

LF Power of HRV Could Be the Piezo2 Activity Level in Baroreceptors with Some Piezo1 Residual Activity Contribution

{kind=link}
Abstract
:1. Introduction
2. Piezo Ion Channels and Baroreceptors
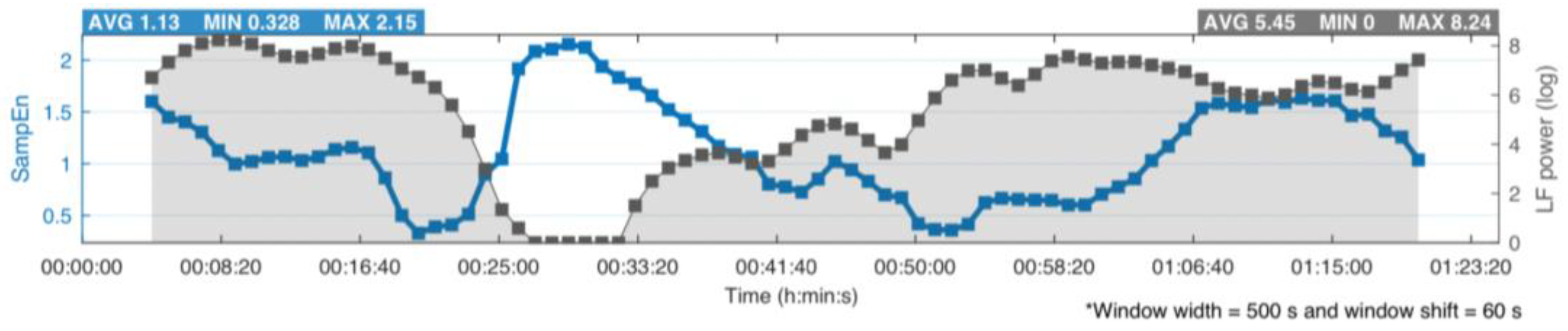
3. LF Power of HRV
4. Proprioceptive vs. Piezo System
5. Conclusions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Goldstein, D.S.; Bentho, O.; Park, M.-Y.; Sharabi, Y. Low-frequency power of heart rate variability is not a measure of cardiac sympathetic tone but may be a measure of modulation of cardiac autonomic outflows by baroreflexes. Exp. Physiol. 2011, 96, 1255–1261. [Google Scholar] [CrossRef] [PubMed]
- Stanley, J.; Peake, J.M.; Buchheit, M. Cardiac Parasympathetic Reactivation Following Exercise: Implications for Training Prescription. Sports Med. 2013, 43, 1259–1277. [Google Scholar] [CrossRef] [PubMed]
- Sonkodi, B.; Kopa, Z.; Nyirády, P. Post Orgasmic Illness Syndrome (POIS) and Delayed Onset Muscle Soreness (DOMS): Do They Have Anything in Common? Cells 2021, 10, 1867. [Google Scholar] [CrossRef] [PubMed]
- Sonkodi, B. Delayed Onset Muscle Soreness and Critical Neural Microdamage-Derived Neuroinflammation. Biomolecules 2022, 12, 1207. [Google Scholar] [CrossRef]
- Perini, R.; Veicsteinas, A. Heart rate variability and autonomic activity at rest and during exercise in various physiological conditions. Eur. J. Appl. Physiol. 2003, 90, 317–325. [Google Scholar] [CrossRef]
- Saul, J. Beat-To-Beat Variations of Heart Rate Reflect Modulation of Cardiac Autonomic Outflow. Physiology 1990, 5, 32–37. [Google Scholar] [CrossRef]
- Coste, B.; Xiao, B.; Santos, J.S.; Syeda, R.; Grandl, J.; Spencer, K.S.; Kim, S.E.; Schmidt, M.; Mathur, J.; Dubin, A.E.; et al. Piezo proteins are pore-forming subunits of mechanically activated channels. Nature 2012, 483, 176–181. [Google Scholar] [CrossRef] [Green Version]
- Zeng, W.-Z.; Marshall, K.L.; Min, S.; Daou, I.; Chapleau, M.W.; Abboud, F.M.; Liberles, S.D.; Patapoutian, A. PIEZOs mediate neuronal sensing of blood pressure and the baroreceptor reflex. Science 2018, 362, 464–467. [Google Scholar] [CrossRef] [Green Version]
- Rowell, L.B.; O’Leary, D.S. Reflex control of the circulation during exercise: Chemoreflexes and mechanoreflexes. J. Appl. Physiol. 1990, 69, 407–418. [Google Scholar] [CrossRef] [Green Version]
- O’Leary, D.S. Heart rate control during exercise by baroreceptors and skeletal muscle afferents. Med. Sci. Sports Exerc. 1996, 28, 210–217. [Google Scholar] [CrossRef]
- Burke, S.D.; Jordan, J.; Harrison, D.G.; Karumanchi, S.A. Solving Baroreceptor Mystery: Role of PIEZO Ion Channels. J. Am. Soc. Nephrol. 2019, 30, 911–913. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Woo, S.-H.; Lukacs, V.; De Nooij, J.C.; Zaytseva, D.; Criddle, C.R.; Francisco, A.G.; Jessell, T.M.; Wilkinson, K.A.; Patapoutian, A. Piezo2 is the principal mechanotransduction channel for proprioception. Nat. Neurosci. 2015, 18, 1756–1762. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Suchyna, T.M. Piezo channels and GsMTx4: Two milestones in our understanding of excitatory mechanosensitive channels and their role in pathology. Prog. Biophys. Mol. Biol. 2017, 130, 244–253. [Google Scholar] [CrossRef] [PubMed]
- Volkers, L.; Mechioukhi, Y.; Coste, B. Piezo channels: From structure to function. Pflugers. Arch. 2015, 467, 95–99. [Google Scholar] [CrossRef] [PubMed]
- Guo, Y.R.; MacKinnon, R. Structure-based membrane dome mechanism for Piezo mechanosensitivity. eLife 2017, 6, e33660. [Google Scholar] [CrossRef]
- Coste, B.; Mathur, J.; Schmidt, M.; Earley, T.J.; Ranade, S.; Petrus, M.J.; Dubin, A.E.; Patapoutian, A. Piezo1 and Piezo2 Are Essential Components of Distinct Mechanically Activated Cation Channels. Science 2010, 330, 55–60. [Google Scholar] [CrossRef] [Green Version]
- Gottlieb, P.A.; Sachs, F. Piezo1: Properties of a cation selective mechanical channel. Channels 2012, 6, 214–219. [Google Scholar] [CrossRef]
- Szczot, M.; Nickolls, A.R.; Lam, R.M.; Chesler, A.T. The Form and Function of PIEZO2. Annu. Rev. Biochem. 2021, 90, 507–534. [Google Scholar] [CrossRef]
- Ranade, S.S.; Woo, S.-H.; Dubin, A.E.; Moshourab, R.A.; Wetzel, C.; Petrus, M.; Mathur, J.; Bégay, V.; Coste, B.; Mainquist, J.; et al. Piezo2 is the major transducer of mechanical forces for touch sensation in mice. Nature 2014, 516, 121–125. [Google Scholar] [CrossRef] [Green Version]
- Woo, S.-H.; Ranade, S.; Weyer, A.D.; Dubin, A.E.; Baba, Y.; Qiu, Z.; Petrus, M.; Miyamoto, T.; Reddy, K.; Lumpkin, E.A.; et al. Piezo2 is required for Merkel-cell mechanotransduction. Nature 2014, 509, 622–626. [Google Scholar] [CrossRef] [Green Version]
- Martins, J.R.; Penton, D.; Peyronnet, R.; Arhatte, M.; Moro, C.; Picard, N.; Kurt, B.; Patel, A.; Honoré, E.; Demolombe, S. Piezo1-dependent regulation of urinary osmolarity. Pflugers Arch 2016, 468, 1197–1206. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.; Liu, S.; Liu, H.; Ru, K.; Jia, Y.; Wu, Z.; Liang, S.; Khan, Z.; Chen, Z.; Qian, A.; et al. Piezo Channels: Awesome Mechanosensitive Structures in Cellular Mechanotransduction and Their Role in Bone. Int. J. Mol. Sci. 2021, 22, 6429. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Hou, B.; Tumova, S.; Muraki, K.; Bruns, A.; Ludlow, M.J.; Sedo, A.; Hyman, A.J.; McKeown, L.; Young, R.S.; et al. Piezo1 integration of vascular architecture with physiological force. Nature 2014, 515, 279–282. [Google Scholar] [CrossRef]
- Sonkodi, B.; Resch, M.D.; Hortobágyi, T. Is the Sex Difference a Clue to the Pathomechanism of Dry Eye Disease? Watch out for the NGF-TrkA-Piezo2 Signaling Axis and the Piezo2 Channelopathy. J. Mol. Neurosci. 2022. [Google Scholar] [CrossRef] [PubMed]
- Ghitani, N.; Chesler, A.T. The anatomy of the baroreceptor reflex. Cell Rep. 2019, 29, 2121–2122. [Google Scholar] [CrossRef] [Green Version]
- Min, S.; Chang, R.B.; Prescott, S.L.; Beeler, B.; Joshi, N.R.; Strochlic, D.E.; Liberles, S.D. Arterial Baroreceptors Sense Blood Pressure through Decorated Aortic Claws. Cell Rep. 2019, 29, 2192–2201. [Google Scholar] [CrossRef] [Green Version]
- Rutlin, M.; Ho, C.-Y.; Abraira, V.E.; Cassidy, C.; Bai, L.; Woodbury, C.J.; Ginty, D.D. The Cellular and Molecular Basis of Direction Selectivity of Aδ-LTMRs. Cell 2014, 159, 1640–1651. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Hamill, O.P. Piezo2-peripheral baroreceptor channel expressed in select neurons of the mouse brain: A putative mechanism for synchronizing neural networks by transducing intracranial pressure pulses. J. Integr. Neurosci. 2021, 20, 825–837. [Google Scholar] [CrossRef]
- Greitz, D.; Wirestam, R.; Franck, A.; Nordell, B.; Thomsen, C. Pulsatile brain movement and associated hydrodynamics studied by magnetic resonance phase imaging. Neuroradiology 1992, 34, 370–380. [Google Scholar] [CrossRef]
- Graves, A.R.; Moore, S.J.; Bloss, E.B.; Mensh, B.D.; Kath, W.L.; Spruston, N. Hippocampal Pyramidal Neurons Comprise Two Distinct Cell Types that Are Countermodulated by Metabotropic Receptors. Neuron 2012, 76, 776–789. [Google Scholar] [CrossRef] [Green Version]
- Wittner, L.; Miles, R. Factors defining a pacemaker region for synchrony in the hippocampus. J. Physiol. 2007, 584, 867–883. [Google Scholar] [CrossRef] [PubMed]
- Mangoni, M.E.; Couette, B.; Bourinet, E.; Platzer, J.; Reimer, D.; Striessnig, J.; Nargeot, J. Functional role of L-type Cav1.3 Ca2+ channels in cardiac pacemaker activity. Proc. Natl. Acad. Sci. USA 2003, 100, 5543–5548. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cohen, M.A.; Taylor, J.A. Short-term cardiovascular oscillations in man: Measuring and modelling the physiologies. J. Physiol. 2002, 542, 669–683. [Google Scholar] [CrossRef]
- Bernardi, L.; Keller, F.; Sanders, M.; Reddy, P.S.; Griffith, B.; Meno, F.; Pinsky, M.R. Respiratory sinus arrhythmia in the denervated human heart. J. Appl. Physiol. 1989, 67, 1447–1455. [Google Scholar] [CrossRef] [PubMed]
- Shin, S.M.; Moehring, F.; Itson-Zoske, B.; Fan, F.; Stucky, C.L.; Hogan, Q.H.; Yu, H. Piezo2 mechanosensitive ion channel is located to sensory neurons and nonneuronal cells in rat peripheral sensory pathway: Implications in pain. Pain 2021, 162, 2750–2768. [Google Scholar] [CrossRef] [PubMed]
- Bainbridge, F.A. The relation between respiration and the pulse-rate. J. Physiol. 1920, 54, 192–202. [Google Scholar] [CrossRef]
- Schwartzman, R.J.; Kerrigan, J. The movement disorder of reflex sympathetic dystrophy. Neurology 1990, 40, 57. [Google Scholar] [CrossRef]
- Hellström, F.; Roatta, S.; Thunberg, J.; Passatore, M.; Djupsjöbacka, M. Responses of muscle spindles in feline dorsal neck muscles to electrical stimulation of the cervical sympathetic nerve. Exp. Brain Res. 2005, 165, 328–342. [Google Scholar] [CrossRef]
- Roatta, S.; Windhorst, U.; Ljubisavljevic, M.; Johansson, H.; Passatore, M. Sympathetic modulation of muscle spindle afferent sensitivity to stretch in rabbit jaw closing muscles. J. Physiol. 2002, 540, 237–248. [Google Scholar] [CrossRef] [Green Version]
- Than, K.; Kim, E.; Navarro, C.; Chu, S.; Klier, N.; Occiano, A.; Ortiz, S.; Salazar, A.; Valdespino, S.R.; Villegas, N.K.; et al. Vesicle-released glutamate is necessary to maintain muscle spindle afferent excitability but not dynamic sensitivity in adult mice. J. Physiol. 2021, 599, 2953–2967. [Google Scholar] [CrossRef]
- Bewick, G.S.; Banks, R.W. Spindles are doin’ it for themselves: Glutamatergic autoexcitation in muscle spindles. J. Physiol. 2021, 599, 2781–2783. [Google Scholar] [CrossRef] [PubMed]
- Sonkodi, B.; Hegedűs, Á.; Kopper, B.; Berkes, I. Significantly Delayed Medium-Latency Response of the Stretch Reflex in Delayed-Onset Muscle Soreness of the Quadriceps Femoris Muscles Is Indicative of Sensory Neuronal Microdamage. J. Funct. Morphol. Kinesiol. 2022, 7, 43. [Google Scholar] [CrossRef] [PubMed]
- Sonkodi, B.; Berkes, I.; Koltai, E. Have We Looked in the Wrong Direction for More Than 100 Years? Delayed Onset Muscle Soreness Is, in Fact, Neural Microdamage Rather Than Muscle Damage. Antioxidants 2020, 9, 212. [Google Scholar] [CrossRef] [Green Version]
- Wilkinson, K.A. Molecular determinants of mechanosensation in the muscle spindle. Curr. Opin. Neurobiol. 2022, 74, 102542. [Google Scholar] [CrossRef]
- Hody, S.; Croisier, J.-L.; Bury, T.; Rogister, B.; Leprince, P. Eccentric Muscle Contractions: Risks and Benefits. Front. Physiol. 2019, 10, 536. [Google Scholar] [CrossRef] [Green Version]
- Sonkodi, B.; Bardoni, R.; Hangody, L.; Radák, Z.; Berkes, I. Does Compression Sensory Axonopathy in the Proximal Tibia Contribute to Noncontact Anterior Cruciate Ligament Injury in a Causative Way?—A New Theory for the Injury Mechanism. Life 2021, 11, 443. [Google Scholar] [CrossRef] [PubMed]
- Perini, R.; Milesi, S.; Biancardi, L.; Pendergast, D.R.; Veicsteinas, A. Heart rate variability in exercising humans: Effect of water immersion. Eur. J. Appl. Physiol. 1998, 77, 326–332. [Google Scholar] [CrossRef] [PubMed]
- Sonkodi, B. Delayed Onset Muscle Soreness (DOMS): The Repeated Bout Effect and Chemotherapy-Induced Axonopathy May Help Explain the Dying-Back Mechanism in Amyotrophic Lateral Sclerosis and Other Neurodegenerative Diseases. Brain Sci. 2021, 11, 108. [Google Scholar] [CrossRef]
- Sonkodi, B.; Varga, E.; Hangody, L.; Poór, G.; Berkes, I. Finishing stationary cycling too early after anterior cruciate ligament reconstruction is likely to lead to higher failure. BMC Sports Sci. Med. Rehabil. 2021, 13, 149. [Google Scholar] [CrossRef]
- Kamath, M.V.; Fallen, E.L.; McKelvie, R. Effects of steady state exercise on the power spectrum of heart rate variability. Med. Sci. Sports Exerc. 1991, 23, 428–434. [Google Scholar] [CrossRef]
- Rode, B.; Shi, J.; Endesh, N.; Drinkhill, M.J.; Webster, P.J.; Lotteau, S.J.; Bailey, M.A.; Yuldasheva, N.Y.; Ludlow, M.J.; Cubbon, R.M.; et al. Piezo1 channels sense whole body physical activity to reset cardiovascular homeostasis and enhance performance. Nat. Commun. 2017, 8, 350. [Google Scholar] [CrossRef] [PubMed]
- Komka, Z.; Szilágyi, B.; Molnár, D.; Sipos, B.; Tóth, M.; Sonkodi, B.; Ács, P.; Elek, J.; Szász, M. Exercise-related hemoconcentration and hemodilution in hydrated and dehydrated athletes: An observational study of the Hungarian canoeists. PLoS ONE 2022, 17, e0277978. [Google Scholar] [CrossRef] [PubMed]
- Sonkodi, B. Psoriasis, Is It a Microdamage of Our “Sixth Sense”? A Neurocentric View. Int. J. Mol. Sci. 2022, 23, 11940. [Google Scholar] [CrossRef] [PubMed]
- Sonkodi, B.; Hortobágyi, T. Amyotrophic lateral sclerosis and delayed onset muscle soreness in light of the impaired blink and stretch reflexes – watch out for Piezo2. Open Med. 2022, 17, 397–402. [Google Scholar] [CrossRef]
- Sonkodi, B. Miswired Proprioception in Amyotrophic Lateral Sclerosis in Relation to Pain Sensation (and in Delayed Onset Muscle Soreness)—Is Piezo2 Channelopathy a Principal Transcription Activator in Proprioceptive Terminals Besides Being the Potential Primary Damage? Life 2023, 13, 657. [Google Scholar] [CrossRef]
- Baltadzhieva, R.; Gurevich, T.; Korczyn, A.D. Autonomic impairment in amyotrophic lateral sclerosis. Curr. Opin. Neurol. 2005, 18, 487–493. [Google Scholar] [CrossRef]
- Nagy, Z.F.; Sonkodi, B.; Pál, M.; Klivényi, P.; Széll, M. Likely Pathogenic Variants of Cav1.3 and Nav1.1 Encoding Genes in Amyotrophic Lateral Sclerosis Could Elucidate the Dysregulated Pain Pathways. Biomedicines 2023, 11, 933. [Google Scholar] [CrossRef]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sonkodi, B. LF Power of HRV Could Be the Piezo2 Activity Level in Baroreceptors with Some Piezo1 Residual Activity Contribution. Int. J. Mol. Sci. 2023, 24, 7038. https://doi.org/10.3390/ijms24087038
Sonkodi B. LF Power of HRV Could Be the Piezo2 Activity Level in Baroreceptors with Some Piezo1 Residual Activity Contribution. International Journal of Molecular Sciences. 2023; 24(8):7038. https://doi.org/10.3390/ijms24087038
Chicago/Turabian StyleSonkodi, Balázs. 2023. "LF Power of HRV Could Be the Piezo2 Activity Level in Baroreceptors with Some Piezo1 Residual Activity Contribution" International Journal of Molecular Sciences 24, no. 8: 7038. https://doi.org/10.3390/ijms24087038