
N-Acetylaspartate and Choline Metabolites in Cortical and Subcortical Regions in Clinical High Risk Relative to Healthy Control Subjects: An Exploratory 7T MRSI Study
 , ,
, ,
Abstract
:1. Introduction
2. Results
2.1. Clinical and Demographic Measures
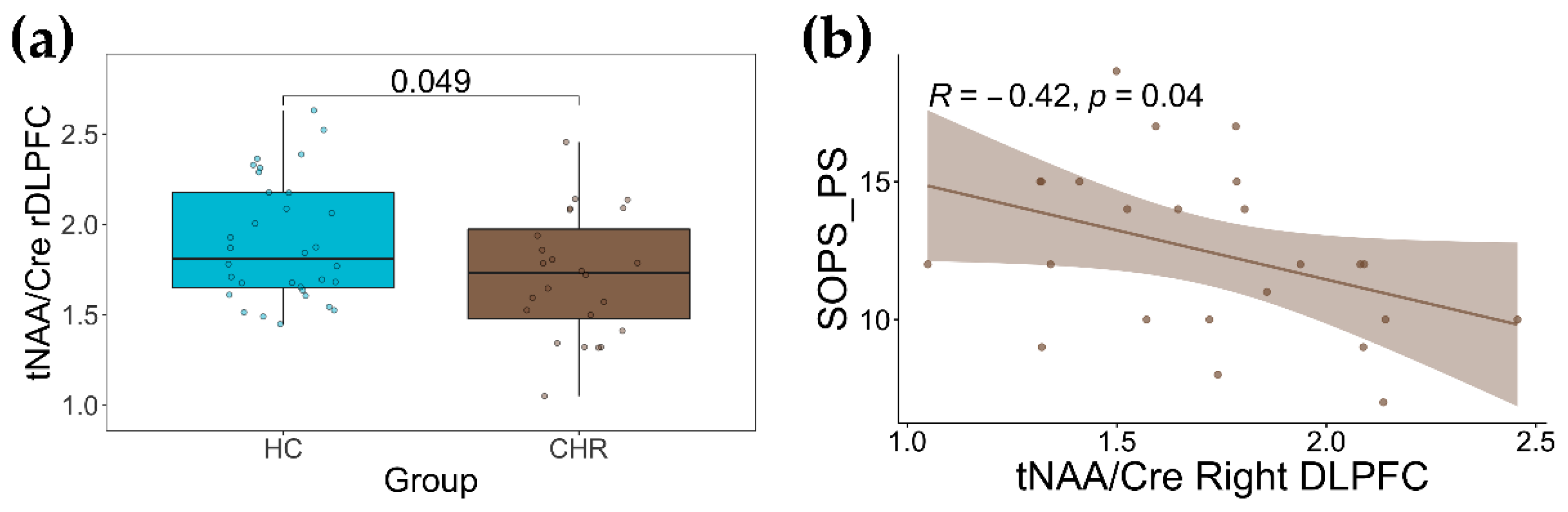
2.2. Group Differences in tNAA/Cre and tCho/Cre
2.3. Association with Clinical Measurements
3. Discussion
4. Materials and Methods
4.1. Participants
4.2. Recruitment, Eligibility Criteria, and Clinical and Cognition Measurements
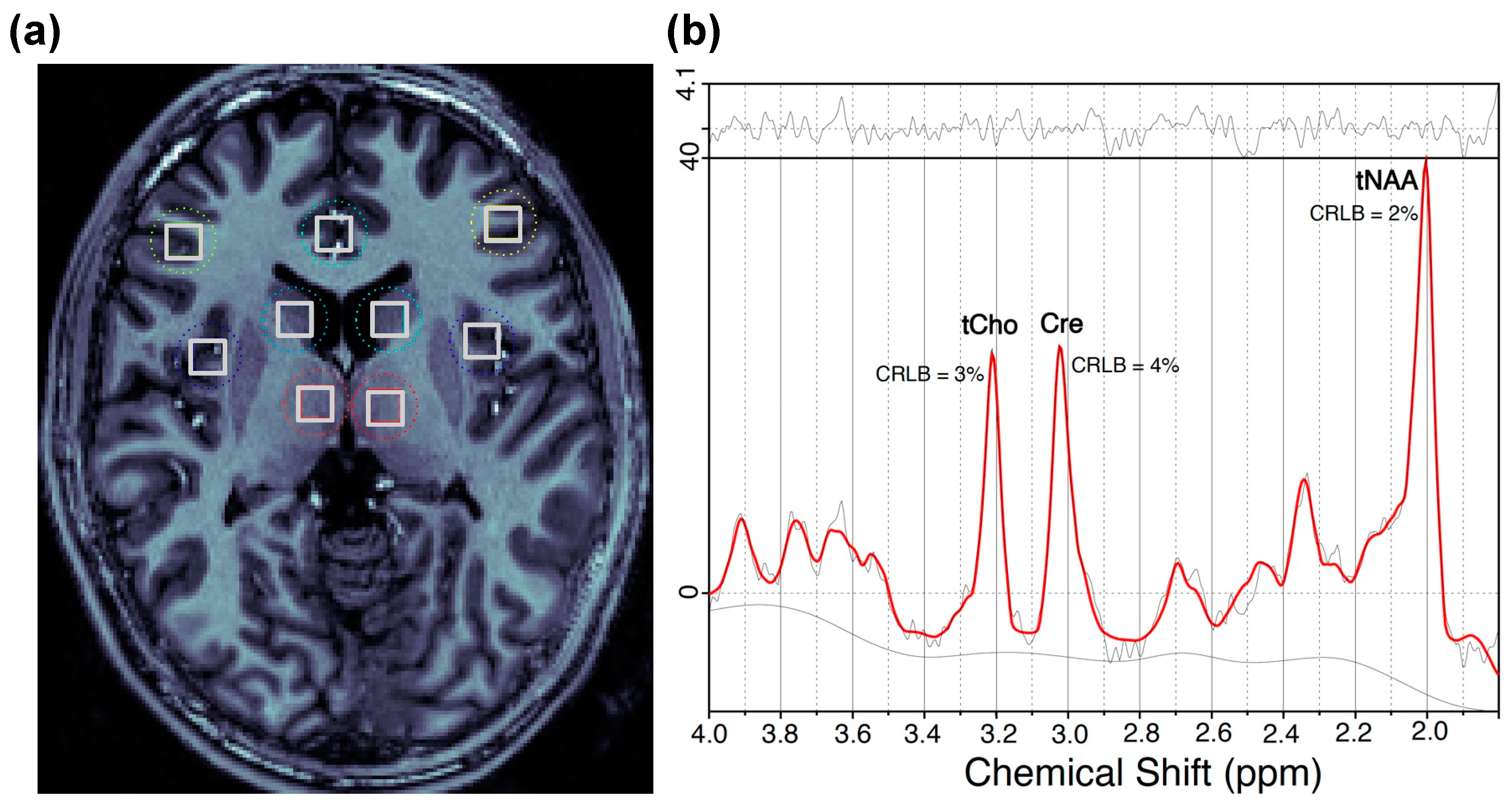
4.3. 1H MRSI Data Acquisition and Processing
4.4. Regions of Interest
4.5. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- GBD 2019 Diseases and Injuries Collaborators. Global burden of 369 diseases and injuries in 204 countries and territories, 1990–2019: A systematic analysis for the Global Burden of Disease Study 2019. Lancet 2020, 396, 1204–1222. [Google Scholar] [CrossRef] [PubMed]
- Lewis-Fernández, R.; Rotheram-Borus, M.J.; Betts, V.T.; Greenman, L.; Essock, S.M.; Escobar, J.I.; Barch, D.; Hogan, M.F.; Arean, P.A.; Druss, B.G.; et al. Rethinking funding priorities in mental health research. Br. J. Psychiatry 2016, 208, 507–509. [Google Scholar] [CrossRef] [PubMed]
- Ferrarelli, F.; Mathalon, D. The prodromal phase: Time to broaden the scope beyond transition to psychosis? Schizophr. Res. 2020, 216, 5. [Google Scholar] [CrossRef] [PubMed]
- Fusar-Poli, P.; Bonoldi, I.; Yung, A.R.; Borgwardt, S.; Kempton, M.J.; Valmaggia, L.; Barale, F.; Caverzasi, E.; McGuire, P. Predicting psychosis: Meta-analysis of transition outcomes in individuals at high clinical risk. Arch. Gen. Psychiatry 2012, 69, 220–229. [Google Scholar] [CrossRef]
- Davison, J.; O’Gorman, A.; Brennan, L.; Cotter, D.R. A systematic review of metabolite biomarkers of schizophrenia. Schizophr. Res. 2018, 195, 32–50. [Google Scholar] [CrossRef]
- Zeisel, S.H.; Da Costa, K.A.; Franklin, P.D.; Alexander, E.A.; Lamont, J.T.; Sheard, N.F.; Beiser, A. Choline, an essential nutrient for humans. FASEB J. 1991, 5, 2093–2098. [Google Scholar] [CrossRef]
- Podo, F. Tumour phospholipid metabolism. NMR Biomed. Int. J. Devoted Dev. Appl. Magn. Reson. Vivo 1999, 12, 413–439. [Google Scholar] [CrossRef]
- Smucny, J.; Carter, C.S.; Maddock, R.J. Magnetic resonance spectroscopic evidence of increased choline in the dorsolateral prefrontal and visual cortices in recent onset schizophrenia. Neurosci. Lett. 2022, 770, 136410. [Google Scholar] [CrossRef]
- Kirtaş, D.; Karadağ, R.F.; Şengül, M.C.B.; Kiroğlu, Y. 1H-magnetic resonance spectroscopy in first episode and chronic schizophrenia patients. Turk. J. Med. Sci. 2016, 46, 862–871. [Google Scholar] [CrossRef]
- Bustillo, J.R.; Rowland, L.M.; Lauriello, J.; Petropoulos, H.; Hammond, R.; Hart, B.; Brooks, W.M. High choline concentrations in the caudate nucleus in antipsychotic-naive patients with schizophrenia. Am. J. Psychiatry 2002, 159, 130–133. [Google Scholar] [CrossRef]
- Yasukawa, R.; Miyaoka, T.; Mizuno, S.; Inagaki, T.; Horiguchi, J.; Oda, K.; Kitagaki, H. Proton magnetic resonance spectroscopy of the anterior cingulate gyrus, insular cortex and thalamus in schizophrenia associated with idiopathic unconjugated hyperbilirubinemia (Gilbert’s syndrome). J. Psychiatry Neurosci. 2005, 30, 416–422. [Google Scholar] [PubMed]
- Wang, Y.-m.; Xiao, Y.-h.; Xie, W.-l. Metabolite abnormalities in psychosis risk: A meta-analysis of proton magnetic resonance spectroscopy studies. Asian J. Psychiatry 2020, 54, 102220. [Google Scholar] [CrossRef]
- Byun, M.S.; Choi, J.-S.; Yoo, S.Y.; Kang, H.; Choi, C.-H.; Jang, N.P.; Jung, W.H.; Jung, M.H.; Jang, J.H.; Lee, J.-M.; et al. Depressive Symptoms and Brain Metabolite Alterations in Subjects at Ultra-high Risk for Psychosis: A Preliminary Study. Psychiatry Investig. 2009, 6, 264–271. [Google Scholar] [CrossRef] [PubMed]
- Wood, S.J.; Berger, G.; Velakoulis, D.; Phillips, L.J.; McGorry, P.D.; Yung, A.R.; Desmond, P.; Pantelis, C. Proton Magnetic Resonance Spectroscopy in First Episode Psychosis and Ultra High-Risk Individuals. Schizophr. Bull. 2003, 29, 831–843. [Google Scholar] [CrossRef]
- Uhl, I.; Mavrogiorgou, P.; Norra, C.; Forstreuter, F.; Scheel, M.; Witthaus, H.; Özgürdal, S.; Gudlowski, Y.; Bohner, G.; Gallinat, J.; et al. 1H-MR spectroscopy in ultra-high risk and first episode stages of schizophrenia. J. Psychiatr. Res. 2011, 45, 1135–1139. [Google Scholar] [CrossRef]
- Jessen, F.; Scherk, H.; Träber, F.; Theyson, S.; Berning, J.; Tepest, R.; Falkai, P.; Schild, H.-H.; Maier, W.; Wagner, M.; et al. Proton magnetic resonance spectroscopy in subjects at risk for schizophrenia. Schizophr. Res. 2006, 87, 81–88. [Google Scholar] [CrossRef]
- He, Y.; Kosciolek, T.; Tang, J.; Zhou, Y.; Li, Z.; Ma, X.; Zhu, Q.; Yuan, N.; Yuan, L.; Li, C.; et al. Gut microbiome and magnetic resonance spectroscopy study of subjects at ultra-high risk for psychosis may support the membrane hypothesis. Eur. Psychiatry 2018, 53, 37–45. [Google Scholar] [CrossRef] [PubMed]
- Romeo, B.; Petillion, A.; Martelli, C.; Benyamina, A. Magnetic resonance spectroscopy studies in subjects with high risk for psychosis: A meta-analysis and review. J. Psychiatr. Res. 2020, 125, 52–65. [Google Scholar] [CrossRef] [PubMed]
- Inglese, M.; Rusinek, H.; George, I.C.; Babb, J.S.; Grossman, R.I.; Gonen, O. Global average gray and white matter N-acetylaspartate concentration in the human brain. Neuroimage 2008, 41, 270–276. [Google Scholar] [CrossRef]
- Steel, A.; Mikkelsen, M.; Edden, R.A.; Robertson, C.E. Regional balance between glutamate+glutamine and GABA+ in the resting human brain. Neuroimage 2020, 220, 117112. [Google Scholar] [CrossRef]
- Bustillo, J.R.; Mayer, E.G.; Upston, J.; Jones, T.; Garcia, C.; Sheriff, S.; Maudsley, A.; Tohen, M.; Gasparovic, C.; Lenroot, R. Increased glutamate plus glutamine in the right middle cingulate in early schizophrenia but not in bipolar psychosis: A whole brain 1H-MRS study. Front. Psychiatry 2021, 12, 660850. [Google Scholar] [CrossRef]
- Schwerk, A.; Alves, F.D.S.; Pouwels, P.J.W.; Van Amelsvoort, T. Metabolic alterations associated with schizophrenia: A critical evaluation of proton magnetic resonance spectroscopy studies. J. Neurochem. 2013, 128, 1–87. [Google Scholar] [CrossRef]
- Whitehurst, T.S.; Osugo, M.; Townsend, L.; Shatalina, E.; Vava, R.; Onwordi, E.C.; Howes, O. Proton Magnetic Resonance Spectroscopy of N-acetyl Aspartate in Chronic Schizophrenia, First Episode of Psychosis and High-Risk of Psychosis: A Systematic Review and Meta-Analysis. Neurosci. Biobehav. Rev. 2020, 119, 255–267. [Google Scholar] [CrossRef]
- Smesny, S.; Berberich, D.; Gussew, A.; Schönfeld, N.; Langbein, K.; Walther, M.; Reichenbach, J.R. Alterations of neurometabolism in the dorsolateral prefrontal cortex and thalamus in transition to psychosis patients change under treatment as usual—A two years follow-up 1H/31P-MR-spectroscopy study. Schizophr. Res. 2021, 228, 7–18. [Google Scholar] [CrossRef]
- Brugger, S.; Davis, J.M.; Leucht, S.; Stone, J.M. Proton Magnetic Resonance Spectroscopy and Illness Stage in Schizophrenia—A Systematic Review and Meta-Analysis. Biol. Psychiatry 2011, 69, 495–503. [Google Scholar] [CrossRef] [PubMed]
- Smesny, S.; Gussew, A.; Schack, S.; Langbein, K.; Wagner, G.; Reichenbach, J.R. Neurometabolic patterns of an “at risk for mental disorders” syndrome involve abnormalities in the thalamus and anterior midcingulate cortex. Schizophr. Res. 2022, 243, 285–295. [Google Scholar] [CrossRef] [PubMed]
- Mondino, M.; Brunelin, J.; Saoud, M. N-Acetyl-Aspartate Level is Decreased in the Prefrontal Cortex in Subjects At-Risk for Schizophrenia. Front. Psychiatry 2013, 4, 99. [Google Scholar] [CrossRef]
- Liemburg, E.; Sibeijn-Kuiper, A.; Bais, L.; Pijnenborg, G.; Knegtering, H.; van der Velde, J.; Opmeer, E.; de Vos, A.; Lange, J.D.-D.; Wunderink, L.; et al. Prefrontal NAA and Glx Levels in Different Stages of Psychotic Disorders: A 3T 1H-MRS Study. Sci. Rep. 2016, 6, 21873. [Google Scholar] [CrossRef] [PubMed]
- Mayeli, A.; Sonnenschein, S.F.; Yushmanov, V.E.; Wilson, J.D.; Blazer, A.; Foran, W.; Perica, M.; Calabro, F.J.; Luna, B.; Hetherington, H.P.; et al. Dorsolateral Prefrontal Cortex Glutamate/Gamma-Aminobutyric Acid (GABA) Alterations in Clinical High Risk and First-Episode Schizophrenia: A Preliminary 7-T Magnetic Resonance Spectroscopy Imaging Study. Int. J. Mol. Sci. 2022, 23, 15846. [Google Scholar] [CrossRef]
- Sonnenschein, S.F.; Mayeli, A.; Yushmanov, V.E.; Blazer, A.; Calabro, F.J.; Perica, M.; Foran, W.; Luna, B.; Hetherington, H.P.; Ferrarelli, F.; et al. A longitudinal investigation of GABA, glutamate, and glutamine across the insula during antipsychotic treatment of first-episode schizophrenia. Schizophr. Res. 2022, 248, 98–106. [Google Scholar] [CrossRef]
- Zhu, H.; Barker, P.B. MR Spectroscopy and Spectroscopic Imaging of the Brain. Magn. Reson. Neuroimaging 2011, 711, 203–226. [Google Scholar]
- Van Zijl, P.C.; Barker, P.B. Magnetic Resonance Spectroscopy and Spectroscopic Imaging for the Study of Brain Metabolism a. Ann. N. Y. Acad. Sci. 1997, 820, 75–96. [Google Scholar] [CrossRef] [PubMed]
- Quiñones, G.M.; Mayeli, A.; Yushmanov, V.E.; Hetherington, H.P.; Ferrarelli, F. Reduced GABA/glutamate in the thalamus of individuals at clinical high risk for psychosis. Neuropsychopharmacology 2021, 46, 1133–1139. [Google Scholar] [CrossRef] [PubMed]
- Balchandani, P.; Naidich, T. Ultra-high-field MR neuroimaging. Am. J. Neuroradiol. 2015, 36, 1204–1215. [Google Scholar] [CrossRef] [PubMed]
- Younis, S.; Hougaard, A.; Christensen, C.E.; Vestergaard, M.B.; Petersen, E.T.; Boer, V.O.; Paulson, O.B.; Ashina, M.; Marsman, A.; Larsson, H.B.W. Feasibility of Glutamate and GABA Detection in Pons and Thalamus at 3T and 7T by Proton Magnetic Resonance Spectroscopy. Front. Neurosci. 2020, 14, 559314. [Google Scholar] [CrossRef]
- Terpstra, M.; Cheong, I.; Lyu, T.; Deelchand, D.K.; Emir, U.E.; Bednařík, P.; Eberly, L.E.; Öz, G. Test-retest reproducibility of neurochemical profiles with short-echo, single-voxel MR spectroscopy at 3T and 7T. Magn. Reson. Med. 2016, 76, 1083–1091. [Google Scholar] [CrossRef] [PubMed]
- Natsubori, T.; Inoue, H.; Abe, O.; Takano, Y.; Iwashiro, N.; Aoki, Y.; Koike, S.; Yahata, N.; Katsura, M.; Gonoi, W.; et al. Reduced frontal glutamate + glutamine and N-acetylaspartate levels in patients with chronic schizophrenia but not in those at clinical high risk for psychosis or with first-episode schizophrenia. Schizophr. Bull. 2014, 40, 1128–1139. [Google Scholar] [CrossRef]
- De la Fuente-Sandoval, C.; Reyes-Madrigal, F.; Mao, X.; León-Ortiz, P.; Rodríguez-Mayoral, O.; Solís-Vivanco, R.; Favila, R.; Graff-Guerrero, A.; Shungu, D.C. Cortico-striatal GABAergic and glutamatergic dysregulations in subjects at ultra-high risk for psychosis investigated with proton magnetic resonance spectroscopy. Int. J. Neuropsychopharmacol. 2016, 19, pyv105. [Google Scholar] [CrossRef]
- Stone, J.M.; Day, F.; Tsagaraki, H.; Valli, I.; McLean, M.A.; Lythgoe, D.J.; O’Gorman, R.L.; Barker, G.J.; McGuire, P.K.; OASIS. Glutamate Dysfunction in People with Prodromal Symptoms of Psychosis: Relationship to Gray Matter Volume. Biol. Psychiatry 2009, 66, 533–539. [Google Scholar] [CrossRef]
- Weber-Fahr, W.; Englisch, S.; Esser, A.; Tunc-Skarka, N.; Meyer-Lindenberg, A.; Ende, G.; Zink, M. Altered phospholipid metabolism in schizophrenia: A phosphorus 31 nuclear magnetic resonance spectroscopy study. Psychiatry Res. 2013, 214, 365–373. [Google Scholar] [CrossRef]
- Wang, M.; Barker, P.B.; Cascella, N.G.; Coughlin, J.M.; Nestadt, G.; Nucifora, F.C.; Sedlak, T.W.; Kelly, A.; Younes, L.; Geman, D.; et al. Longitudinal changes in brain metabolites in healthy controls and patients with first episode psychosis: A 7-Tesla MRS study. Mol. Psychiatry 2023. [Google Scholar] [CrossRef]
- Tarumi, R.; Tsugawa, S.; Noda, Y.; Plitman, E.; Honda, S.; Matsushita, K.; Chavez, S.; Sawada, K.; Wada, M.; Matsui, M.; et al. Levels of glutamatergic neurometabolites in patients with severe treatment-resistant schizophrenia: A proton magnetic resonance spectroscopy study. Neuropsychopharmacology 2020, 45, 632–640. [Google Scholar] [CrossRef]
- Öngür, D. Magnetic Resonance Spectroscopy Studies in Schizophrenia. In Neuroimaging in Schizophrenia; Springer: Cham, Switzerland, 2020; pp. 179–189. [Google Scholar]
- Paslakis, G.; Träber, F.; Roberz, J.; Block, W.; Jessen, F. N-acetyl-aspartate (NAA) as a correlate of pharmacological treatment in psychiatric disorders: A systematic review. Eur. Neuropsychopharmacol. 2014, 24, 1659–1675. [Google Scholar] [CrossRef] [PubMed]
- Huang, M.; Guo, W.; Lu, S.; Pan, F.; Chen, J.; Hu, J.; Hu, S.; Xu, W.; Shang, D.; Xu, Y. The relationship between the alterations in metabolite levels in the dorsolateral prefrontal cortex and clinical symptoms of patients with first-episode schizophrenia: A one year follow-up study. Oncotarget 2019, 10, 606–615. [Google Scholar] [CrossRef]
- Pan, J.W.; Lo, K.-M.; Hetherington, H.P. Role of very high order and degree B 0 shimming for spectroscopic imaging of the human brain at 7 tesla. Magn. Reson. Med. 2011, 68, 1007–1017. [Google Scholar] [CrossRef]
- Pan, J.; Avdievich, N.; Hetherington, H. J-refocused coherence transfer spectroscopic imaging at 7 T in human brain. Magn. Reson. Med. 2010, 64, 1237–1246. [Google Scholar] [CrossRef]
- Pradhan, S.; Bonekamp, S.; Gillen, J.S.; Rowland, L.M.; Wijtenburg, S.A.; Edden, R.A.; Barker, P.B. Comparison of single voxel brain MRS AT 3T and 7T using 32-channel head coils. Magn. Reson. Imaging 2015, 33, 1013–1018. [Google Scholar] [CrossRef]
- Marques, J.P.; Kober, T.; Krueger, G.; van der Zwaag, W.; Van de Moortele, P.-F.; Gruetter, R. MP2RAGE, a self bias-field corrected sequence for improved segmentation and T1-mapping at high field. Neuroimage 2010, 49, 1271–1281. [Google Scholar] [CrossRef] [PubMed]
- Provencher, S.W. Automatic quantitation of localized in vivo 1H spectra with LCModel. NMR Biomed. Int. J. Devoted Dev. Appl. Magn. Reson. Vivo 2001, 14, 260–264. [Google Scholar]
- Smith, S.; Levante, T.; Meier, B.; Ernst, R. Computer Simulations in Magnetic Resonance. An Object-Oriented Programming Approach. J. Magn. Reson. Ser. A 1994, 106, 75–105. [Google Scholar] [CrossRef]
- Faul, F.; Erdfelder, E.; Lang, A.-G.; Buchner, A. G* Power 3: A flexible statistical power analysis program for the social, behavioral, and biomedical sciences. Behav. Res. Methods 2007, 39, 175–191. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Measures | Healthy Control | Clinical High Risk | p-Value |
|---|---|---|---|
| Number of Subjects | 32 | 26 | - |
| Sex (# female) | 18 | 15 | 0.912 1 |
| Average Age ± SD in Years (range) | 21.48 ± 4.98 (14–33) | 19.93 ± 3.16 (15–27) | 0.172 2 |
| SOPS Positive Symptoms (average ± SD) | - | 12.19 ± 3.10 | - |
| SOPS Negative Symptoms (average ± SD) | - | 11.60 ± 4.34 | - |
| SOPS Disorganized Symptoms (average ± SD) | - | 5.00 ± 1.94 | - |
| SOPS General Symptoms (average ± SD) | - | 7.68 ± 2.56 | - |
| tCho/Cre | tNAA/Cre | ||||
|---|---|---|---|---|---|
| F-Stat | p-Value | F-Stat | p-Value | ||
| Right DLPFC | Group | 1.086 | 0.302 | 4.038 | 0.049 |
| Age | 0.692 | 0.410 | 0.048 | 0.827 | |
| Sex | 0.268 | 0.607 | <0.001 | 0.999 | |
| GM% | 0.058 | 0.811 | 1.746 | 0.192 | |
| WM% | 3.330 | 0.074 | 2.362 | 0.131 | |
| Left DLPFC | Group | 0.436 | 0.513 | 2.070 | 0.158 |
| Age | 2.059 | 0.156 | 0.905 | 0.347 | |
| Sex | 0.003 | 0.954 | 0.064 | 0.802 | |
| GM% | 0.170 | 0.683 | 0.317 | 0.577 | |
| WM% | 2.280 | 0.139 | 0.400 | 0.531 | |
| ACC | Group | 0.227 | 0.636 | 0.253 | 0.617 |
| Age | 2.557 | 0.116 | 0.021 | 0.887 | |
| Sex | 8.289 | 0.006 | 0.419 | 0.520 | |
| GM% | 0.144 | 0.706 | 0.099 | 0.755 | |
| WM% | 2.265 | 0.138 | <0.001 | 0.993 | |
| Right Caudate | Group | 0.723 | 0.399 | 1.584 | 0.214 |
| Age | 0.864 | 0.357 | 1.290 | 0.261 | |
| Sex | 6.758 | 0.012 | 0.970 | 0.329 | |
| GM% | 0.075 | 0.785 | 0.035 | 0.852 | |
| WM% | 0.080 | 0.778 | 2.907 | 0.094 | |
| Left Caudate | Group | 0.319 | 0.575 | 0.115 | 0.736 |
| Age | 0.280 | 0.599 | 0.072 | 0.790 | |
| Sex | 2.121 | 0.152 | 0.816 | 0.371 | |
| GM% | 0.523 | 0.473 | 0.326 | 0.571 | |
| WM% | <0.001 | 0.996 | 0.702 | 0.406 | |
| Right Insula | Group | 0.110 | 0.742 | 0.048 | 0.826 |
| Age | 0.051 | 0.822 | 0.095 | 0.759 | |
| Sex | 6.390 | 0.015 | 1.703 | 0.198 | |
| GM% | 2.047 | 0.159 | 0.317 | 0.576 | |
| WM% | 0.007 | 0.931 | 0.428 | 0.516 | |
| Left Insula | Group | 0.413 | 0.523 | 0.173 | 0.679 |
| Age | 0.557 | 0.459 | 0.184 | 0.970 | |
| Sex | 2.735 | 0.104 | 3.316 | 0.074 | |
| GM% | 10.045 | 0.003 | 4.232 | 0.045 | |
| WM% | 0.002 | 0.967 | 1.401 | 0.242 | |
| Right Thalamus | Group | 0.525 | 0.472 | 0.351 | 0.556 |
| Age | 0.558 | 0.458 | 0.269 | 0.606 | |
| Sex | 10.316 | 0.002 | 0.449 | 0.506 | |
| GM% | 0.166 | 0.685 | 0.042 | 0.839 | |
| WM% | 1.960 | 0.168 | 0.003 | 0.953 | |
| Left Thalamus | Group | 0.523 | 0.473 | 0.926 | 0.341 |
| Age | 0.182 | 0.671 | 0.001 | 0.974 | |
| Sex | 6.227 | 0.016 | 0.917 | 0.172 | |
| GM% | 2.568 | 0.115 | 0.341 | 0.562 | |
| WM% | 2.718 | 0.106 | 0.356 | 0.131 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mayeli, A.; Janssen, S.A.; Huston, C.A.; Rupp, J.S.; Sharma, K.; Moon, C.-H.; Keihani, A.; Hetherington, H.P.; Ferrarelli, F. N-Acetylaspartate and Choline Metabolites in Cortical and Subcortical Regions in Clinical High Risk Relative to Healthy Control Subjects: An Exploratory 7T MRSI Study. Int. J. Mol. Sci. 2023, 24, 7682. https://doi.org/10.3390/ijms24097682
Mayeli A, Janssen SA, Huston CA, Rupp JS, Sharma K, Moon C-H, Keihani A, Hetherington HP, Ferrarelli F. N-Acetylaspartate and Choline Metabolites in Cortical and Subcortical Regions in Clinical High Risk Relative to Healthy Control Subjects: An Exploratory 7T MRSI Study. International Journal of Molecular Sciences. 2023; 24(9):7682. https://doi.org/10.3390/ijms24097682
Chicago/Turabian StyleMayeli, Ahmad, Sabine A. Janssen, Chloe A. Huston, Julia S. Rupp, Kamakashi Sharma, Chan-Hong Moon, Ahmadreza Keihani, Hoby P. Hetherington, and Fabio Ferrarelli. 2023. "N-Acetylaspartate and Choline Metabolites in Cortical and Subcortical Regions in Clinical High Risk Relative to Healthy Control Subjects: An Exploratory 7T MRSI Study" International Journal of Molecular Sciences 24, no. 9: 7682. https://doi.org/10.3390/ijms24097682