
Genome-Wide Identification and Evolution Analysis of the CYP76 Subfamily in Rice (Oryza sativa)

 , ,
, ,
Abstract
:1. Introduction
2. Results
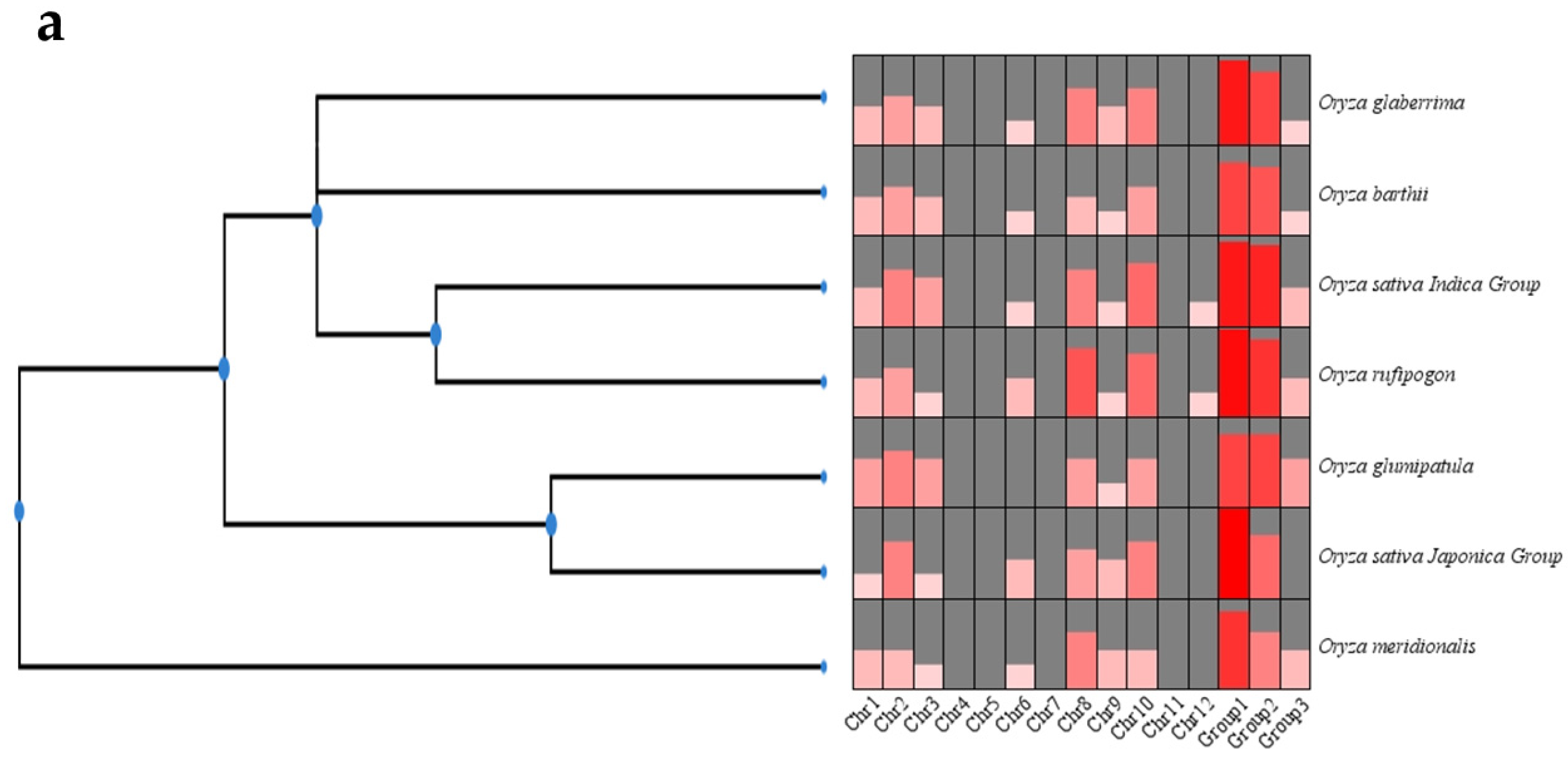
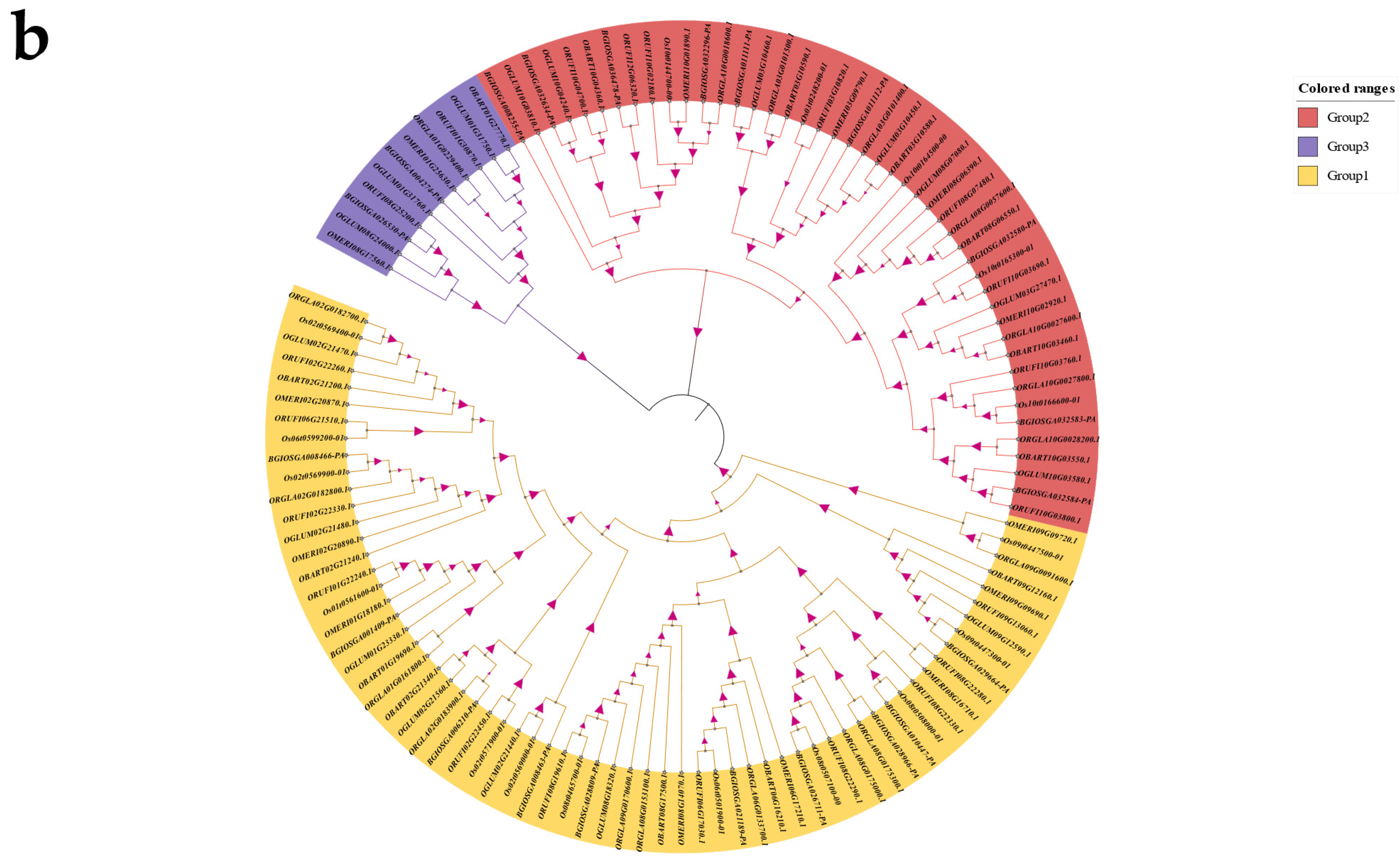
2.1. Identification of CYP76 Genes in Oryza Genus
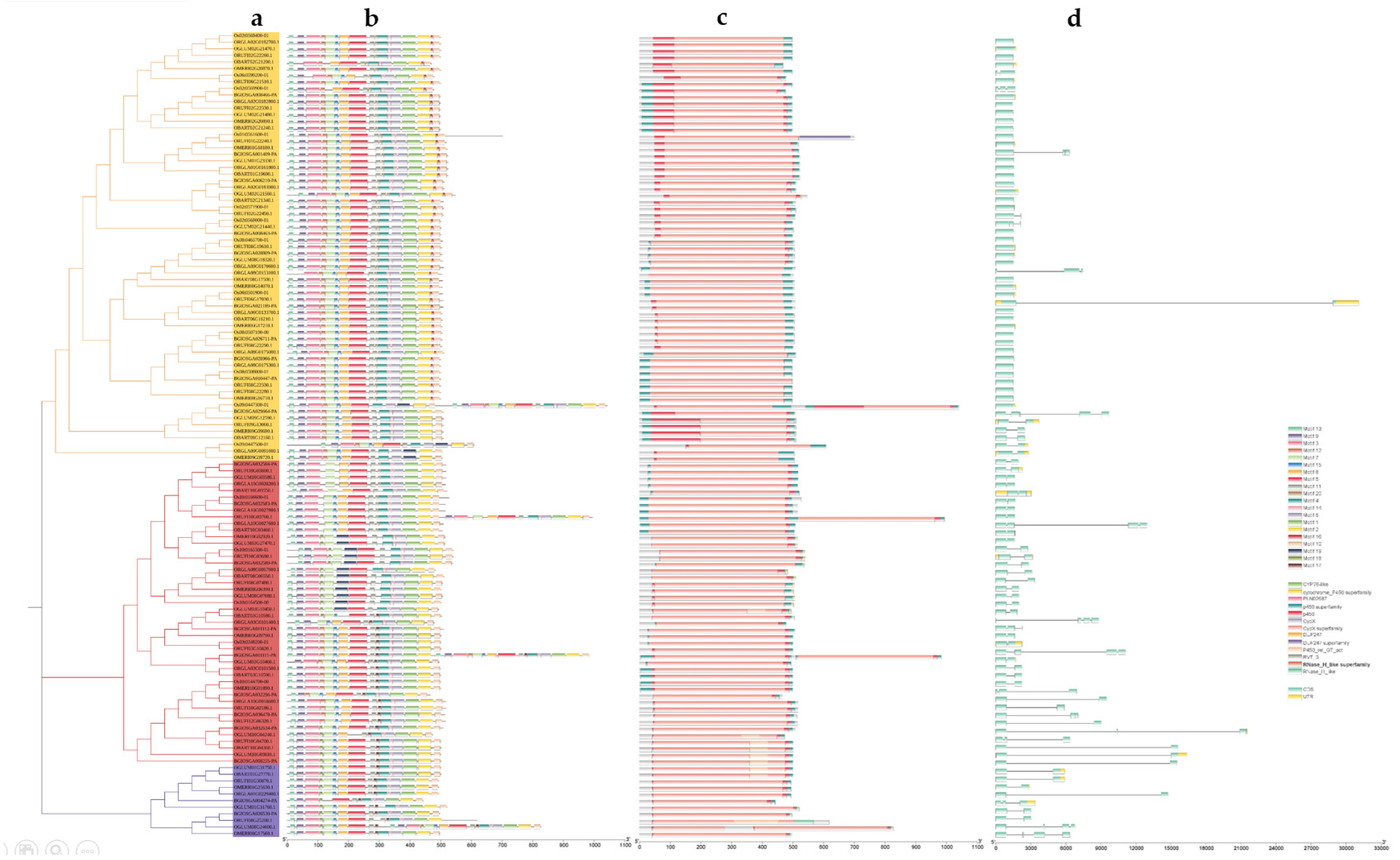
2.2. Gene Structure, Conserved Motif, and Domain Analysis
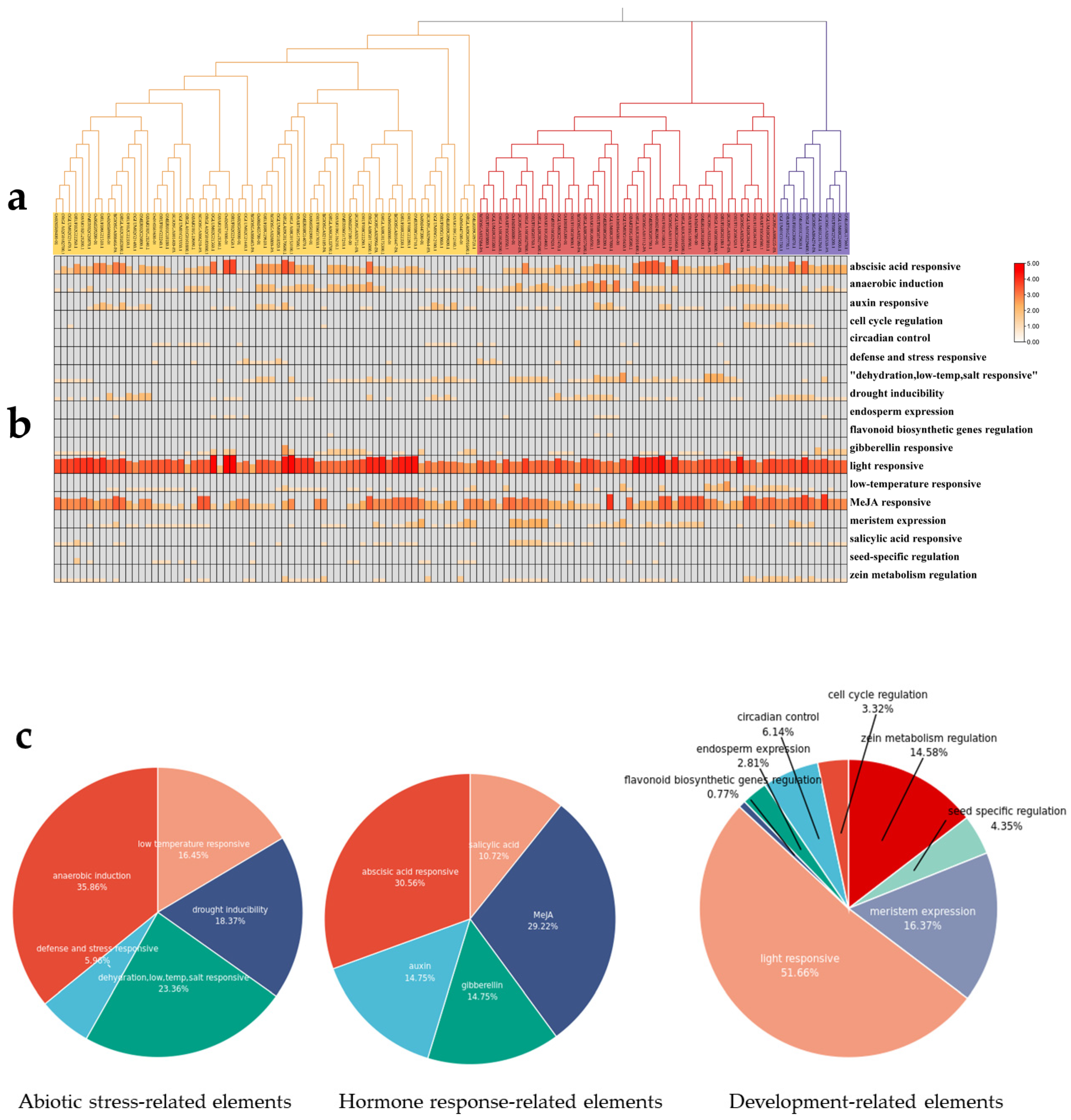
2.3. Cis-Elements Analysis of CYP76s
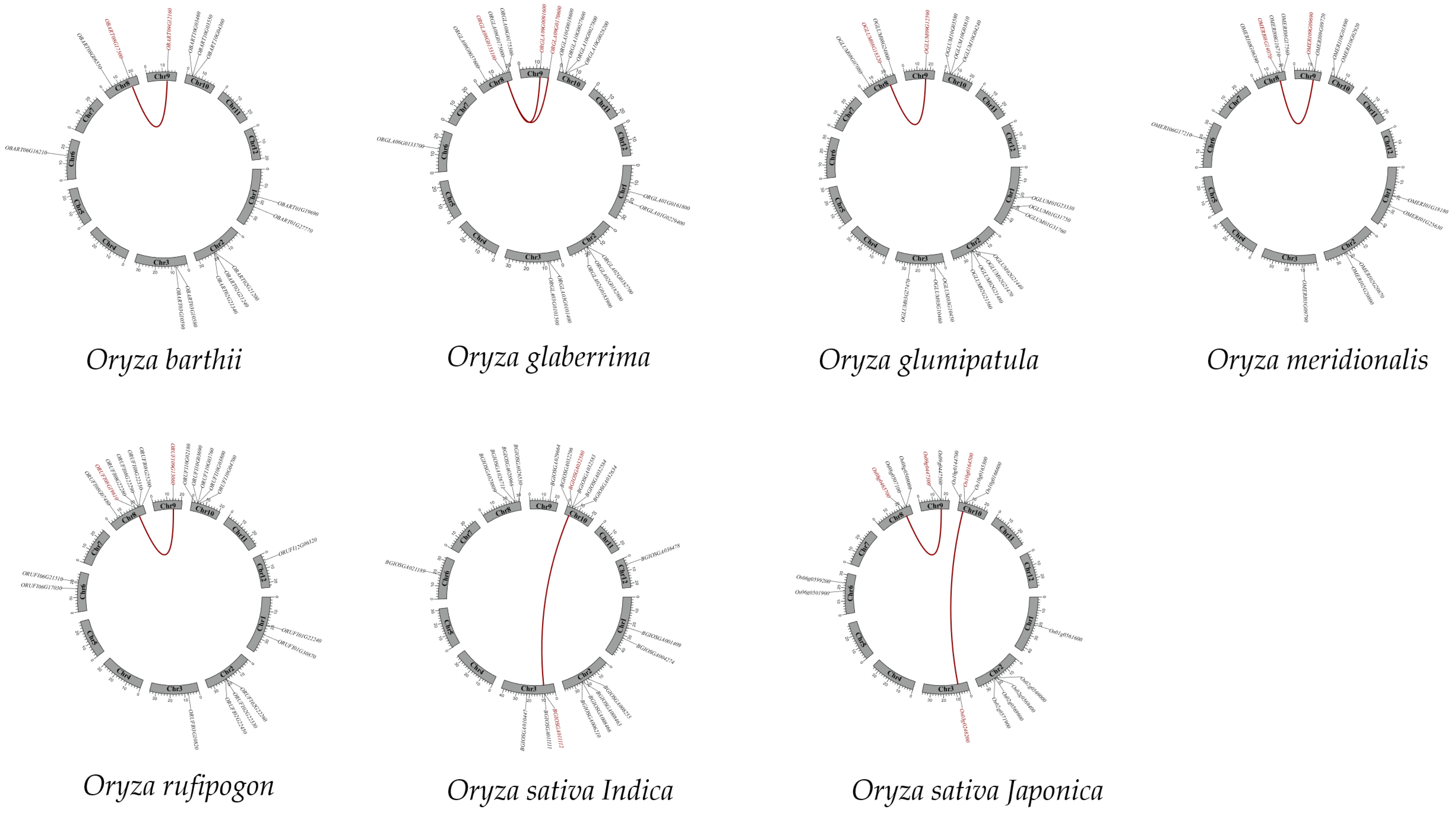
2.4. Collinearity and Selective Forces Analysis of CYP76s
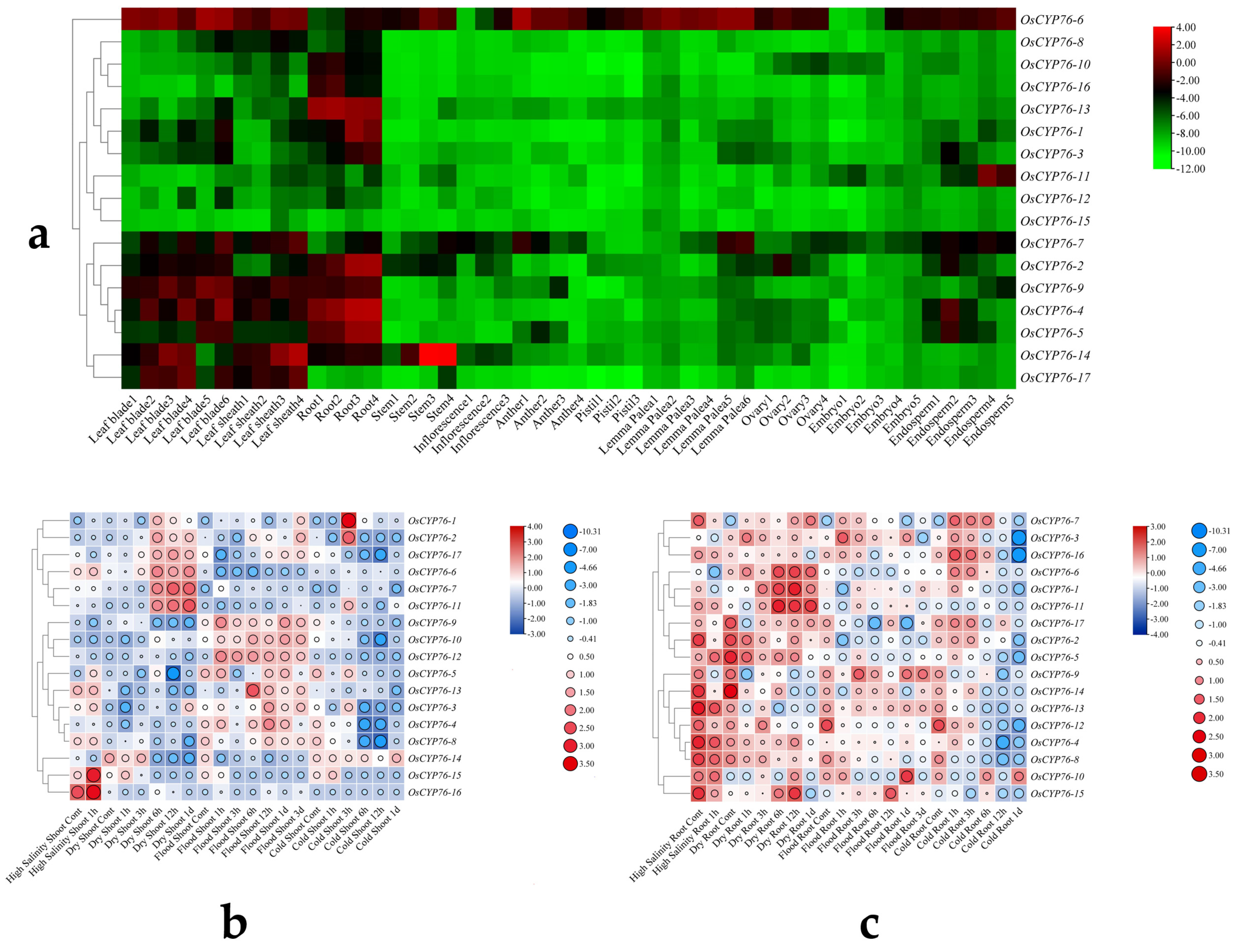
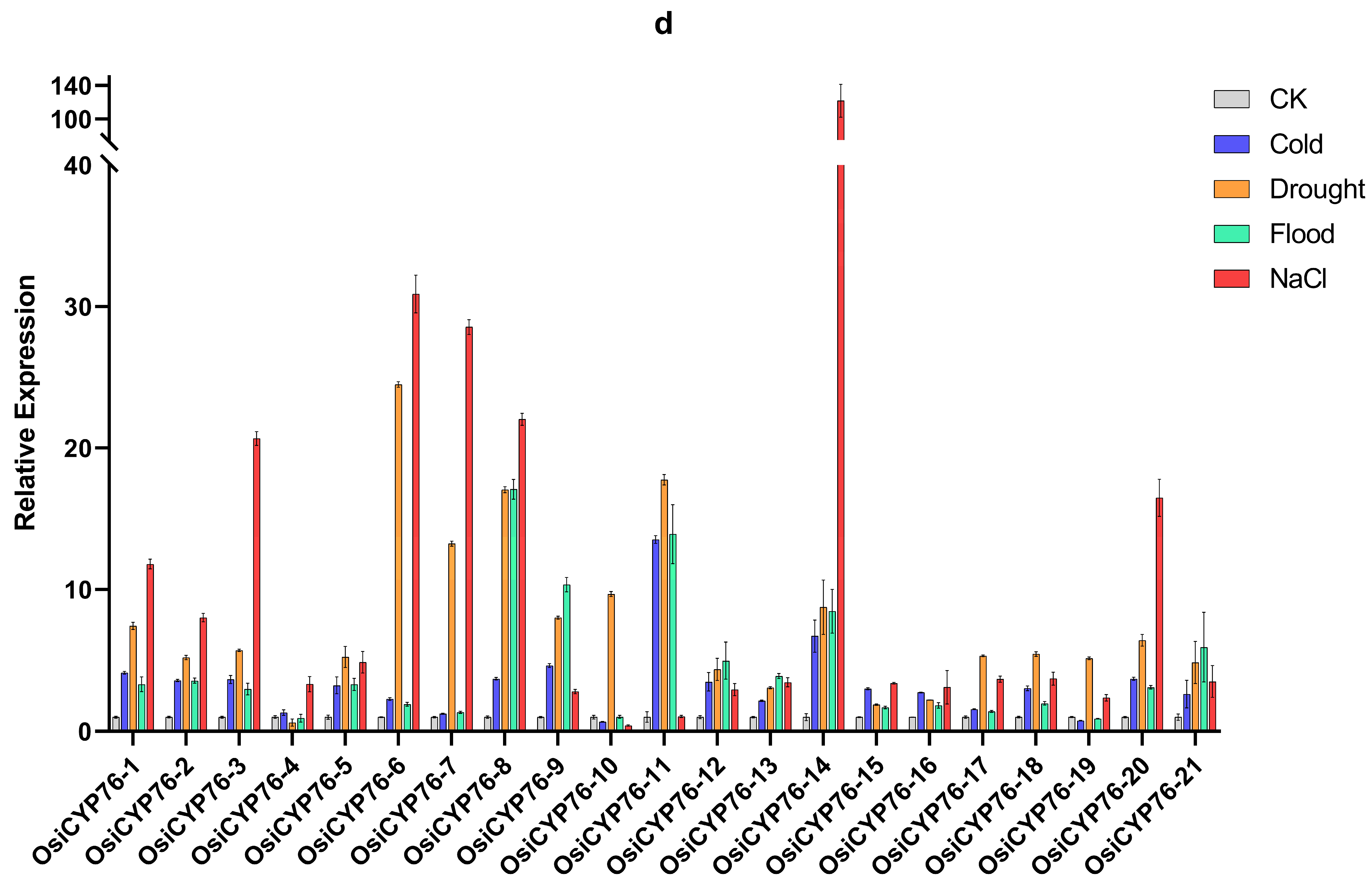
2.5. Expression Pattern Analysis of OsCYP76 Genes and RNA-seq Analysis under Abiotic Stresses
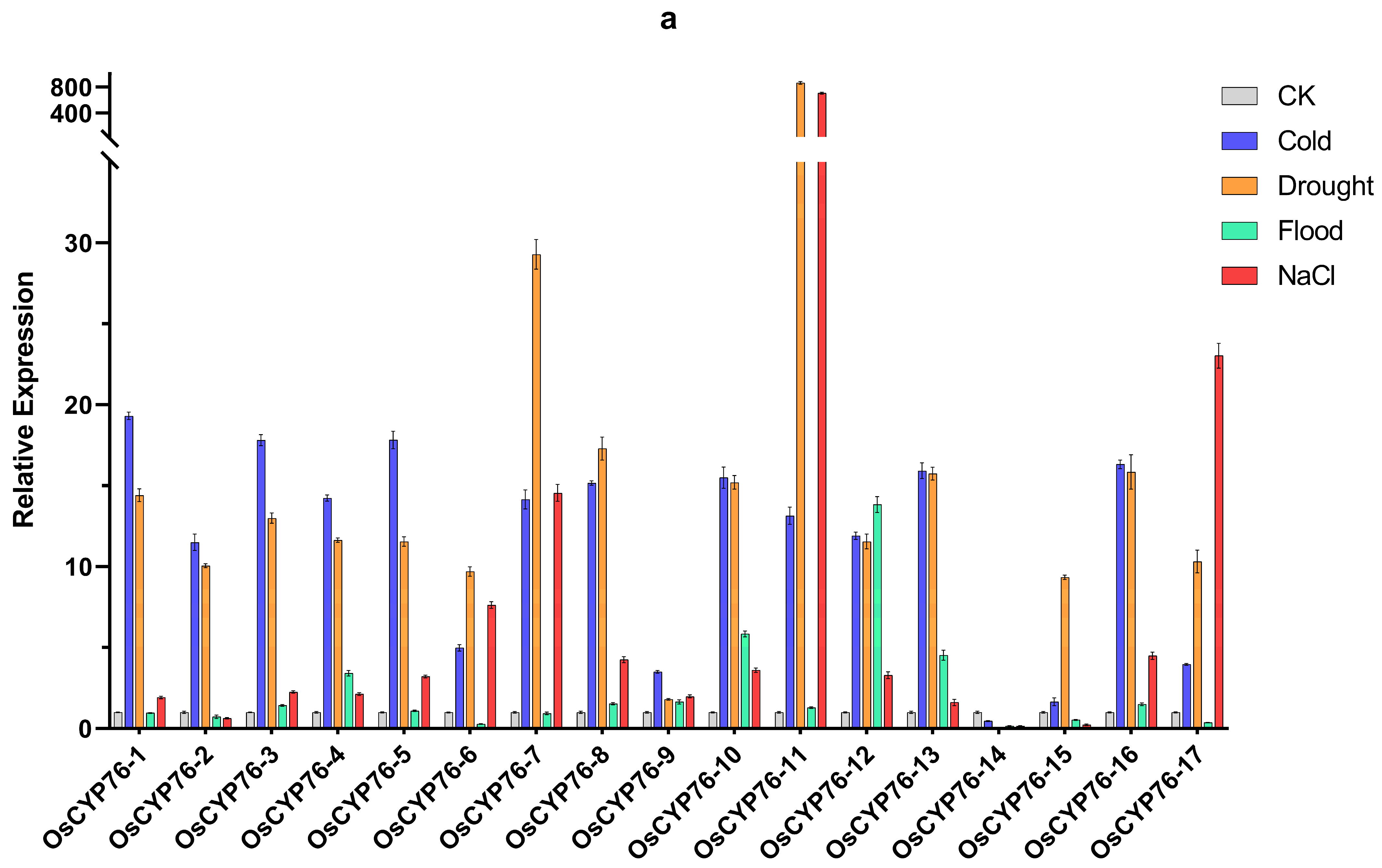
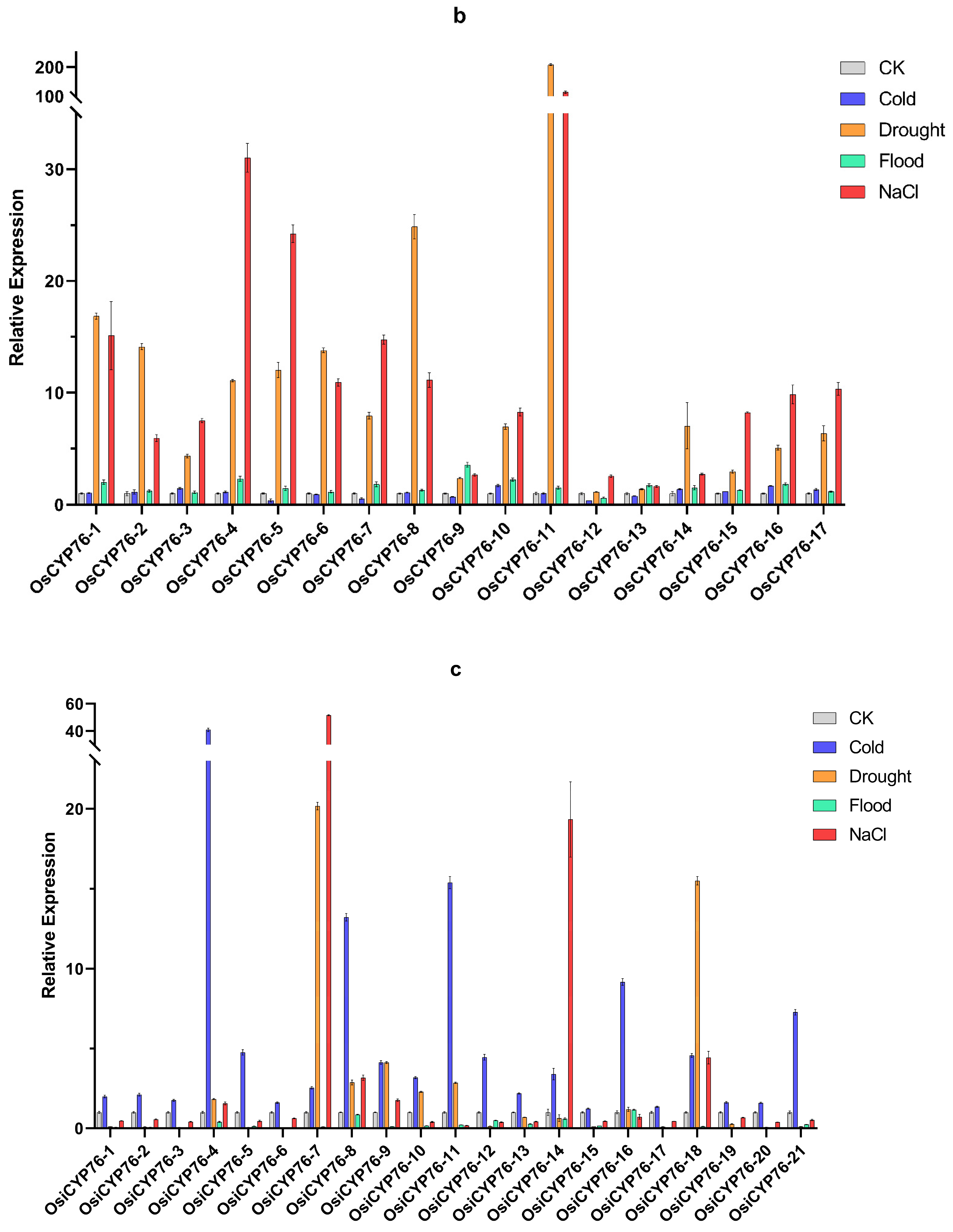
2.6. CYP76 in O. sativa, Japonica and O. sativa, Indica Were Shown Different Responses to Abiotic Stresses through qRT-PCR Analysis
3. Discussion
4. Materials and Methods
4.1. Plant Material, Growth, and Abiotic Stress Conditions
4.2. Identification and Phylogenetic Analysis of CYP76 Genes in Rice
4.3. Sequence and Protein Properties of CYP76 Genes in Rice
4.4. Chromosomal Localization, Orthogroup Identification, and Collinearity Analysis
4.5. Expression Analysis of OsCYP76 Genes
4.6. RNA Extraction and qPCR Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Bathe, U.; Tissier, A. Cytochrome P450 enzymes: A driving force of plant diterpene diversity. Phytochemistry 2019, 161, 149–162. [Google Scholar] [CrossRef] [PubMed]
- Bernhardt, R. Cytochromes P450 as versatile biocatalysts. J. Biotechnol. 2006, 124, 128–145. [Google Scholar] [CrossRef] [PubMed]
- Nelson, D.; Werck-Reichhart, D. A P450-centric view of plant evolution. Plant J. 2011, 66, 194–211. [Google Scholar] [CrossRef] [PubMed]
- Capdevila, J.H.; Falck, J.R.; Estabrook, R.W. Cytochrome-P450 and the Arachidonate Cascade. Faseb J. 1992, 6, 731–736. [Google Scholar] [CrossRef]
- Hohn, T.; Corsten, S.; Rieke, S.; Müller, M.; Rothnie, H. Methylation of coding region alone inhibits gene expression in plant protoplasts. Proc. Natl. Acad. Sci. USA 1996, 93, 8334–8339. [Google Scholar] [CrossRef]
- Wrighton, S.A.; Stevens, J.C. The Human Hepatic Cytochromes-P450 Involved in Drug-Metabolism. Crit. Rev. Toxicol. 1992, 22, 1–21. [Google Scholar] [CrossRef]
- León, J.; Yalpani, N.; Raskin, I.; Lawton, M.A. Induction of Benzoic-Acid 2-Hydroxylase in Virus-Inoculated Tobacco. Plant Physiol. 1993, 103, 323–328. [Google Scholar] [CrossRef]
- Pauli, H.H.; Kutchan, T.M. Molecular cloning and functional heterologous expression of two alleles encoding (S)-N-methylcoclaurine 3’-hydroxylase (CYP80B1), a new methyl jasmonate-inducible cytochrome P-450-dependent mono-oxygenase of benzylisoquinoline alkaloid biosynthesis. Plant J. 1998, 13, 793–801. [Google Scholar] [CrossRef]
- Mizutani, M.; Ward, E.; Ohta, D. Cytochrome p450 superfamily in Arabidopsis thaliana: Isolation of cDNAs, differential expression, and RFLP mapping of multiple cytochromes p450. Plant Mol. Biol. 1998, 37, 39–52. [Google Scholar] [CrossRef]
- Collu, G.; Unver, N.; Peltenburg-Looman, A.M.; van der Heijden, R.; Verpoorte, R.; Memelink, J. Geraniol 10-hydroxylase, a cytochrome P450 enzyme involved in terpenoid indole alkaloid biosynthesis. FEBS Lett. 2001, 508, 215–220. [Google Scholar] [CrossRef]
- Höfer, R.; Dong, L.; André, F.; Ginglinger, J.F.; Lugan, R.; Gavira, C.; Werck-Reichhart, D. Geraniol hydroxylase and hydroxygeraniol oxidase activities of the CYP76 family of cytochrome P450 enzymes and potential for engineering the early steps of the (seco)iridoid pathway. Metab Eng. 2013, 20, 221–232. [Google Scholar] [CrossRef] [PubMed]
- Miettinen, K.; Dong, L.; Navrot, N.; Schneider, T.; Burlat, V.; Pollier, J.; Werck-Reichhart, D. The seco-iridoid pathway from Catharanthus roseus. Nat. Commun. 2014, 5, 3606. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Hillwig, M.; Okada, K.; Yamazaki, K.; Wu, Y.; Swaminathan, S.; Yamane, H.; Peters, R.J. Characterization of CYP76M5-8 Indicates Metabolic Plasticity within a Plant Biosynthetic Gene Cluster. J. Biol. Chem. 2012, 287, 6159–6168. [Google Scholar] [CrossRef]
- Rastogi, S.; Satapathy, S.; Shah, S.; Mytrai; Prakash, H. In silico identification of cytochrome P450s involved in Ocimum tenuiflorum subjected to four abiotic stresses. Gene Rep. 2020, 20, 100781. [Google Scholar] [CrossRef]
- Wang, C.; Xue, L.; Cui, Q.; Liu, Q.; Zhang, J.; Rao, G. Genome-wide identification of the cytochrome P450 superfamily in Olea europaea helps elucidate the synthesis pathway of oleuropein to improve the quality of olive oil. Sci. Hortic. 2022, 304, 111291. [Google Scholar] [CrossRef]
- Wang, J.; Liu, Y.; Cai, Y.; Zhang, F.; Xia, G.; Xiang, F. Cloning and Functional Analysis of Geraniol 10-Hydroxylase, a Cytochrome P450 from Swertia mussotii Franch. Biosci. Biotechnol. Biochem. 2010, 74, 1583–1590. [Google Scholar] [CrossRef]
- Godiard, L.; Sauviac, L.; Dalbin, N.; Liaubet, L.; Callard, D.; Czernic, P.; Marco, Y. CYP76C2, an Arabidopsis thaliana cytochrome P450 gene expressed during hypersensitive and developmental cell death. FEBS Lett. 1998, 438, 245–249. [Google Scholar] [CrossRef]
- Zhai, X.Y.; Chen, Z.J.; Liu, J.; Zhang, N.; Yang, H. Expression of CYP76C6 Facilitates Isoproturon Metabolism and Detoxification in Rice. J. Agric. Food Chem. 2022, 70, 4599–4610. [Google Scholar] [CrossRef]
- Swaminathan, S.; Morrone, D.; Wang, Q.; Fulton, D.B.; Peters, R.J. CYP76M7 is an ent-cassadiene C11alpha-hydroxylase defining a second multifunctional diterpenoid biosynthetic gene cluster in rice. Plant Cell 2009, 21, 3315–3325. [Google Scholar] [CrossRef] [PubMed]
- Ralston, L.; Kwon, S.T.; Schoenbeck, M.; Ralston, J.; Schenk, D.J.; Coates, R.M.; Chappell, J. Cloning, heterologous expression, and functional characterization of 5-epi-aristolochene-1,3-dihydroxylase from tobacco (Nicotiana tabacum). Arch. Biochem. Biophys. 2001, 393, 222–235. [Google Scholar] [CrossRef] [PubMed]
- Wang, E.; Wang, R.; DeParasis, J.; Loughrin, J.H.; Gan, S.; Wagner, G.J. Suppression of a P450 hydroxylase gene in plant trichome glands enhances natural-product-based aphid resistance. Nat. Biotechnol. 2001, 19, 371–374. [Google Scholar] [CrossRef] [PubMed]
- Salim, V.; Wiens, B.; Masada-Atsumi, S.; Yu, F.; De Luca, V. 7-Deoxyloganetic acid synthase catalyzes a key 3 step oxidation to form 7-deoxyloganetic acid in Catharanthus roseus iridoid biosynthesis. Phytochemistry 2014, 101, 23–31. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; Wang, Q.; Hillwig, M.; Peters, R.J. Picking sides: Distinct roles for CYP76M6 and CYP76M8 in rice oryzalexin biosynthesis. Biochem. J. 2013, 454, 209–216. [Google Scholar] [CrossRef] [PubMed]
- Höfer, R.; Boachon, B.; Renault, H.; Gavira, C.; Miesch, L.; Iglesias, J.; Werck-Reichhart, D. Dual Function of the Cytochrome P450 CYP76 Family from Arabidopsis thaliana in the Metabolism of Monoterpenols and Phenylurea Herbicides. Plant Physiol. 2014, 166, 1149–1161. [Google Scholar] [CrossRef]
- Song, Y.; Wang, C.; Linderholm, H.W.; Fu, Y.; Cai, W.; Xu, J.; Zhuang, L.; Wu, M.; Shi, Y.; Wang, G.; et al. The negative impact of increasing temperatures on rice yields in southern China. Sci. Total Environ. 2022, 820, 153262. [Google Scholar] [CrossRef]
- Zhang, Z.H.; Gao, S.P.; Chu, C.C. Improvement of nutrient use efficiency in rice: Current toolbox and future perspectives. Theor. Appl. Genet. 2020, 133, 1365–1384. [Google Scholar] [CrossRef]
- Kong, W.; Zhang, Y.; Deng, X.; Li, S.; Zhang, C.; Li, Y. Comparative Genomic and Transcriptomic Analysis Suggests the Evolutionary Dynamic of GH3 Genes in Gramineae Crops. Front. Plant Sci. 2019, 10, 1297. [Google Scholar] [CrossRef]
- Jiang, Y.; Liu, X.; Zhou, M.; Yang, J.; Ke, S.; Li, Y. Genome-Wide Identification of the AGC Protein Kinase Gene Family Related to Photosynthesis in Rice (Oryza sativa). Int. J. Mol. Sci. 2022, 23, 12557. [Google Scholar] [CrossRef]
- Zhang, C.; Nie, X.; Kong, W.; Deng, X.; Sun, T.; Liu, X.; Li, Y. Genome-Wide Identification and Evolution Analysis of the Gibberellin Oxidase Gene Family in Six Gramineae Crops. Genes 2022, 13, 863. [Google Scholar] [CrossRef]
- Winter, D.; Vinegar, B.; Nahal, H.; Ammar, R.; Wilson, G.V.; Provart, N.J. An “Electronic Fluorescent Pictograph” Browser for Exploring and Analyzing Large-Scale Biological Data Sets. PLoS ONE 2007, 2, e718. [Google Scholar] [CrossRef]
- Paysan-Lafosse, T.; Blum, M.; Chuguransky, S.; Grego, T.; Pinto, B.L.; Salazar, G.A.; Bateman, A. InterPro in 2022. Nucleic Acids Res. 2022, 51, D418–D427. [Google Scholar] [CrossRef] [PubMed]
- Potter, S.C.; Luciani, A.; Eddy, S.R.; Park, Y.; López, R.; Finn, R.D. HMMER web server: 2018 update. Nucleic Acids Res. 2018, 46, W200–W204. [Google Scholar] [CrossRef] [PubMed]
- Kong, W.; Sun, T.; Zhang, C.; Qiang, Y.; Li, Y. Micro-Evolution Analysis Reveals Diverged Patterns of Polyol Transporters in Seven Gramineae Crops. Front. Genet. 2020, 11, 565. [Google Scholar] [CrossRef] [PubMed]
- Larkin, M.A.; Blackshields, G.; Brown, N.P.; Chenna, R.; McGettigan, P.A.; McWilliam, H.; Valentin, F.; Wallace, I.M.; Wilm, A.; Lopez, R.; et al. Clustal W and clustal X version 2.0. Bioinformatics 2007, 23, 2947–2948. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across Computing Platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Letunic, I.; Bork, P. Bork, Interactive Tree of Life (iTOL) v5: An online tool for phylogenetic tree display and annotation. Nucleic Acids Res. 2021, 49, W293–W296. [Google Scholar] [CrossRef]
- Chen, C.J.; Chen, H.; Zhang, Y.; Thomas, H.R.; Frank, M.H.; He, Y.H.; Xia, R. TBtools: An Integrative Toolkit Developed for Interactive Analyses of Big Biological Data. Mol. Plant 2020, 13, 1194–1202. [Google Scholar] [CrossRef]
- Bailey, T.L.; Johnson, J.; Grant, C.E.; Noble, W.S. The MEME Suite. Nucleic Acids Res. 2015, 43, W39–W49. [Google Scholar] [CrossRef]
- Chou, K.-C.; Shen, H.-B. Cell-PLoc 2.0: An improved package of web-servers for predicting subcellular localization of proteins in various organisms. Nat. Sci. 2010, 2, 1090–1103. [Google Scholar] [CrossRef]
- Horton, P.; Park, K.-J.; Obayashi, T.; Fujita, N.; Harada, H.; Adams-Collier, C.J.; Nakai, K. WoLF PSORT: Protein localization predictor. Nucleic Acids Res. 2007, 35, W585–W587. [Google Scholar] [CrossRef]
- Lescot, M.; Déhais, P.; Thijs, G.; Marchal, K.; Moreau, Y.; Van de Peer, Y.; Rouzé, P.; Rombauts, S. PlantCARE, a database of plant cis-acting regulatory elements and a portal to tools for in silico analysis of promoter sequences. Nucleic Acids Res. 2002, 30, 325–327. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Tang, H.; DeBarry, J.D.; Tan, X.; Li, J.; Wang, X.; Lee, T.-H.; Jin, H.; Marler, B.; Guo, H.; et al. MCScanX: A toolkit for detection and evolutionary analysis of gene synteny and collinearity. Nucleic Acids Res. 2012, 40, e49. [Google Scholar] [CrossRef]
- Yuan, S.; Xu, B.; Zhang, J.; Xie, Z.; Cheng, Q.; Yang, Z.; Cai, Q.; Huang, B. Comprehensive analysis of CCCH-type zinc finger family genes facilitates functional gene discovery and reflects recent allopolyploidization event in tetraploid switchgrass. BMC Genom. 2015, 16, 129. [Google Scholar] [CrossRef] [PubMed]
- Buchfink, B.; Xie, C.; Huson, D.H. Fast and sensitive protein alignment using DIAMOND. Nat. Methods 2015, 12, 59–60. [Google Scholar] [CrossRef]
- Emms, D.M.; Kelly, S. OrthoFinder: Solving fundamental biases in whole genome comparisons dramatically improves orthogroup inference accuracy. Genome Biol. 2015, 16, 157. [Google Scholar] [CrossRef]
- Kong, W.; Gong, Z.; Zhong, H.; Zhang, Y.; Zhao, G.; Gautam, M.; Deng, X.; Liu, C.; Zhang, C.; Li, Y. Expansion and Evolutionary Patterns of Glycosyltransferase Family 8 in Gramineae Crop Genomes and Their Expression under Salt and Cold Stresses in Oryza sativa ssp. japonica. Biomolecules 2019, 9, 188. [Google Scholar] [CrossRef]
- Zhong, H.; Kong, W.; Gong, Z.; Fang, X.; Deng, X.; Liu, C.; Li, Y. Evolutionary Analyses Reveal Diverged Patterns of SQUAMOSA Promoter Binding Protein-Like (SPL) Gene Family in Oryza Genus. Front. Plant Sci. 2019, 10, 565. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Oryza | Gene Pairs | Ka/Ks | Date (Mya) | Type of Selection | Type of Duplication |
|---|---|---|---|---|---|
| O. sativa, japonica | Os10t0164500-00/Os03t0248200-01 | 0.33 | 70.73 | Purifying | SD/WGD |
| Os08t0465700-01/Os09t0447300-01 | 0.58 | 53.93 | Purifying | SD/WGD | |
| Os02t0569000-01/Os02t0569400-01 | 0.62 | 28.38 | Purifying | Tandem | |
| Os09t0447300-01/Os09t0447500-01 | 0.62 | 50.43 | Purifying | Tandem | |
| O. sativa, indica | BGIOSGA032580-PA/BGIOSGA011112-PA | 0.30 | 88.64 | Purifying | SD/WGD |
| BGIOSGA011112-PA/BGIOSGA011111-PA | 0.49 | 22.41 | Purifying | Tandem | |
| O. rufipogon | ORUFI08G19610.1/ORUFI09G13060.1 | 0.58 | 53.97 | Purifying | SD/WGD |
| ORUFI08G22280.1/ORUFI08G22290.1 | 0.47 | 16.07 | Purifying | Tandem | |
| O. meridionalis | OMERI08G14070.1/OMERI09G09690.1 | 0.62 | 51.28 | Purifying | SD/WGD |
| O. glumaepatula | OGLUM08G18320.1/OGLUM09G12590.1 | 0.60 | 53.01 | Purifying | SD/WGD |
| OGLUM01G31750.1/OGLUM01G31760.1 | 0.22 | 63.73 | Purifying | Tandem | |
| OGLUM02G21470.1/OGLUM02G21480.1 | 0.29 | 11.64 | Purifying | Tandem | |
| O. glaberrima | ORGLA08G0153100.1/ORGLA09G0091600.1 | 0.67 | 49.01 | Purifying | SD/WGD |
| ORGLA08G0153100.1/ORGLA09G0170600.1 | 0.62 | 50.55 | Purifying | SD/WGD | |
| ORGLA02G0182700.1/ORGLA02G0182800.1 | 0.27 | 12.99 | Purifying | Tandem | |
| ORGLA03G0101400.1/ORGLA03G0101500.1 | 0.48 | 23.09 | Purifying | Tandem | |
| O. barthii | OBART08G17500.1/OBART09G12160.1 | 0.58 | 53.54 | Purifying | SD/WGD |
| OBART03G10580.1/OBART03G10590.1 | 0.47 | 23.33 | Purifying | Tandem |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhou, M.; Jiang, Y.; Liu, X.; Kong, W.; Zhang, C.; Yang, J.; Ke, S.; Li, Y. Genome-Wide Identification and Evolution Analysis of the CYP76 Subfamily in Rice (Oryza sativa). Int. J. Mol. Sci. 2023, 24, 8522. https://doi.org/10.3390/ijms24108522
Zhou M, Jiang Y, Liu X, Kong W, Zhang C, Yang J, Ke S, Li Y. Genome-Wide Identification and Evolution Analysis of the CYP76 Subfamily in Rice (Oryza sativa). International Journal of Molecular Sciences. 2023; 24(10):8522. https://doi.org/10.3390/ijms24108522
Chicago/Turabian StyleZhou, Mingao, Yifei Jiang, Xuhui Liu, Weilong Kong, Chenhao Zhang, Jian Yang, Simin Ke, and Yangsheng Li. 2023. "Genome-Wide Identification and Evolution Analysis of the CYP76 Subfamily in Rice (Oryza sativa)" International Journal of Molecular Sciences 24, no. 10: 8522. https://doi.org/10.3390/ijms24108522