Influence of Tumor Microenvironment on Immune Escape of Melanoma. Int. J. Mol. Sci. 2020, 21, 8359. [Google Scholar] [CrossRef] [PubMed]
- Li, T.; Fan, J.; Wang, B.; Traugh, N.; Chen, Q.; Liu, J.S.; Li, B.; Liu, X.S. TIMER: A Web Server for Comprehensive Analysis of Tumor-Infiltrating Immune Cells. Cancer Res. 2017, 77, e108–e110. [Google Scholar] [CrossRef] [Green Version]
- Li, T.; Fu, J.; Zeng, Z.; Cohen, D.; Li, J.; Chen, Q.; Li, B.; Liu, X.S. TIMER2.0 for analysis of tumor-infiltrating immune cells. Nucleic Acids Res. 2020, 48, W509–W514. [Google Scholar] [CrossRef]
- Zhang, Z.; Zeng, P.; Gao, W.; Zhou, Q.; Feng, T.; Tian, X. Circadian clock: A regulator of the immunity in cancer. Cell. Commun. Signal. 2021, 19, 37. [Google Scholar] [CrossRef]
- Qin, R.; Peng, W.; Wang, X.; Li, C.; Xi, Y.; Zhong, Z.; Sun, C. Identification of Genes Related to Immune Infiltration in the Tumor Microenvironment of Cutaneous Melanoma. Front. Oncol. 2021, 11, 615963. [Google Scholar] [CrossRef]
- Fridman, W.H.; Pagès, F.; Sautès-Fridman, C.; Galon, J. The immune contexture in human tumours: Impact on clinical outcome. Nat. Rev. Cancer 2012, 12, 298–306. [Google Scholar] [CrossRef]
- Taggart, D.; Andreou, T.; Scott, K.J.; Williams, J.; Rippaus, N.; Brownlie, R.J.; Ilett, E.J.; Salmond, R.J.; Melcher, A.; Lorger, M. Anti–PD-1/anti–CTLA-4 efficacy in melanoma brain metastases depends on extracranial disease and augmentation of CD8+ T cell trafficking. Proc. Natl. Acad. Sci. USA 2018, 115, E1540–E1549. [Google Scholar] [CrossRef] [Green Version]
- Forsthuber, A.; Lipp, K.; Andersen, L.; Ebersberger, S.; Graña, C.; Ellmeier, W.; Petzelbauer, P.; Lichtenberger, B.M.; Loewe, R. CXCL5 as Regulator of Neutrophil Function in Cutaneous Melanoma. J. Investig. Dermatol. 2019, 139, 186–194. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, X.; Rollins, D.; Ruhn, K.A.; Stubblefield, J.J.; Green, C.B.; Kashiwada, M.; Rothman, P.B.; Takahashi, J.S.; Hooper, L.V. TH17 cell differentiation is regulated by the circadian clock. Science 2013, 342, 727–730. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Poole, J.; Kitchen, G.B. Circadian regulation of innate immunity in animals and humans and implications for human disease. Semin. Immunopathol. 2022, 44, 183–192. [Google Scholar] [CrossRef] [PubMed]
- Xu, H.; Huang, L.; Zhao, J.; Chen, S.; Liu, J.; Li, G. The circadian clock and inflammation: A new insight. Clin. Chim. Acta Int. J. Clin. Chem. 2021, 512, 12–17. [Google Scholar] [CrossRef] [PubMed]
- Spengler, M.L.; Kuropatwinski, K.K.; Comas, M.; Gasparian, A.V.; Fedtsova, N.; Gleiberman, A.S.; Gitlin, I.I.; Artemicheva, N.M.; Deluca, K.A.; Gudkov, A.V.; et al. Core circadian protein CLOCK is a positive regulator of NF-κB-mediated transcription. Proc. Natl. Acad. Sci. USA 2012, 109, E2457–E2465. [Google Scholar] [CrossRef] [Green Version]
- Mirian, M.; Hariri, A.; Yadollahi, M.; Kohandel, M. Circadian and Immunity Cycle Talk in Cancer Destination: From Biological Aspects to In Silico Analysis. Cancers 2022, 14, 1578. [Google Scholar] [CrossRef]
- Lam, M.T.Y.; Cho, H.; Lesch, H.P.; Gosselin, D.; Heinz, S.; Tanaka-Oishi, Y.; Benner, C.; Kaikkonen, M.U.; Kim, A.S.; Kosaka, M.; et al. Rev-Erbs repress macrophage gene expression by inhibiting enhancer-directed transcription. Nature 2013, 498, 511–515. [Google Scholar] [CrossRef] [Green Version]
- Hergenhan, S.; Holtkamp, S.; Scheiermann, C. Molecular Interactions between Components of the Circadian Clock and the Immune System. J. Mol. Biol. 2020, 432, 3700–3713. [Google Scholar] [CrossRef]
- Tang, Z.; Kang, B.; Li, C.; Chen, T.; Zhang, Z. GEPIA2: An enhanced web server for large-scale expression profiling and interactive analysis. Nucleic Acids Res. 2019, 47, W556–W560. [Google Scholar] [CrossRef] [Green Version]
- Tang, Z.; Li, C.; Kang, B.; Gao, G.; Li, C.; Zhang, Z. GEPIA: A web server for cancer and normal gene expression profiling and interactive analyses. Nucleic Acids Res. 2017, 45, W98–W102. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cerami, E.; Gao, J.; Dogrusoz, U.; Gross, B.E.; Sumer, S.O.; Aksoy, B.A.; Jacobsen, A.; Byrne, C.J.; Heuer, M.L.; Larsson, E.; et al. The cBio cancer genomics portal: An open platform for exploring multidimensional cancer genomics data. Cancer Discov. 2012, 2, 401–404. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gao, J.; Aksoy, B.A.; Dogrusoz, U.; Dresdner, G.; Gross, B.; Sumer, S.O.; Sun, Y.; Jacobsen, A.; Sinha, R.; Larsson, E.; et al. Integrative analysis of complex cancer genomics and clinical profiles using the cBioPortal. Sci. Signal. 2013, 6, pl1. [Google Scholar] [CrossRef] [PubMed] [Green Version]
























{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| SKCM n = 471 | SKCM—Primary N = 103 | SKCM—Metastasis N = 368 | |||||||
|---|---|---|---|---|---|---|---|---|---|
| HR | 95% CI_l | p | HR | 95% CI_l | p | HR | 95% CI_l | p | |
| CLOCK | 0.90 | 0.757–1.081 | 0.27 | 1.13 | 0.685–1.849 | 0.64 | 0.89 | 0.733–1.086 | 0.26 |
| ARNTL/BMAL1 | 0.71 | 0.562–0.889 | 0.00 | 1.16 | 0.643–2.108 | 0.62 | 0.68 | 0.529–0.877 | 0.00 |
| CRY1 | 0.97 | 0.787–1.183 | 0.73 | 1.18 | 0.673–2.053 | 0.57 | 0.96 | 0.763–1.199 | 0.70 |
| CRY2 | 0.82 | 0.637–1.043 | 0.11 | 1.10 | 0.621–1.979 | 0.72 | 0.83 | 0.63–1.086 | 0.17 |
| PER1 | 0.93 | 0.775–1.107 | 0.40 | 1.03 | 0.591–1.796 | 0.92 | 0.93 | 0.769–1.128 | 0.47 |
| PER2 | 0.98 | 0.768–1.248 | 0.86 | 1.61 | 0.77–3.368 | 0.21 | 0.96 | 0.736–1.241 | 0.73 |
| PER3 | 1.00 | 0.849–1.176 | 0.99 | 0.80 | 0.456–1.391 | 0.42 | 1.02 | 0.856–1.217 | 0.82 |
| TEF | 0.78 | 0.652–0.93 | 0.01 | 0.97 | 0.594–1.568 | 0.89 | 0.80 | 0.654–0.966 | 0.02 |
| DBP | 0.86 | 0.720–1.03 | 0.1 | 0.84 | 0.486–1.46 | 0.54 | 0.88 | 0.719–1.077 | 0.21 |
| TIMELESS | 1.16 | 0.932–1.445 | 0.19 | 3.05 | 1.36–6.846 | 0.01 | 1.14 | 0.895–1.439 | 0.30 |
| RORA | 0.94 | 0.759–1.167 | 0.58 | 2.29 | 1.195–4.37 | 0.01 | 0.88 | 0.698–1.118 | 0.30 |
| RORB | 1.11 | 0.692–1.793 | 0.66 | 2.23 | 0.199–24.88 | 0.52 | 1.20 | 0.734–1.948 | 0.47 |
| RORC | 0.95 | 0.828–1.097 | 0.51 | 0.98 | 0.633–1.521 | 0.93 | 0.97 | 0.832–1.12 | 0.64 |
| NR1D1 | 0.79 | 0.637–0.97 | 0.03 | 0.30 | 0.134–0.647 | 0.00 | 0.86 | 0.686–1.075 | 0.18 |
| NR1D2 | 0.79 | 0.681–0.91 | 0.00 | 0.95 | 0.646–1.389 | 0.78 | 0.78 | 0.665–0.921 | 0.00 |
| CSNK1E | 1.17 | 0.922–1.493 | 0.19 | 0.62 | 0.311–1.244 | 0.18 | 1.26 | 0.97–1.636 | 0.08 |
| CSNK1A1 | 0.81 | 0.652–0.995 | 0.05 | 1.17 | 0.701–1.953 | 0.55 | 0.76 | 0.598–0.964 | 0.02 |
| HLF | 1.05 | 0.826–1.336 | 0.69 | 0.93 | 0.42–2.076 | 0.87 | 1.04 | 0.804–1.346 | 0.77 |
| BHLHE40 | 0.92 | 0.785–1.085 | 0.33 | 1.05 | 0.698–1.569 | 0.83 | 0.91 | 0.764–1.087 | 0.30 |
| BHLHE41 | 0.96 | 0.873–1.058 | 0.42 | 0.89 | 0.651–1.207 | 0.44 | 0.98 | 0.884–1.085 | 0.69 |
| Primary | Metastasis | |||
|---|---|---|---|---|
| Circadian Genes | Correlation Coefficient (r) p | Correlation Coeffi-cient (r) p | ||
| ARNTL | CTLA4 IFNG NFKB1 TNF | 0.20 0.043 0.21 0.032 0.61 0.000 0.20 0.044 | CTLA4 CXCL1 NFKB1 IFNG IL6 PDCD1 TNF | 0.35 0.0000 0.23 0.0000 0.43 0.0000 0.34 0.0000 0.24 0.0000 0.25 0.0000 0.30 0.0000 |
| BHLHE40 | CXCL1 IL6 NFKB1 | 0.17 0.087 0.39 0.000 0.45 0.000 | CXCL1 IL6 NFKB1 | 0.24 0.0000 0.26 0.0000 0.28 0.0000 |
| BHLHE41 | NFKB1 | 0.34 0.001 | CTLA4 CXCL1 IFNG NFKB1 PDCD1 TNF | 0.14 0.0084 0.23 0.0000 0.12 0.0271 0.15 0.0058 0.11 0.0321 0.15 0.0049 |
| CLOCK | NFKB1 IL6 | 0.61 0.000 0.22 0.022 | NFKB1 | 0.61 0.0000 |
| CRY1 | NFKB1 IL6 | 0.50 0.000 0.20 0.047 | NFKB1 | 0.38 0.0000 |
| CRY2 | NFKB1 | 0.36 0.000 | NFKB1 TNF | 0.23 0.0000 0.14 0.0066 |
| CSNK1A1 | NFKB1 IL6 | 0.55 0.000 0.25 0.012 | IFNG NFKB1 | 0.10 0.0734 0.40 0.0000 |
| CSNK1A1L | CXCL1 IL6 NFKB1 | 0.19 0.053 0.34 0.000 0.49 0.000 | NFKB1 | 0.30 0.0000 |
| CSNK1E | IL6 NFKB1 | 0.21 0.034 0.21 0.032 | NFKB1 | 0.14 0.0108 |
| DBP | PDCD1 | 0.18 0.062 | PDCD1 TNF | 0.20 0.0001 0.11 0.0464 |
| HLF | IFNG PDCD1 TNF | 0.19 0.058 0.18 0.069 0.20 0.048 | ||
| NPAS2 | CTLA4 NFKB1 | 0.34 0.001 0.27 0.006 | CTLA4 CXCL1 | 0.20 0.0001 0.22 0.0000 |
| NR1D2 | NFKB1 | 0.58 0.000 | IFNG NFKB1 | 0.10 0.0610 0.45 0.0000 |
| PER2 | IL6 NFKB1 TNF | 0.19 0.050 0.33 0.001 0.20 0.046 | NFKB1 TNF | 0.34 0.0000 0.10 0.0592 |
| PER3 | NFKB1 | 0.28 0.005 | NFKB1 | 0.27 0.0000 |
| RORA | NFKB1 TNF | 0.33 0.001 0.24 0.013 | CTLA4 IFNG IL6 NFKB1 PDCD1 TNF | 0.26 0.0000 0.27 0.0000 0.31 0.0000 0.40 0.0000 0.20 0.0002 0.28 0.0000 |
| RORC | IFNG NFKB1 PDCD1 TNF | 0.23 0.022 0.25 0.010 0.22 0.025 0.32 0.001 | CTLA4 CXCL1 IFNG IL6 NFKB1 PDCD1 TNF | 0.35 0.0000 0.13 0.0147 0.28 0.0000 0.18 0.0008 0.13 0.0175 0.29 0.0000 0.39 0.0000 |
| RORB | CXCL1 IFNG PDCD1 | 0.10 0.0649 0.14 0.0069 0.10 0.0634 | ||
| TEF | IL6 NFKB1 | 0.19 0.060 0.43 0.000 | NFKB1 | 0.29 0.0000 |
| TIMELESS | CXCL1 IL6 NFKB1 | 0.18 0.064 0.18 0.062 0.39 0.000 | NFKB1 | 0.23 0.0000 |
| B Cell | CD4+ T Cells | CD8+ T Cells | Dendritic Cells | Macrophages | Neutrophils | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Correlation | p Value | Correlation | p Value | Correlation | p Value | Correlation | p Value | Correlation | p Value | Correlation | p Value | |
| ARNTL/ BMAL1 | 0.114 | 0.016 | 0.176 | 0.000 | 0.394 | 0.000 | 0.291 | 0.000 | 0.283 | 0.000 | 0.509 | 0.000 |
| BHLHE40 | 0.095 | 0.045 | 0.053 | 0.266 | 0.140 | 0.003 | 0.137 | 0.004 | 0.121 | 0.010 | 0.240 | 0.000 |
| BHLHE41 | 0.067 | 0.154 | −0.029 | 0.548 | 0.104 | 0.030 | 0.101 | 0.033 | 0.025 | 0.594 | 0.096 | 0.041 |
| CLOCK | 0.095 | 0.044 | 0.119 | 0.012 | 0.359 | 0.000 | 0.214 | 0.000 | 0.272 | 0.000 | 0.452 | 0.000 |
| CRY1 | 0.124 | 0.009 | 0.080 | 0.092 | 0.276 | 0.000 | 0.154 | 0.001 | 0.253 | 0.000 | 0.343 | 0.000 |
| CRY2 | 0.138 | 0.003 | 0.372 | 0.000 | 0.170 | 0.000 | 0.207 | 0.000 | 0.257 | 0.000 | 0.257 | 0.000 |
| CSNK1A1 | −0.020 | 0.678 | 0.125 | 0.008 | 0.305 | 0.000 | 0.116 | 0.014 | 0.264 | 0.000 | 0.449 | 0.000 |
| CSNK1A1LL | −0.026 | 0.587 | 0.059 | 0.213 | 0.170 | 0.000 | 0.074 | 0.120 | 0.148 | 0.002 | 0.280 | 0.000 |
| CSNK1E | −0.039 | 0.405 | 0.058 | 0.220 | −0.091 | 0.057 | −0.046 | 0.330 | 0.052 | 0.271 | −0.028 | 0.552 |
| DBP | 0.048 | 0.310 | 0.309 | 0.000 | −0.022 | 0.649 | 0.136 | 0.004 | −0.009 | 0.849 | 0.028 | 0.552 |
| HLF | −0.045 | 0.343 | 0.161 | 0.001 | 0.064 | 0.182 | −0.022 | 0.642 | 0.054 | 0.252 | 0.065 | 0.166 |
| NFIL3 | 0.068 | 0.151 | −0.009 | 0.851 | 0.200 | 0.000 | 0.099 | 0.038 | 0.222 | 0.000 | 0.305 | 0.000 |
| NPAS2 | 0.007 | 0.880 | −0.093 | 0.049 | 0.130 | 0.007 | 0.046 | 0.335 | −0.036 | 0.449 | 0.088 | 0.062 |
| NR1D1 | −0.052 | 0.272 | 0.059 | 0.211 | −0.046 | 0.332 | −0.029 | 0.546 | −0.094 | 0.045 | −0.036 | 0.450 |
| NR1D2 | 0.124 | 0.008 | 0.187 | 0.000 | 0.398 | 0.000 | 0.226 | 0.000 | 0.289 | 0.000 | 0.520 | 0.000 |
| PER1 | 0.000 | 0.998 | 0.134 | 0.005 | −0.007 | 0.880 | −0.089 | 0.060 | 0.055 | 0.245 | 0.007 | 0.885 |
| PER2 | 0.122 | 0.010 | 0.237 | 0.000 | 0.157 | 0.001 | 0.143 | 0.002 | 0.219 | 0.000 | 0.264 | 0.000 |
| PER3 | 0.087 | 0.065 | 0.172 | 0.000 | 0.253 | 0.000 | 0.156 | 0.001 | 0.110 | 0.019 | 0.209 | 0.000 |
| RORA | 0.071 | 0.135 | 0.178 | 0.000 | 0.303 | 0.000 | 0.159 | 0.001 | 0.288 | 0.000 | 0.445 | 0.000 |
| RORB | 0.051 | 0.283 | −0.020 | 0.674 | 0.147 | 0.002 | 0.015 | 0.749 | −0.044 | 0.355 | 0.104 | 0.027 |
| RORC | 0.138 | 0.003 | 0.023 | 0.628 | 0.194 | 0.000 | 0.080 | 0.090 | 0.015 | 0.749 | 0.130 | 0.006 |
| TEF | 0.080 | 0.091 | 0.240 | 0.000 | 0.091 | 0.057 | 0.103 | 0.029 | 0.161 | 0.001 | 0.184 | 0.000 |
| TIMELESS | 0.148 | 0.002 | 0.059 | 0.215 | 0.149 | 0.002 | 0.165 | 0.000 | 0.091 | 0.052 | 0.198 | 0.000 |
| Gene | Number of Mutated Samples | % of Mutated Samples | Place of Mutated Samples Among Other Cancer Types |
|---|---|---|---|
| PER3 | 37 | 8% | 2 |
| PER2 | 36 | 8% | 2 |
| RORC | 34 | 8% | 1 |
| RORB | 33 | 7% | 1 |
| TIMELESS | 25 | 6% | 2 |
| TEF | 22 | 5% | 3 |
| PER1 | 21 | 5% | 2 |
| NPAS2 | 20 | 5% | 1 |
| CLOCK | 18 | 4% | 4 |
| NR1D2 | 18 | 4% | 2 |
| CRY2 | 17 | 4% | 2 |
| CSNK1E | 17 | 4% | 10 |
| NR1D1 | 16 | 4% | 1 |
| BMAL1 | 15 | 3% | 2 |
| CRY1 | 14 | 3% | 3 |
| HLF | 14 | 3% | 1 |
| RORA | 13 | 3% | 2 |
| CSNK1A1 | 13 | 3% | 3 |
| CIPC | 12 | 3% | 2 |
| BHLHE40 | 10 | 2% | 2 |
| CSNK1A1L | 10 | 2% | 5 |
| DBP | 5 | 1% | 4 |
| NFIL3 | 5 | 1% | 8 |
| BHLHE41 | 4 | <1% | 14 |
| CpG Island | CpG Site | Impact of Methylated CpG to mRNA Expression | Correlation Coefficient (r), p |
|---|---|---|---|
| CLOCK (cg05960024) | 5′UTR | Not significant | |
| ARNTL/BMAL1 (cg13250711) | TSS1500 | Not significant | |
| ARNTL/BMAL1 (cg06499652) | 1ST EXON 5′UTR | Not significant | |
| BHLHE40 (cg08587820) | TSS200 | Significant | r = −0.12 p = 0.01 |
| BHLHE40 (cg03223878) | 1ST EXON 5′UTR | Not significant | |
| BHLHE41 (cg03046445) | 1ST EXON 5′UTR | Not significant | |
| BHLHE41 (cg19243777) | TSS1500 | Not significant | |
| PER1 (cg16545079) | TSS200 | Not significant | |
| PER2 (cg05664072) | TSS1500 | Not significant | |
| PER2 (cg23905308) | 5′UTR | Significant | r = −0.26 p < 0.001 |
| PER3 (cg06487986) | TSS1500 | Not significant | |
| PER3 (cg25514503) | BODY | Significant | r = −0.53 p < 0.001 |
| CRY1 (cg03742214) | 1ST EXON 5′UTR | Significant | r = −0.11 p = 0.02 |
| CRY1 (cg10126874) | 1ST EXON 5′UTR | Significant | r = −0.17 p < 0.01 |
| CRY2 (cg01618083) | BODY 1ST EXON | Not significant | r = −0.18 p < 0.001 |
| NPAS2 (cg07799947) | TSS1500 | Not significant | |
| NPAS2 (cg14383135) | 5′UTR | Significant | r = −0.13 p < 0.001 |
| NR1D1 (cg02337166) | TSS1500 | Significant | r = −0.16 p = 0.009 |
| NR1D1 (cg22640452) | 1ST EXON 5′UTR | Not significant | |
| NR1D2 (cg14452875 | Body TSS1500 | Not significant | |
| TEF (cg04137037) | Body TSS200 | Not significant | |
| HLF (cg04219321) | TSS1500 | Significant | r = 0.32 p < 0.001 |
| RORA (cg13301933) | 1ST EXON 5′UTR | Not significant | |
| RORA (cg27167601) | TSS1500 | Significant | r = 0.23 p < 0.001 |
| RORB (cg07536920) | 1ST EXON 5′UTR | Not significant | |
| RORB (cg14331163) | TSS1500 | Significant | r = −0.13 p = 0.06 |
| RORC (cg18149207) | TSS1500 | Significant | r = −0.54 p < 0.001 |
| RORC (cg25112191) | 1ST EXON 5′UTR | Significant | |
| TIMELESS (cg00819696) | 5′UTR | Not significant | |
| TIMELESS (cg22580905) | TSS200 | Not significant | |
| CSNK1E (cg01346718) | TSS1500 5′UTR | Significant | r = −0.34 p < 0.001 |
| CSNK1E (cg01441777) | TSS1500 | Significant | r = −0.39 p < 0.001 |
| CSNK1A1 (cg05607472) | BODY | Not significant | |
| CSNK1A1 (cg06899802) | TSS200 | Not significant | |
| CSNK1A1L (cg01668383) | 1ST EXON 5′UTR | Not significant | |
| NFIL3 (cg10708828) | 5′UTR | Not significant | |
| NFIL3 (cg15919045) | TSS1500 | Not significant |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lesicka, M.; Nedoszytko, B.; Reszka, E. Disruptions of Circadian Genes in Cutaneous Melanoma—An In Silico Analysis of Transcriptome Databases. Int. J. Mol. Sci. 2023, 24, 10140. https://doi.org/10.3390/ijms241210140
Lesicka M, Nedoszytko B, Reszka E. Disruptions of Circadian Genes in Cutaneous Melanoma—An In Silico Analysis of Transcriptome Databases. International Journal of Molecular Sciences. 2023; 24(12):10140. https://doi.org/10.3390/ijms241210140
Chicago/Turabian StyleLesicka, Monika, Bogusław Nedoszytko, and Edyta Reszka. 2023. "Disruptions of Circadian Genes in Cutaneous Melanoma—An In Silico Analysis of Transcriptome Databases" International Journal of Molecular Sciences 24, no. 12: 10140. https://doi.org/10.3390/ijms241210140