
Saline-Alkali Soil Property Improved by the Synergistic Effects of Priestia aryabhattai JL-5, Staphylococcus pseudoxylosus XW-4, Leymus chinensis and Soil Microbiota

 and
and
Abstract
:1. Introduction
2. Results
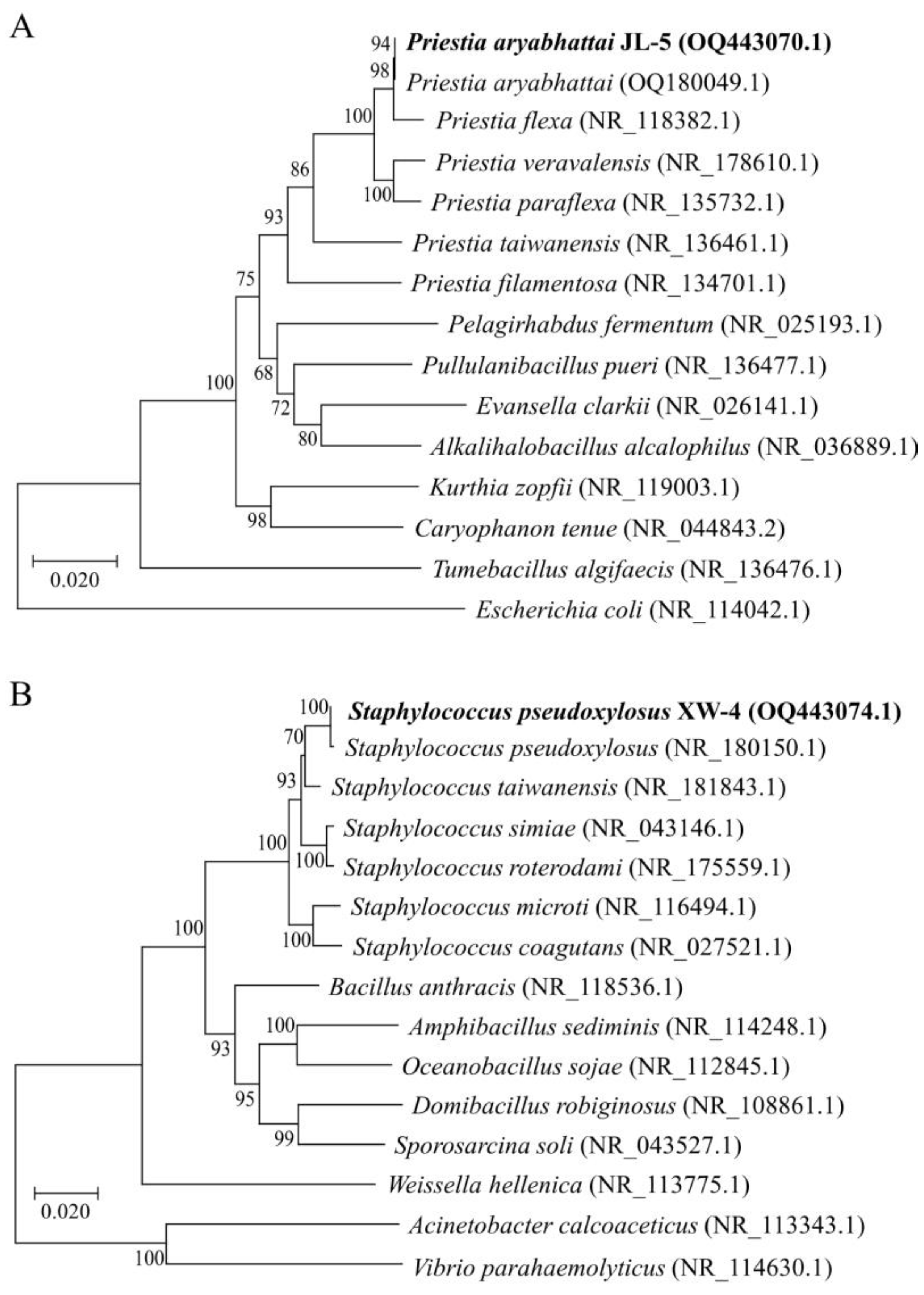
2.1. Isolation and Identification of Saline-alkali-Tolerant Bacteria
2.2. Transcriptome Sequencing and Data Analysis
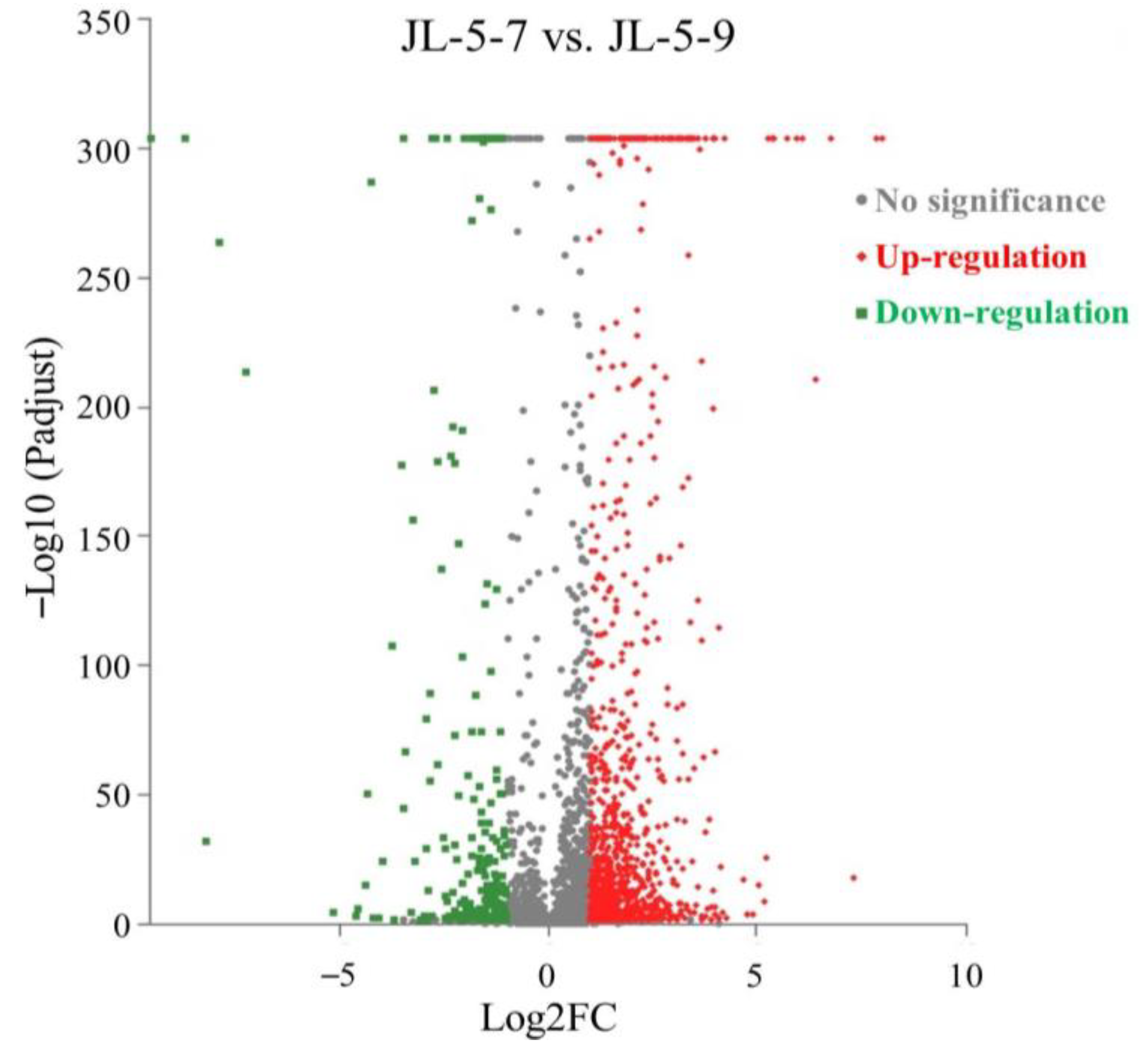
2.2.1. Selection of High-Quality Transcriptome Sequences
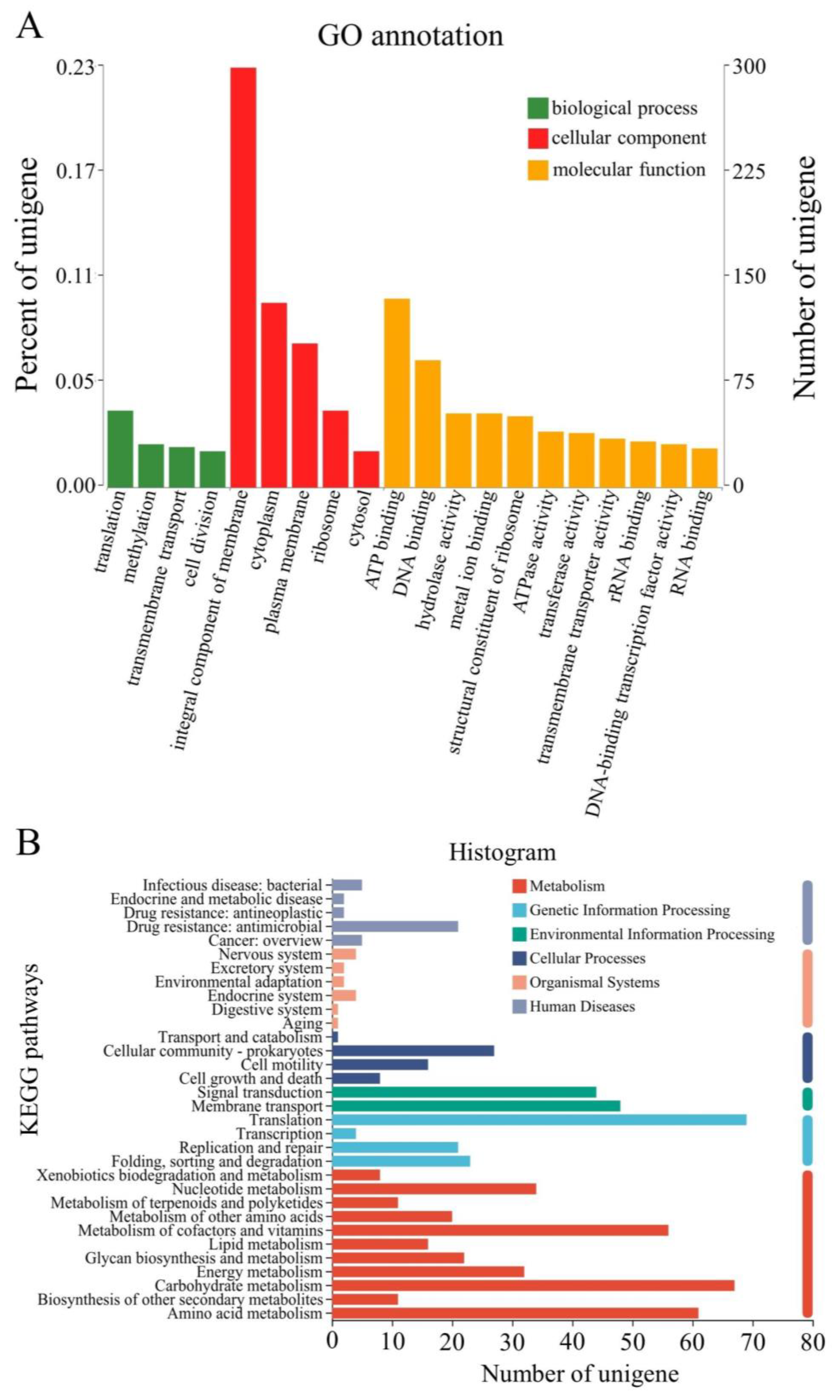
2.2.2. GO Annotation and KEGG Enrichment Analyses of Differentially Expressed Genes
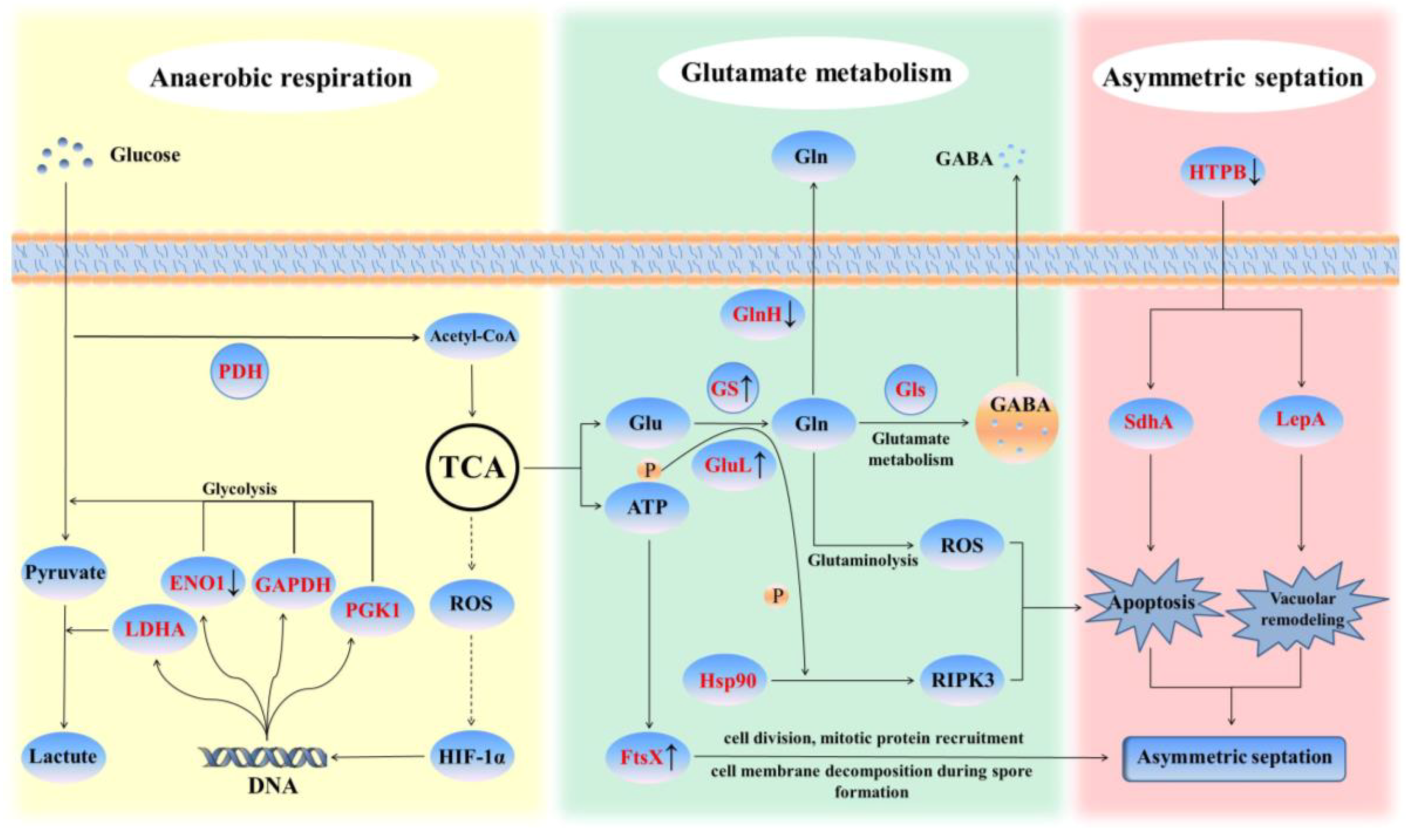
2.2.3. Glutamate Metabolism in Response to Saline-alkali Stress in Priestia aryabhattai JL-5
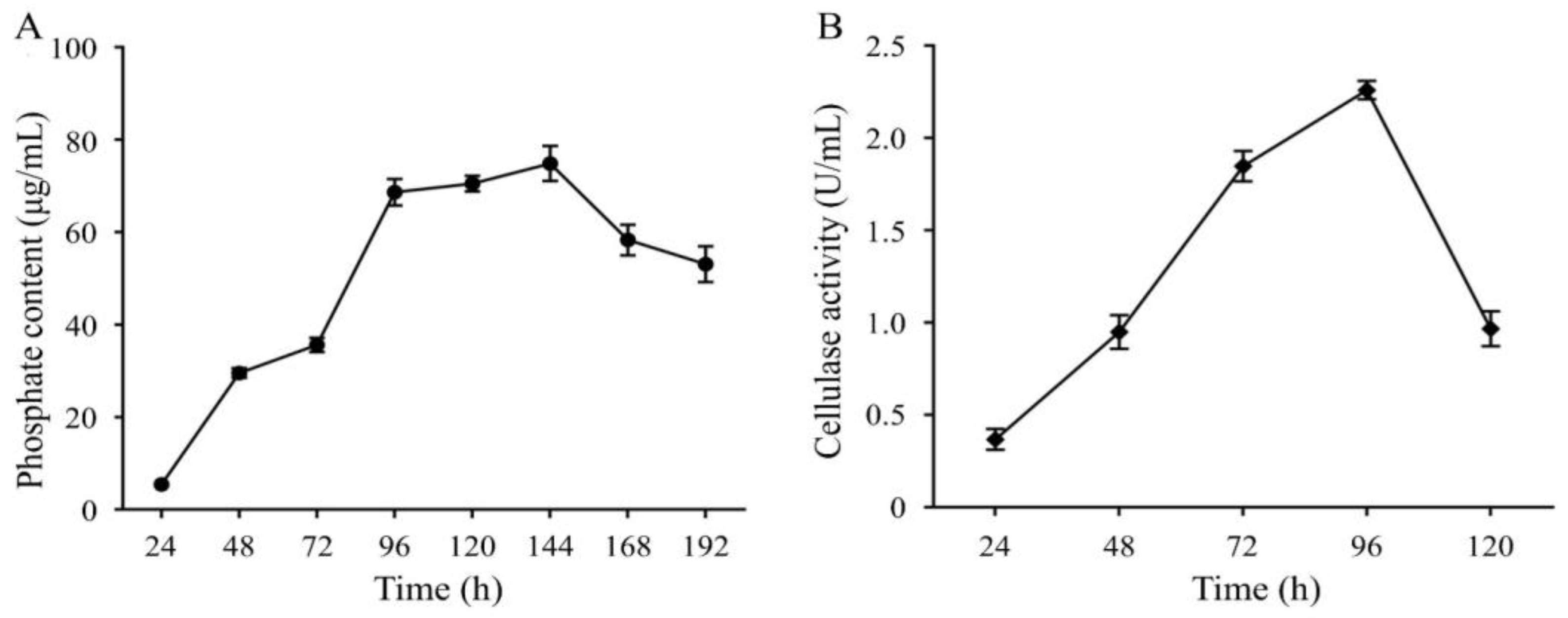
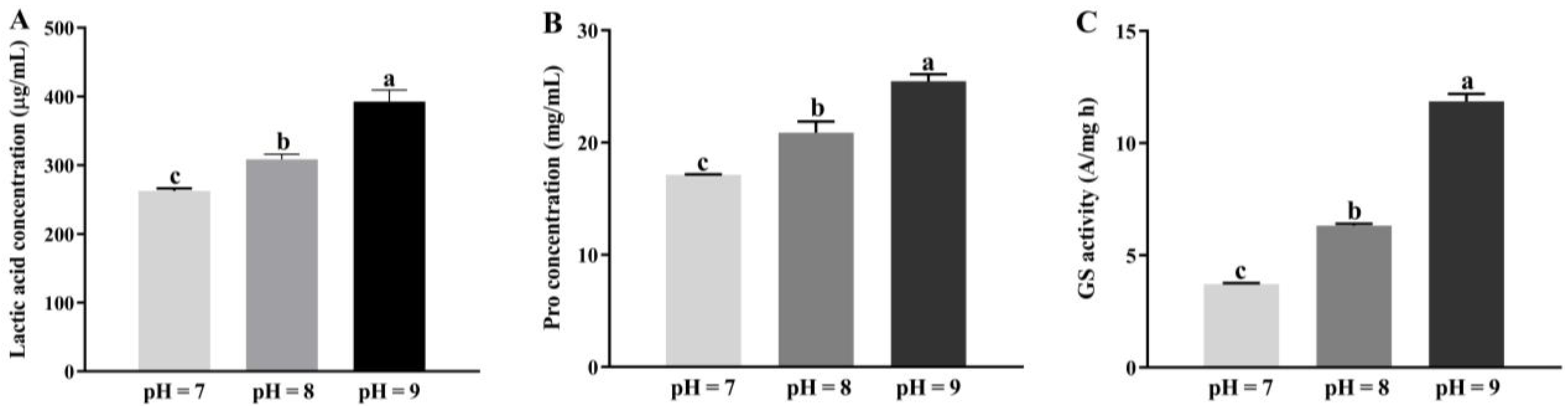
2.2.4. Effect of Saline-alkali Stress on Priestia aryabhattai JL-5
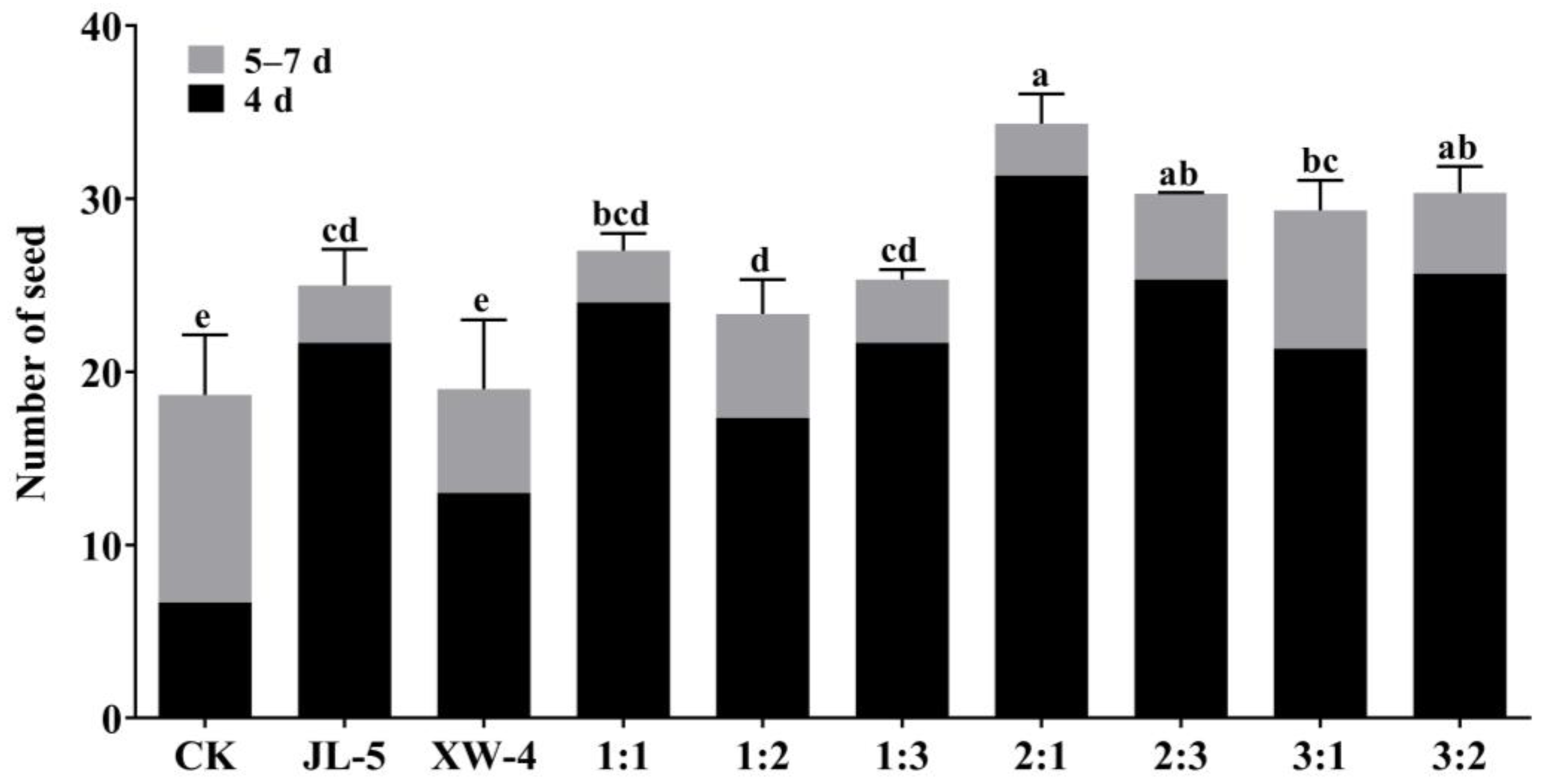
2.3. Optimal Ratio of the Strains JL-5 and XW-4 in the Bacterial Solutions and Their Effects on the Growth of Leymus chinensis
2.4. Effects of Bacterial Solutions on the Soil Nutrients and Microbial Community
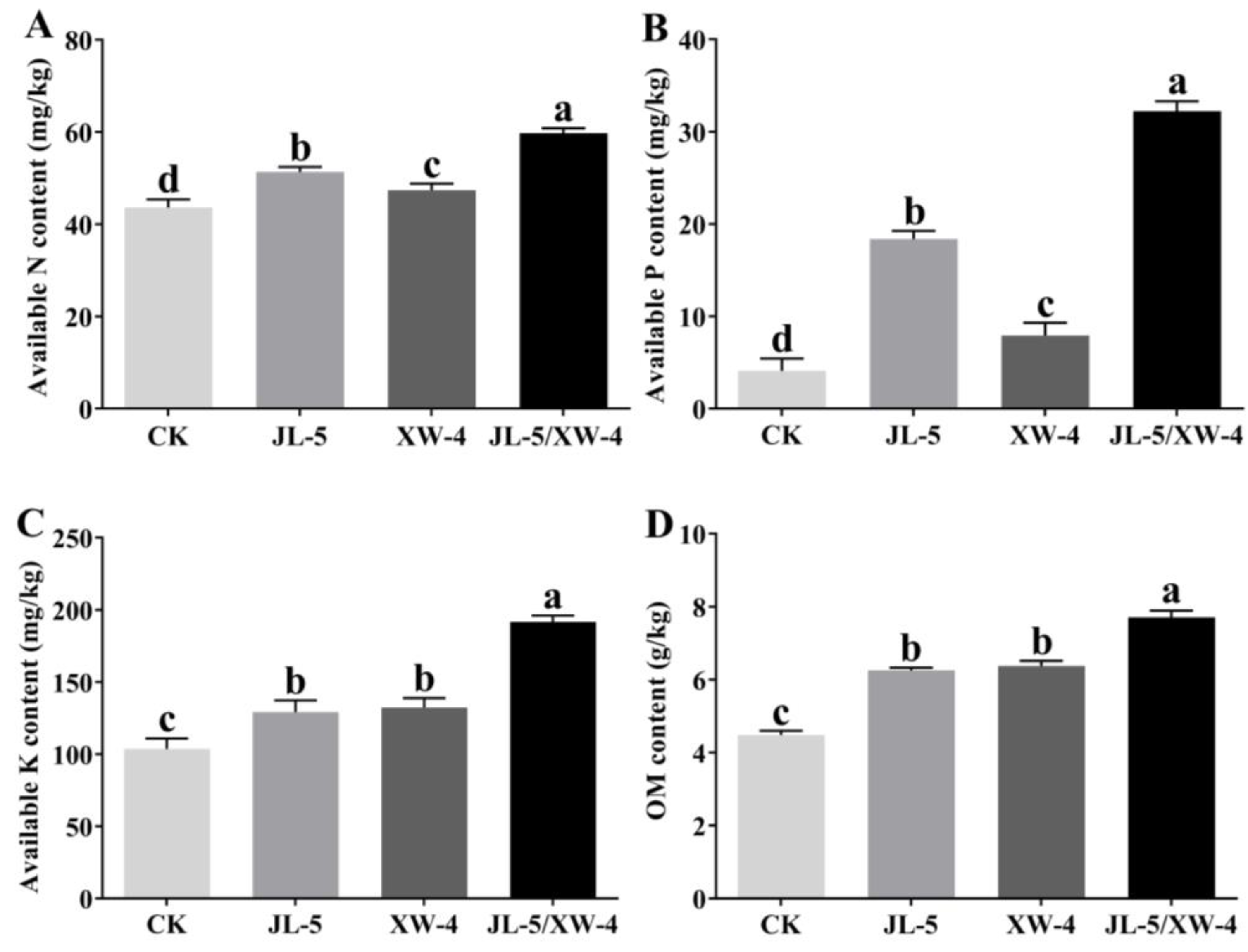
2.4.1. Effects of Bacterial Solutions on Available Nitrogen, Phosphorus and Potassium and Organic Matter in Soil
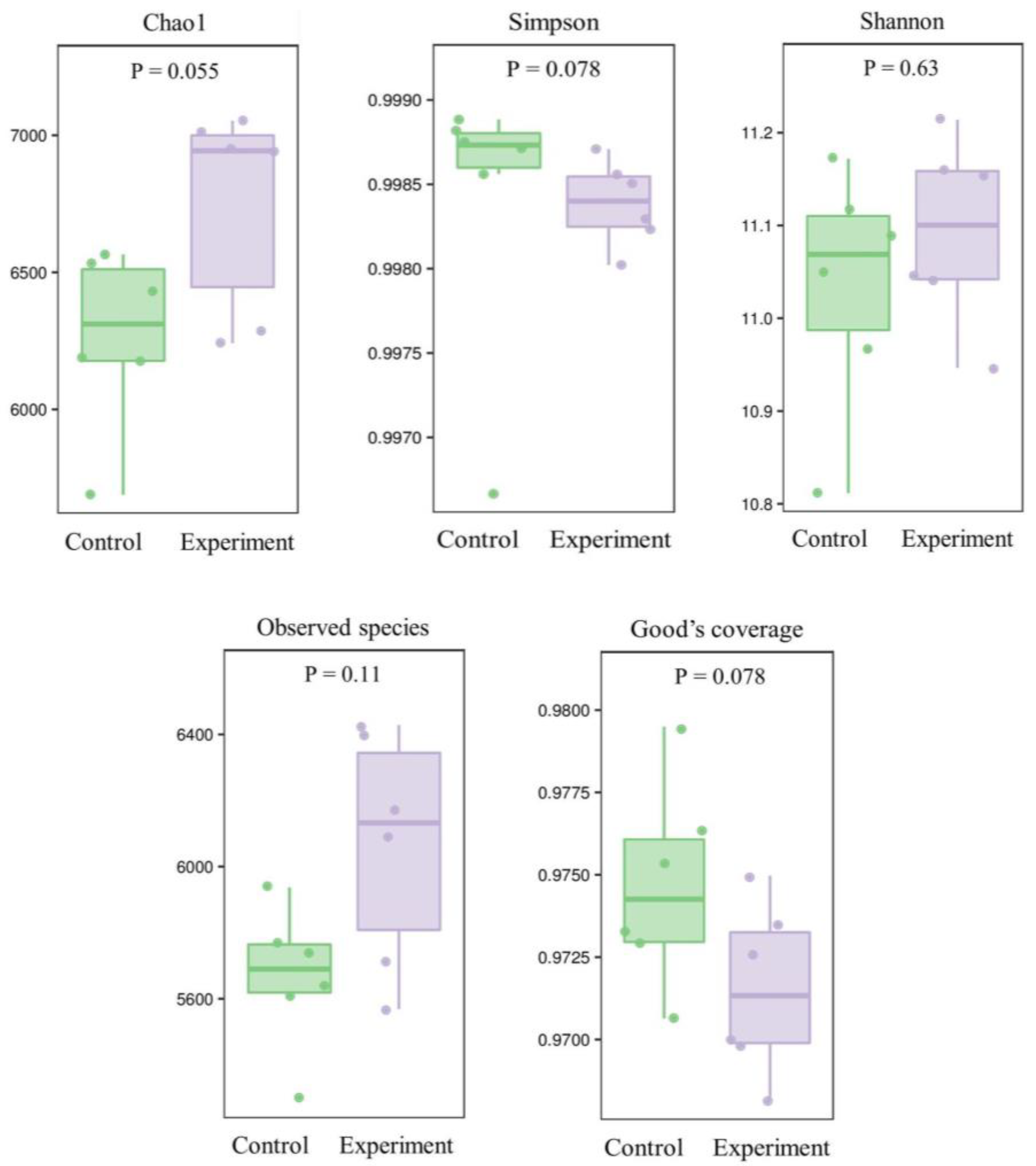
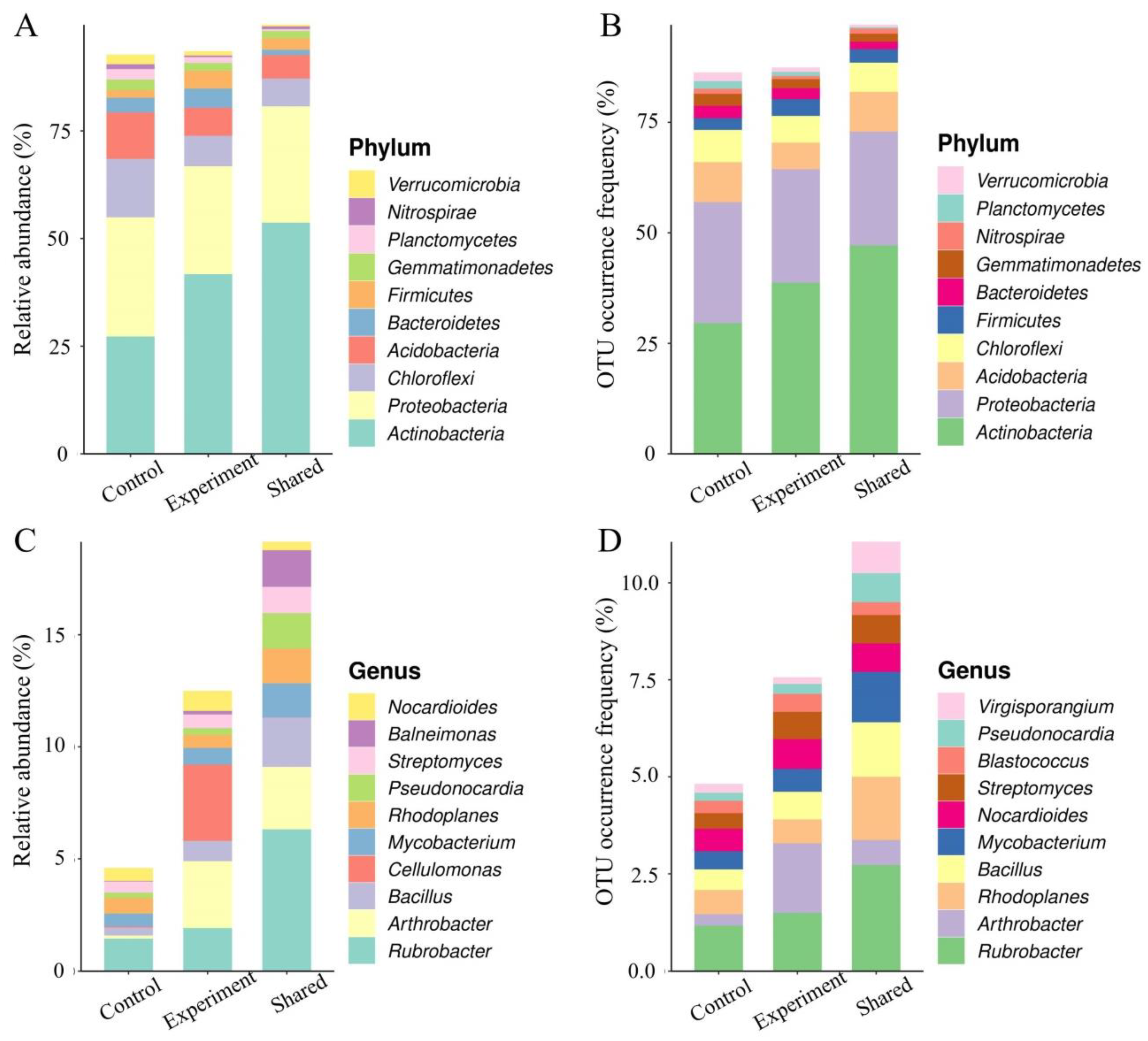
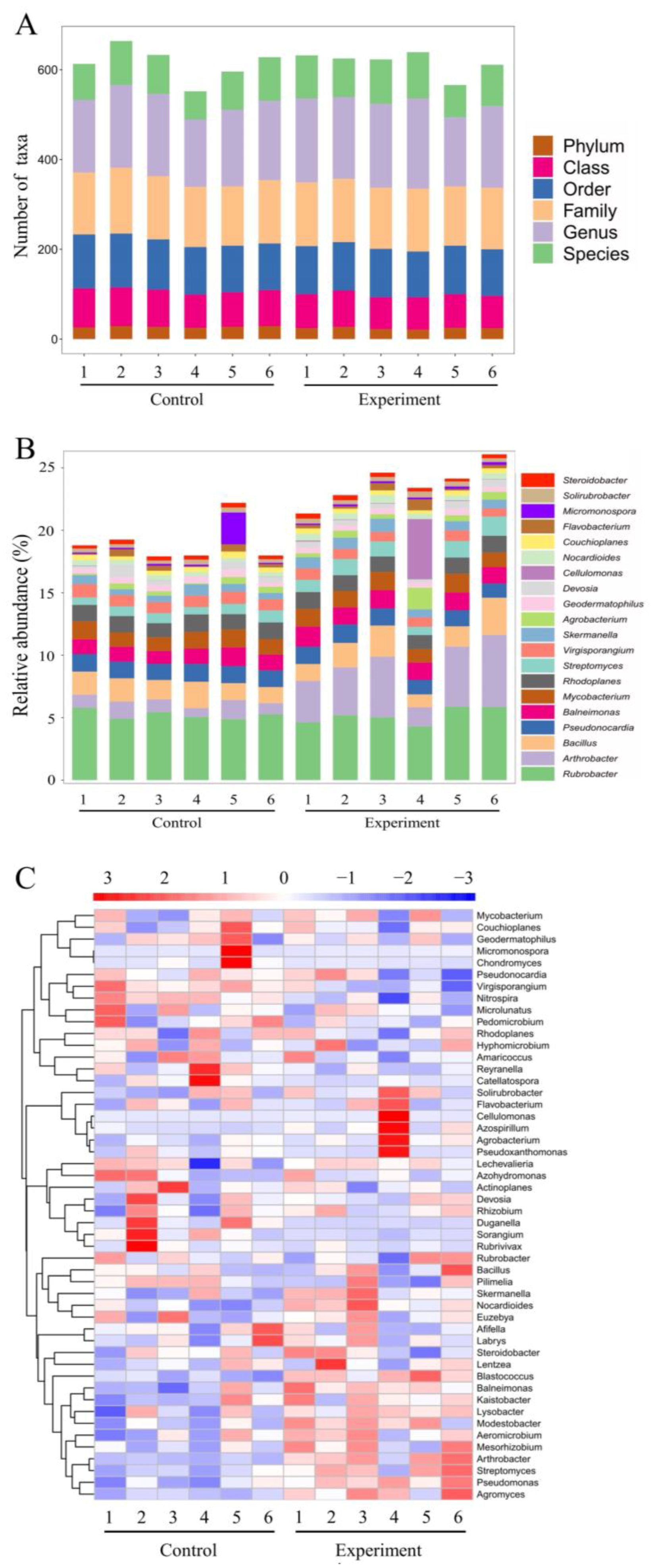
2.4.2. Effects of the Combined Bacterial Solution on Soil Microbial Diversity
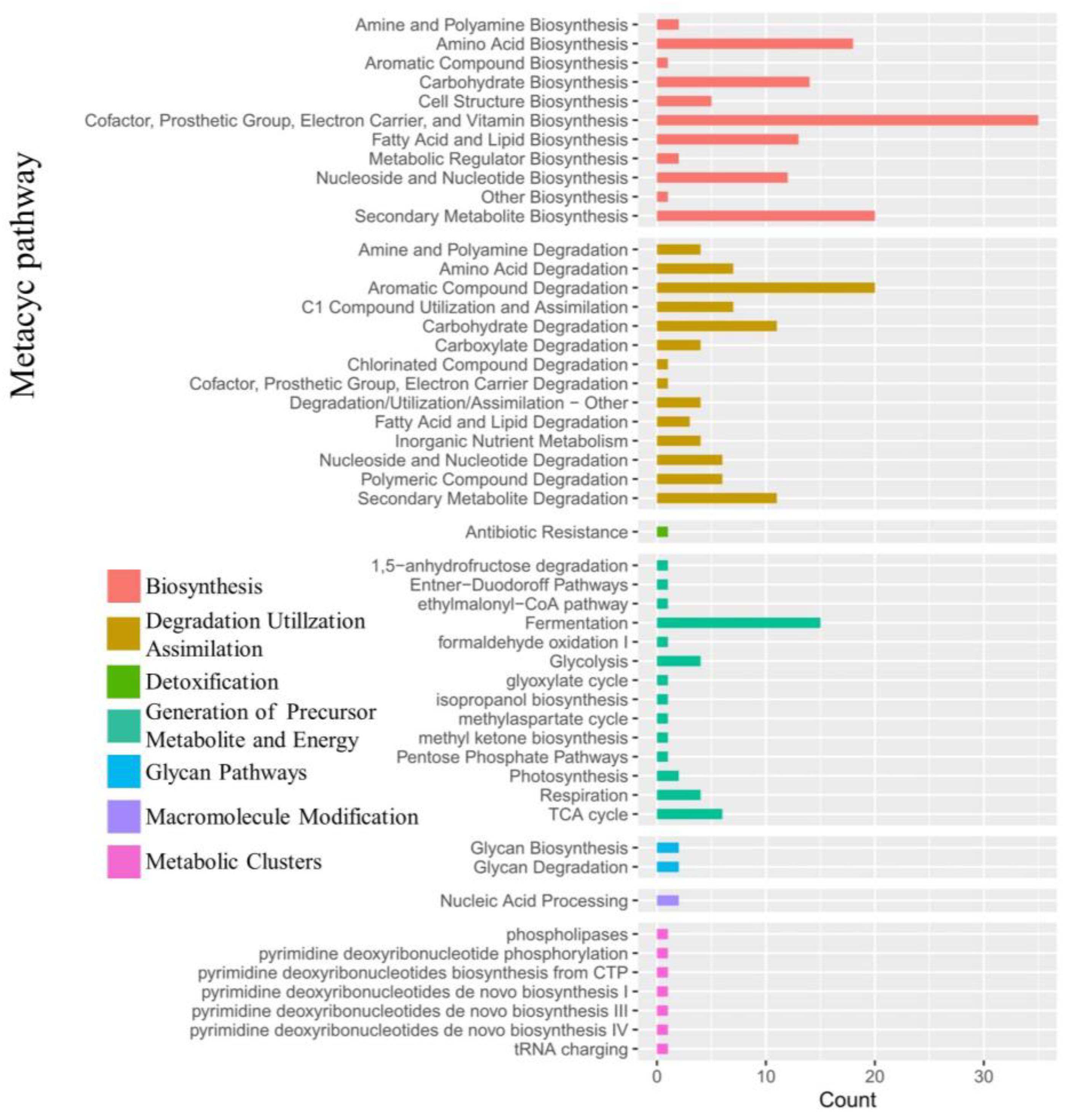
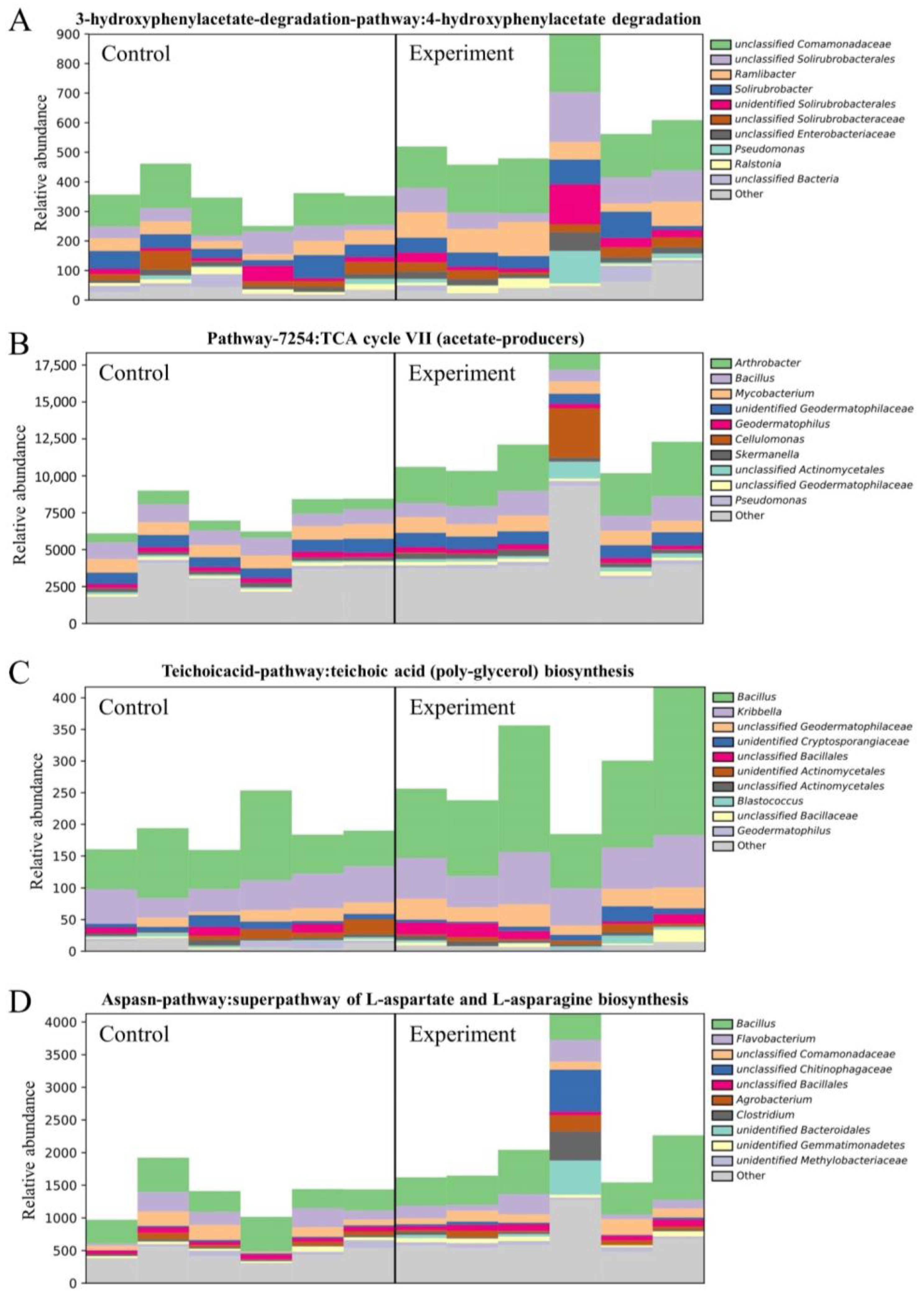
2.4.3. Effects of the Combined Bacterial Solution on Soil Microbial Metabolic Pathways
3. Discussion
3.1. Alleviated Saline-alkali Stress by Priestia aryabhattai JL-5 via Glutamate Metabolism
3.2. Physiological Variations in Priestia aryabhattai JL-5 in Response to Saline-alkali Stress
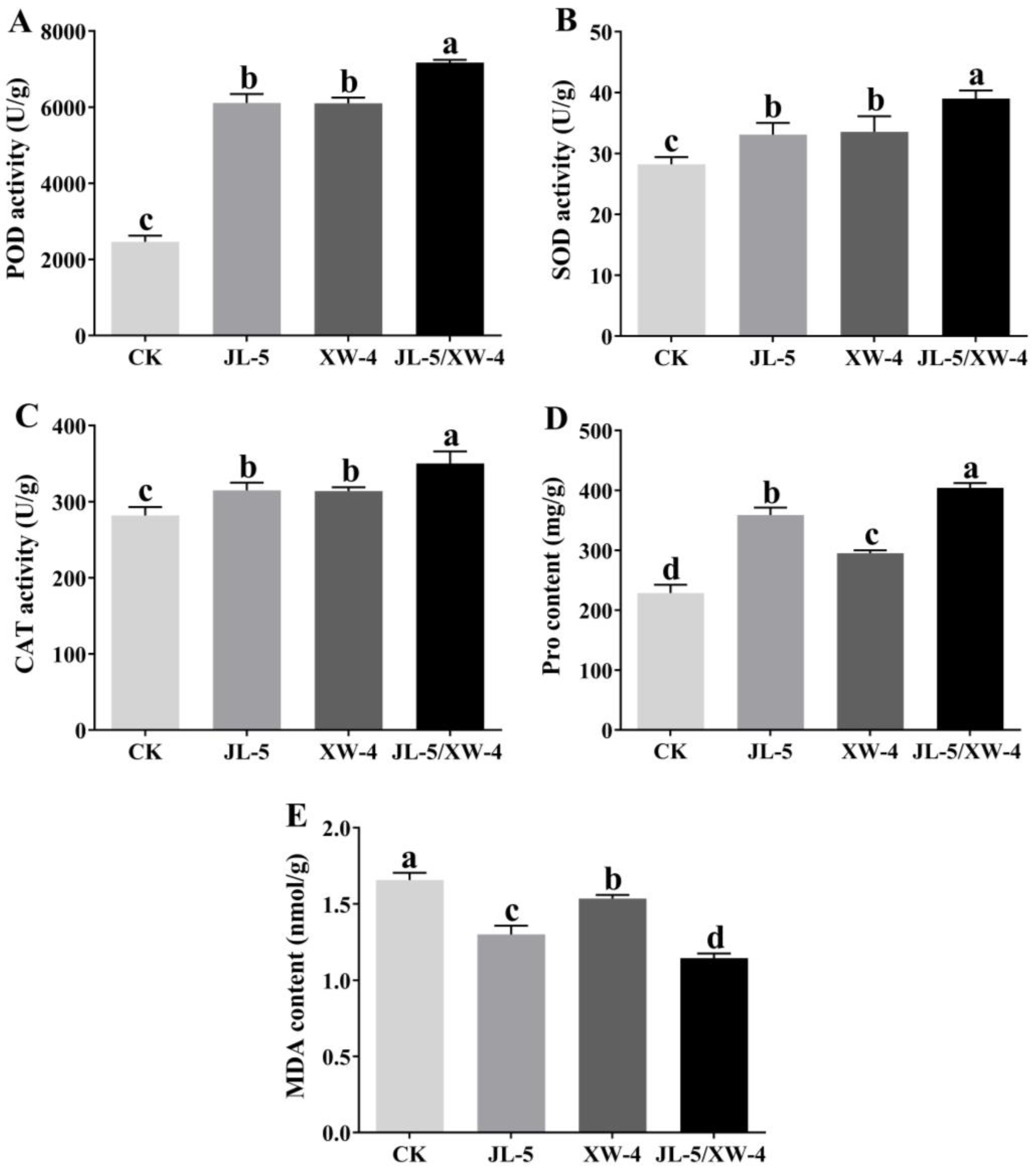
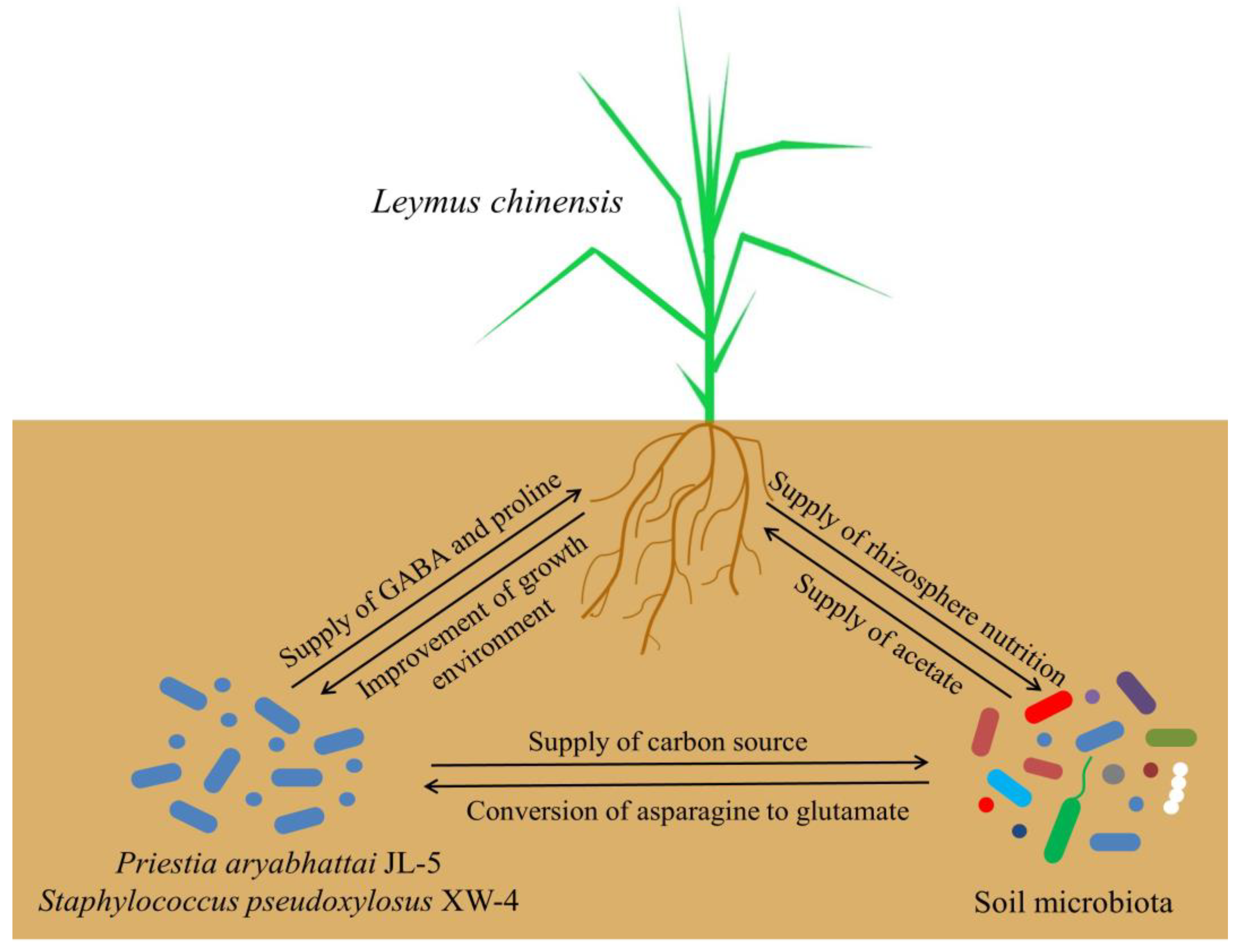
3.3. Improved Saline-alkali Soils by Combined Bacterial Solutions Promoting the Interaction of Leymus chinensis and Other Microorganisms
4. Materials and Methods
4.1. Isolation and Identification of Saline-alkali-Tolerant Bacteria
4.1.1. Isolation and Screening of Saline-alkali-Tolerant Bacteria
4.1.2. Functional Selection and Identification of Saline-alkali-Tolerant Bacteria
4.2. Transcriptome Sequencing and Analysis
4.2.1. Bacterial Sample Preparation
4.2.2. Analysis and Annotation of Differentially Expressed Genes
4.2.3. Determination of Biochemical, Physiological and Morphological Characteristics of Bacteria
4.3. Promoted Growth and Alleviated Saline-alkali Stress of Leymus chinensis by the Strains JL-5 and XW-4
4.4. Evaluation of Soil Properties and Microbiota
4.5. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Huang, J.; Zhu, C.; Kong, Y.; Cao, X.; Zhu, L.; Zhang, J. Biochar application alleviated rice salt stress via modifying soil properties and regulating soil bacterial abundance and community structure. Agronomy 2022, 12, 409. [Google Scholar] [CrossRef]
- Patel, B.B.; Dave, R.S. Studies on infiltration of saline-alkali soils of several parts of Mehsana and Patan districts of North Gujarat. J. Appl. Technol. Environ. Sani 2011, 1, 87–92. [Google Scholar]
- Zhang, H.; Yu, F.; Xie, P.; Sun, S.; Qiao, X.; Tang, S.; Chen, C.; Yang, S.; Mei, C.; Yang, D.; et al. A Gγ protein regulates alkaline sensitivity in crops. Science 2023, 379, e8416. [Google Scholar] [CrossRef]
- Ibarra-Villarreal, A.L.; Gándara-Ledezma, A.; Godoy-Flores, A.D.; Herrera-Sepúlveda, A.; Díaz-Rodríguez, A.M.; Parra-Cota, F.I.; De los Santos-Villalobos, S. Salt-tolerant Bacillus species as a promising strategy to mitigate the salinity stress in wheat (Triticum turgidum subsp. durum). J. Arid. Environ. 2021, 186, 104399. [Google Scholar] [CrossRef]
- Spadaro, D.; Gullino, M.L. Improving the efficiency of biocontrol agents against soilborne pathogens. Crop Protection 2005, 24, 601–613. [Google Scholar] [CrossRef]
- Li, K.; Li, Q.; Geng, Y.; Liu, C. An evaluation of the effects of microstructural characteristics and frost heave on the remediation of saline-alkali soils in the Yellow River Delta, China. Land Degrad. Dev. 2021, 32, 1325–1337. [Google Scholar] [CrossRef]
- Razeghi, H.R.; Ghadir, P.; Javadi, A.A. Mechanical strength of saline sandy soils stabilized with alkali-activated cements. Sustainability 2022, 14, 13669. [Google Scholar] [CrossRef]
- Alkharabsheh, H.M.; Seleiman, M.F.; Battaglia, M.L.; Shami, A.; Jalal, R.S.; Alhammad, B.A. Biochar and its broad impacts in soil quality and fertility, nutrient leaching and crop productivity: A review. Agronomy 2021, 11, 993. [Google Scholar] [CrossRef]
- Zhao, W.; Zhou, Q.; Tian, Z.; Cui, Y.; Liang, Y.; Wang, H. Apply biochar to ameliorate soda saline-alkali land, improve soil function and increase corn nutrient availability in the Songnen Plain. Sci. Total Environ. 2020, 722, 137428. [Google Scholar] [CrossRef] [PubMed]
- Anam, G.B.; Reddy, M.S.; Ahn, Y.H. Characterization of Trichoderma asperellum RM-28 for its sodic/saline-alkali tolerance and plant growth promoting activities to alleviate toxicity of red mud. Sci. Total Environ. 2019, 662, 462–469. [Google Scholar] [CrossRef]
- Wang, X.; Xu, X.; Lu, A.; Li, X.; Zhang, F. Cultivation of two barnyard variations improves physicochemical properties of saline-alkali land through mediating rhizospheric microbiome and metabolome. Agronomy 2022, 12, 1322. [Google Scholar] [CrossRef]
- Xu, Y.; Li, Y.; Long, C.; Han, L. Alleviation of salt stress and promotion of growth in peanut by Tsukamurella tyrosinosolvens and Burkholderia pyrrocinia. Biologia 2022, 9, 2423–2433. [Google Scholar] [CrossRef]
- Zhang, J.; Fan, X.; Wang, X.; Tang, Y.; Zhang, H.; Yuan, Z.; Zhou, J.; Han, Y.; Li, T. Bioremediation of a saline-alkali soil polluted with Zn using ryegrass associated with Fusarium incarnatum. Environ. Pollut. 2022, 312, 119929. [Google Scholar] [CrossRef] [PubMed]
- Hovik, P.; Anna, H.; Nils-Kåre, B.; Armen, T. Bacilli community of saline–alkaline soils from the Ararat Plain (Armenia) assessed by molecular and culture-based methods. Syst. Appl. Microbiol. 2018, 3, 232–240. [Google Scholar]
- Farghaly, F.A.; Nafady, N.A.; Abdel–Wahab, D.A. The efficiency of Arbuscular mycorrhiza in increasing tolerance of Triticum aestivum L. to alkaline stress. BMC Plant Biol. 2022, 22, 490. [Google Scholar] [CrossRef] [PubMed]
- Sánchez, E.; Zabaleta, R.; Fabani, M.P.; Rodriguez, R.; Mazza, G. Effects of the amendment with almond shell, bio-waste and almond shell-based biochar on the quality of saline-alkali soils. J. Environ. Manag. 2022, 318, 115604. [Google Scholar] [CrossRef]
- Slonczewski, J.L.; Fujisawa, M.; Dopson, M.; Krulwich, T.A. Cytotoxic pH measurement and homeostasis in bacteria and archaea. Adv. Microb. Phys. 2009, 55, 1–79. [Google Scholar]
- Sorokin, D.Y.; van Pelt, S.; Tourova, T.P.; Evtushenko, L.I. Nitriliruptor alkaliphilus gen. nov., sp. nov., a deep-lineage haloalkaliphilic actinobacterium from soda lakes capable of growth on aliphatic nitrogen, and proposal of Nitriliruptoraceae fam nov. and Nitriliruptorales ord. nov. Int. J. Syst. Evol. Microbiol. 2009, 59, 248–253. [Google Scholar] [CrossRef] [PubMed]
- Athen, S.R.; Dubey, S.; Kyndt, J.A. The eastern Nebraska salt marsh microbiome is well adapted to an alkaline and extreme saline environment. Life 2021, 11, 446. [Google Scholar] [CrossRef]
- Ahmed, V.; Verma, M.K.; Gupta, S.; Mandhan, V.; Chauhan, N.S. Metagenomic profiling of soil microbes to mine salt stress tolerance genes. Front. Microbiol. 2018, 9, 159. [Google Scholar] [CrossRef]
- Coker, J.A.; DasSarma, P.; Kumar, J.; Müller, J.A.; DasSarma, S. Transcriptional profiling of the model archaeon Halobacterium sp. NRC-1: Responses to changes in salinity and temperature. Saline Syst. 2007, 3, 6. [Google Scholar] [CrossRef] [PubMed]
- Chen, D.; Fang, B.; Manzoor, A.; Liu, Y.; Li, L.; Mohamad, O.; Shu, W.; Li, W. Reviving the salinity adaptation mechanism in halolerant bacterium Egicoccus halophilus EGI 80432T by physiological analysis and comparative transcriptomics. Appl. Microbiol. Biotechnol. 2021, 105, 2497–2511. [Google Scholar] [CrossRef] [PubMed]
- Williams, P. Quorum sensing, communication and cross-kingdom signalling in the bacterial world. Microbiology 2007, 153, 3923–3938. [Google Scholar] [CrossRef] [PubMed]
- Layeghifard, M.; Hwang, D.M.; Guttman, D.S. Disentangling interactions in the microbiome: A network perspective. Trends Microbiol. 2017, 25, 217–228. [Google Scholar] [CrossRef]
- Bever, J.D.; Platt, T.G.; Morton, E.R. Microbial population and community dynamics on plant roots and their feedbacks on plant communities. Annu. Rev. Microbiol. 2012, 66, 265–283. [Google Scholar] [CrossRef]
- Liu, Y.; Liu, X.; Dong, X.; Yan, J.; Xie, Z.; Luo, Y. The effect of Azorhizobium caulinodans ORS571 and γ-aminobutyric acid on salt tolerance of Sesbania rostrata. Front. Plant Sci. 2022, 13, 926850. [Google Scholar] [CrossRef]
- Li, Y.; Li, Q.; Liu, H.; Li, S.; Wang, Y.; Liu, Y. Ecological stoichiometry-based study of the influence of soil saline-alkali stress on nutrient homeostasis in L. chinensis. Ecotoxicol. Environ. Saf. 2018, 165, 243–249. [Google Scholar] [CrossRef]
- Liu, B.; Kang, C.; Wang, X.; Bao, G. Physiological and morphological responses of Leymus chinensis to saline-alkali stress. Grassl. Sci. 2015, 61, 217–226. [Google Scholar] [CrossRef]
- Wang, L.; Fang, C.; Wang, K. Physiological responses of Leymus chinensis to long-term salt, alkali and mixed salt-alkali stresses. J. Plant Nutr. 2015, 38, 526–540. [Google Scholar] [CrossRef]
- Kristanti, R.A.; Hadibarata, T.; Al Farraj, D.A.; Elshikh, S.M.; Alkufeidy, M.R. Biodegradation mechanism of phenanthrene by halophilic Hortaea sp. B15. Water Air Soil Poll. 2018, 229, 1–8. [Google Scholar] [CrossRef]
- Fritts, R.; McCully, A.; McKinley, J. Extracellular metabolism sets the table for microbial cross-feeding. Microbiol. Mol. Biol. Rev. 2021, 85, e00135-20. [Google Scholar] [CrossRef]
- Sunny-Roberts, E.O.; Knorr, D. Evaluation of the response of Lactobacillus rhamnosus VTT E-97800 to sucrose-induced osmotic stress. Food Microbiol. 2008, 25, 183–189. [Google Scholar] [CrossRef]
- Songserm, P.; Srimongkol, P.; Thitiprasert, S. Differential gene expression analysis of Aspergillus terreus revels metabolic response and transcription suppression under dissolved oxygen and pH stress. J. Evol. Biochem. Physiol. 2020, 56, 577–586. [Google Scholar] [CrossRef]
- Orruño, M.; Kaberdin, V.; Arana, I. Survival strategies of Escherichia coli and Vibrio spp.: Contribution of the viable but nonculturable phenotype to their stress-resistance and persistence in adverse environments. World J. Microbiol. Biotechnol. 2017, 33, 45. [Google Scholar] [CrossRef]
- Hicks, D.B.; Liu, J.; Fujisawa, M.; Krulwich, T.A. F1F0-ATP synthases of alkaliphilic bacteria: Lessons from their adaptations. Biochim. Biophys. Acta 2010, 1797, 1362–1377. [Google Scholar] [CrossRef] [PubMed]
- Chew, J.; Zilm, P.S.; Fuss, J.M.; Gully, N.J. A proteomic investigation of Fusobacterium nucleatum alkaline-induced biofilms. BMC Microbiol. 2012, 12, 189. [Google Scholar] [CrossRef]
- Keshri, J.; Mishra, A.; Jha, B. Microbial population index and community structure in saline-alkaline soil using gene targeted metrologics. Microbiol. Res. 2013, 168, 165–173. [Google Scholar] [CrossRef] [PubMed]
- Rangeshwaran, R.; Ashwitha, K.; Sivakumar, G.; Jalali, S.K. Analysis of proteins expressed by an abiotic stress tolerant Pseudomonas putida (NBAII-RPF9) isolate under saline and high temperature conditions. Curr. Microbiol. 2013, 67, 659–667. [Google Scholar] [CrossRef]
- Eguchi, K.; Nakayama, K. Prolonged hypoxia decreases nuclear pyrovate dehydrogenase complex and regulates the gene expression. BBRC 2019, 520, 128–135. [Google Scholar]
- Kikuchi, D.; Minamishima, Y.; Nakayama, K. Prolyl hydroxylase PHD3 interacts with pyruvate dehydrogenase (PDH–E1 β and regulates the cellular PDH activity. BBRC 2014, 451, 288–294. [Google Scholar] [PubMed]
- Kim, J.; Tchernyshyov, I.; Semenza, G.; Dang, C. Hif-1-mediated expression of pyruvate dehydrogenase kinase: A metabolic switch required for cellular adaptation to hypoxia. Cell Metab. 2006, 3, 177–185. [Google Scholar] [CrossRef] [PubMed]
- UrbańSka, K.; Orzechowski, A. Unappreciated role of LDHA and LDHB to control apoptosis and autophagy in tumor cells. Int. J. Mol. Sci. 2019, 20, 2085. [Google Scholar] [CrossRef]
- Pandey, S.; Phulara, S.C.; Jha, A.; Chauhan, P.S.; Gupta, P.; Shukla, V. 3-Methyl-3–buten-1-ol (isoprenol) confers longevity and stress tolerance in Caenorhabditis elegans. Int. J. Food Sci. Nutr. 2019, 70, 595–602. [Google Scholar] [CrossRef]
- Sanadhya, P.; Bucki, P.; Liarzi, O.; Ezra, D.; Gamliel, A.; Braun Miyara, S. Caenorhabditis elegans susceptibility to Daldinia cf. concentrica bioactive volatiles is coupled with expression activation of the stress-response transcription factor daf-16, a part of distinct nematicidal action. PLoS ONE 2018, 13, e0196870. [Google Scholar] [CrossRef]
- Reyna-Beltran, E.; Iranzo, M.; Calderon-Gonzalez, K.G.; Mondragon-Flores, R.; Labra-Barrios, M.L.; Mormeneo, S.; Luna-Arias, J.P. The Candida albicans ENO1 gene encodes a transglutaminase involved in growth, cell division, morphogenesis, and osmotic protection. J. Biol. Chem. 2018, 293, 4304–4323. [Google Scholar] [CrossRef] [PubMed]
- Reading, D.S.; Hallberg, R.L.; Myers, A.M. Characterization of the yeast HSP60 gene coding for a mitochondrial assembly factor. Nature 1989, 337, 655–659. [Google Scholar] [CrossRef]
- Yoneda, Y.; Kawada, K.; Kuramoto, N. Selective Upregulation by Theanine of Slc38a1 Expression in Neural Stem Cell for Brain Wellness. Molecules 2020, 25, 347. [Google Scholar] [CrossRef]
- Hori, K.; Yamamoto, Y.; Tokita, K.; Saito, F.; Kurotsu, T.; Kanda, M.; Okamura, K.; Furuyama, J.; Saito, Y. The nucleotide sequence for a proline-activating domain of gramicidin S synthetase 2 gene from Bacillus brevis. J. Biochem. 1991, 110, 111–119. [Google Scholar] [CrossRef] [PubMed]
- Tran, T.T.; Bott, N.J.; Gelderen, R.V.; Nguyen, N.T.; Chu, P.M.T.; Chu, H.H. Comparative genome analysis of two heterotrophic nitrifying Pseudomonas putida strains isolated from freshwater shrimp ponds in Soc trang province. Fermenation 2022, 8, 336. [Google Scholar] [CrossRef]
- Verslues, P.; Sharma, S. Proline metabolism and its implications for plant environment interaction. ASPB 2010, 8, e0140. [Google Scholar] [CrossRef]
- Rodríguez-Herrero, V.; Payá, G.; Bautista, V.; Vegara, A.; Cortés-Molina, M.; Camacho, M.; Esclapez, J.; Bonete, M. Essentiality of the glnA gene in Haloferax mediterranei: Gene conversion and transcriptional analysis. Extremes 2020, 24, 433–446. [Google Scholar] [CrossRef]
- Abiala, M.A.; Sahoo, L. Bacillus aryabhattai enhanced proline content, stabilized membrane and improved growth of cowpea under NaCl-induced salinity stress. J. Appl. Microbiol. 2022, 133, 1520–1533. [Google Scholar] [CrossRef] [PubMed]
- Nasrallah, G.; Allan, D. The Legionella pneumophila Chaperonin—An Unusual Multifunctional Protein in Unusual Locations. Front. Microbiol. 2011, 2, 122. [Google Scholar]
- Fishbein, S.; Tomasi, F.; Wolf, I.; Dulberger, C.; Wang, A.; Keshishian, H.; Wallace, L.; Carr, S.; Ioerger, T.; Rego, E. The considered translation factor LepA is required for optimal synthesis of a porin family in Mycobacterium smegmatis. J. Bacteriol. 2021, 203, e00604-20. [Google Scholar] [CrossRef]
- Sharma, A.; Boise, L.H.; Shanmugam, M. Cancer metabolism and the evasion of apoptotic cell death. Cancers 2019, 11, 1144. [Google Scholar] [CrossRef] [PubMed]
- Ma, B.; Marcotte, D.; Paramasivam, M.; Michelsen, K.; Wang, T.; Bertolotti-Ciarlet, A.; Jones, J.H.; Moree, B.; Butko, M.; Salafsky, J. ATP competitive mlkl binders have no functional impact on necroptosis. PLoS ONE 2016, 11, 0165983. [Google Scholar] [CrossRef]
- Karl, F. Glutamine signalling in bacteria. Front. Biosci. 2007, 12, 358–370. [Google Scholar]
- Vandenabeele, P.; Galluzzi, L.; Vanden Berghe, T.; Kroemer, G. Molecular mechanisms of necroptosis: An ordered cellular explosion. Nat. Rev. Mol. Cell. Biol. 2010, 11, 700–714. [Google Scholar] [CrossRef]
- Matsuno, K.; Sonenshein, L. Role of SpoVG in asymmetric septation in Bacillus subtilis. J. Bacteriol. 1999, 181, 3392–3401. [Google Scholar] [CrossRef]
- O, Y.K.; Freese, E.B.; Freese, E. abnormal septation and inhibition of dispersion by accumulation of L-α-Glycophosphate in Bacillus subtilis mutants. J. Bacteriol. 1973, 113, 1034–1045. [Google Scholar]
- Huang, Q.; Zhang, Z.; Liu, Q.; Liu, F.; Liu, Y.; Zhang, J.; Wang, G. SpoVG is an important regulator of sporulation and effects biofilm formation by regulating Spo0A transcription in Bacillus cereus 0–9. BMC Microbiol. 2021, 21, 172. [Google Scholar] [CrossRef] [PubMed]
- Alcorlo, M.; Straume, D.; Lutkenhaus, J.; Håvarstein, L.S.; Hermoso, J.A. Structural characterization of the essential cell division protein FtsE and its interaction with FtsX in Streptococcus pneumoniae. Mbio 2020, 11, e01488-20. [Google Scholar] [CrossRef] [PubMed]
- Wang, G.; Zhang, L.; Zhang, S.; Li, B.; Li, J.; Wang, X.; Zhang, J.; Guan, C.; Ji, J. The combined use of a plant growth promoting Bacillus sp. strain and GABA promotes the growth of rice under salt stress by regulating antioxidant enzyme system, enhancing photosynthesis and improving soil enzyme activities. Microbiol. Res. 2023, 266, 127225. [Google Scholar] [CrossRef] [PubMed]
- Dhakal, R.; Bajpai, V.K.; Baek, K.H. Production of gaba (γ-aminobutyric acid) by microorganisms: A review. Braz. J. Microbiol. 2012, 43, 1230–1241. [Google Scholar] [CrossRef] [PubMed]
- Bao, H.; Chen, X.; Lv, S.; Jiang, P.; Feng, J.; Fan, P.; Nie, L.; Li, Y. Virus-induced gene silencing reveals control of reactive oxygen species accumulation and salt tolerance in tomato by γ-Aminobutyric acid metabolic pathway. Plant Cell. Environ. 2015, 38, 600–613. [Google Scholar] [CrossRef] [PubMed]
- Watteau, F.; Villemin, G. Soil microstructures examined through transmission electron microscopy reveal soil-microorganism interactions. Front. Environ. Sci. 2018, 6, 106. [Google Scholar] [CrossRef]
- Hart, E.; Creevey, C.; Hitch, T.; Kingston-Smith, A. Meta proteomics of rumen microbiota indicators niche competitiveness and functional governance in a limited number of metallic pathways between anaerobic bacteria. Sci. Rep. 2018, 8, 10504. [Google Scholar] [CrossRef] [PubMed]
- Khalid, R.; Zhang, X.X.; Hayat, R.; Ahmed, M. Molecular Characteristics of Rhizobia Isolated from Arachis hypogaea Grown under stress environment. Sustainability 2020, 12, 6259. [Google Scholar] [CrossRef]
- Roesgaard, M.; Lundsgaard, J.; Newcombe, E.; Jacobsen, N.; Pesce, F.; Tranchant, E.; Lindemose, S.; Prestel, A.; Hartmann-Petersen, R.; Lindorff-Larsen, K.; et al. Deciding the alphabet of order Glu and ASP act differently on local but not global properties. Biomolecules 2022, 12, 1426. [Google Scholar] [CrossRef]
- Yu, L.; Fink, B.; Som, R. Metabolic clearance of oxaloacetate and mitochondrial complex II resuscitation: Diversion control in skeletal muscle and brown adipose tissue. Biochim. Biophys. Acta Bioenerg. 2023, 1864, 148930. [Google Scholar] [CrossRef]
- Adácsi, C.; Kovács, S.; Pócsi, I.; Pusztahelyi, T. Elimination of deoxynivalenol, aflatoxin B1, and zearalenone by Gram positive microbes (Firmicutes). Toxins 2022, 14, 591. [Google Scholar] [CrossRef]
- Liu, Z.; Huang, M.; Li, A.; Hu, Y. Flocculation and antimicrobial properties of a cationalized starch. Water Res. 2017, 119, 57–66. [Google Scholar] [CrossRef] [PubMed]
- Tinajero-Trejo, M.; Carnell, O.; Kabli, A.; Pasquina-Lemonche, L.; Lafage, L.; Han, A.; Hobbs, J.; Foster, S. The Staphylococcus aureus cell division protein, DivlC, interacts with the cell wall and controls its biosynthesis. Commun. Biol. 2022, 5, 1228. [Google Scholar] [CrossRef] [PubMed]
- Molina-Santiago, C.; Udaondo, Z.; Ramos, J.L. Draft whole-genome sequence of the antibiotic-producing soil isolate Pseudomonas sp. strain 250J. Environ. Microbiol. Rep. 2015, 7, 288–292. [Google Scholar] [CrossRef]
- Alvarez, F.; Simonetti, E.; Draghi, W.O.; Vinacour, M.; Palumbo, M.C.; Do Porto, D.F.; Montecchia, M.S.; Roberts, I.N.; Ruiz, J.A. Genome mining of Burkholderia ambifaria strain T16, a rhizobacterium able to produce antimicrobial compounds and degrade the mycotoxin fusaric acid. World J. Microb. Biot. 2022, 38, 114. [Google Scholar] [CrossRef] [PubMed]
- Suman, B.; Triveni, S.; Latha, P.C.; Srilatha, M.; Rani, C.D. Salinity tolerant phosphorous solubilising bacteria from saline soils of telangana. J. Pharmacogn. Phytochem. 2018, 7, 175–182. [Google Scholar]
- Ahmad, A.; Zafar, U.; Khan, A.; Haq, T.; Mujahid, T.; Wali, M. Effectiveness of compost inoculated with phosphate solubilizing bacteria. J. Appl. Microbiol. 2022, 133, 1115–1129. [Google Scholar] [CrossRef]
- Alburquerque, J.A.; Fuente, C.; Campoy, M.; Carrasco, L.; Nájerab, I.; Baixaulib, C.; Caravaca, F.; Roldán, A.; Cegarra, J.; Bernal, M.P. Agricultural use of digestate for horticultural crop production and improvement of soil properties. Eur. J. Agron. 2012, 43, 119–128. [Google Scholar] [CrossRef]
- Ghosh, S.; Chowdhury, R.; Bhattacharya, P. Sustainability of cereal straws for the fermentative production of second generation biofuels: A review of the efficiency and economics of biochemical pretreatment processes. Appl. Energy 2017, 198, 284–298. [Google Scholar] [CrossRef]


















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Characteristics | JL-5-7 (%) | JL-5-9 (%) |
|---|---|---|
| Total raw read | 30,741,042 | 29,035,034 |
| Genome mapped read (%) | 28,333,104 (92.2) | 26,955,132 (92.8) |
| Unmapped read (%) | 2,407,938 (7.8) | 2,079,902 (7.2) |
| Uniquely mapped read (%) | 26,264,786 (85.4) | 25,249,848 (87.0) |
| Coding sequence (CDS) mapped reads (%) | 25,504,272 (88.2) | 24,246,954 (88.2) |
| Gene ID [log2fc(JL-5-9/JL-5-7)] | Gene: Description | KEGG Pathway: Description | Pathway Component |
|---|---|---|---|
| FORC47_RS19095 [2.724 ↑] | glnA: type I glutamate-ammonia ligase | K01915: glutamine synthetase | map00630; map00220; map00910; map02020; map00250; map04727 (GS); map04724; map04217 (GluL, Hsp90 and RIPK3) |
| FORC47_RS26675 [2.343 ↑] | ftsX: permease-like cell division protein FtsX | K09811: cell division transport system permease protein | map02010 |
| FORC47_RS20430 [1.987 ↑] | FORC47_RS20430: protein-glutamine γ-glutamyltransferase | K00686: protein-glutamine gamma-glutamyltransferase | – |
| FORC47_RS01880 [1.551 ↑] | gatA: Asp-tRNA(Asn)/Glu-tRNA(Gln) amidotransferase subunit GatA | K02433: aspartyl-tRNA(Asn)/glutamyl-tRNA(Gln) amidotransferase subunit A | map00970 |
| FORC47_RS01875 [1.389 ↑] | gatC: Asp-tRNA(Asn)/Glu-tRNA(Gln) amidotransferase subunit GatC | K02435: aspartyl-tRNA(Asn)/glutamyl-tRNA(Gln) amidotransferase subunit C | map00970 |
| FORC47_RS01885 [1.054 ↑] | gatB: Asp-tRNA(Asn)/Glu-tRNA(Gln) amidotransferase subunit GatB | K02434: aspartyl-tRNA(Asn)/glutamyl-tRNA(Gln) amidotransferase subunit B | map00970 |
| FORC47_RS04480 [−1.028 ↓] | FORC47_RS04480: type 1 glutamine amidotransferase | – | – |
| FORC47_RS01550 [−1.150 ↓] | groL: chaperonin GroEL | K04077: chaperonin GroEL | map03018; map05152; map04212; map05134 (LepA, SdhA and HTPB); map04940 |
| FORC47_RS26415 [−1.648 ↓] | eno: phosphopyruvate hydratase | K01689: enolase | map00680; map00010; map03018; map04066 (ENO1, GAPDH, PFKL, PGK1, LDHA and HIF-1) |
| FORC47_RS03370 [−4.241 ↓] | glnH: glutamine ABC transporter substrate-binding protein GlnH | K10039: aspartate/glutamate/glutamine transport system substrate-binding protein | map02010 |
| Treatment | Aboveground Portion (cm) | Underground Portion (cm) | Plant at Tillering Stage (%) |
|---|---|---|---|
| Control | 5.3 ± 0.7c | 3.5 ± 0.5c | 22 ± 6c |
| JL-5 | 5.4 ± 0.4c | 6.1 ± 0.8b | 78 ± 4a |
| XW-4 | 5.6 ± 0.6b | 4.1 ± 0.4c | 51 ± 5b |
| JL-5 and XW-4 | 7.4 ± 0.6a | 6.6 ± 1.0a | 57 ± 3b |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, Y.; Wang, Y.; Zhang, Q.; Fan, H.; Wang, X.; Wang, J.; Zhou, Y.; Chen, Z.; Sun, F.; Cui, X. Saline-Alkali Soil Property Improved by the Synergistic Effects of Priestia aryabhattai JL-5, Staphylococcus pseudoxylosus XW-4, Leymus chinensis and Soil Microbiota. Int. J. Mol. Sci. 2023, 24, 7737. https://doi.org/10.3390/ijms24097737
Wang Y, Wang Y, Zhang Q, Fan H, Wang X, Wang J, Zhou Y, Chen Z, Sun F, Cui X. Saline-Alkali Soil Property Improved by the Synergistic Effects of Priestia aryabhattai JL-5, Staphylococcus pseudoxylosus XW-4, Leymus chinensis and Soil Microbiota. International Journal of Molecular Sciences. 2023; 24(9):7737. https://doi.org/10.3390/ijms24097737
Chicago/Turabian StyleWang, Yujue, Yan Wang, Qian Zhang, Hangzhe Fan, Xinyu Wang, Jianan Wang, Ying Zhou, Zhanyu Chen, Fengjie Sun, and Xiyan Cui. 2023. "Saline-Alkali Soil Property Improved by the Synergistic Effects of Priestia aryabhattai JL-5, Staphylococcus pseudoxylosus XW-4, Leymus chinensis and Soil Microbiota" International Journal of Molecular Sciences 24, no. 9: 7737. https://doi.org/10.3390/ijms24097737