

Meta-Analysis of Mechano-Sensitive Ion Channels in Human Hearts: Chamber- and Disease-Preferential mRNA Expression
 , ,
, ,
Abstract
:1. Introduction
2. Results
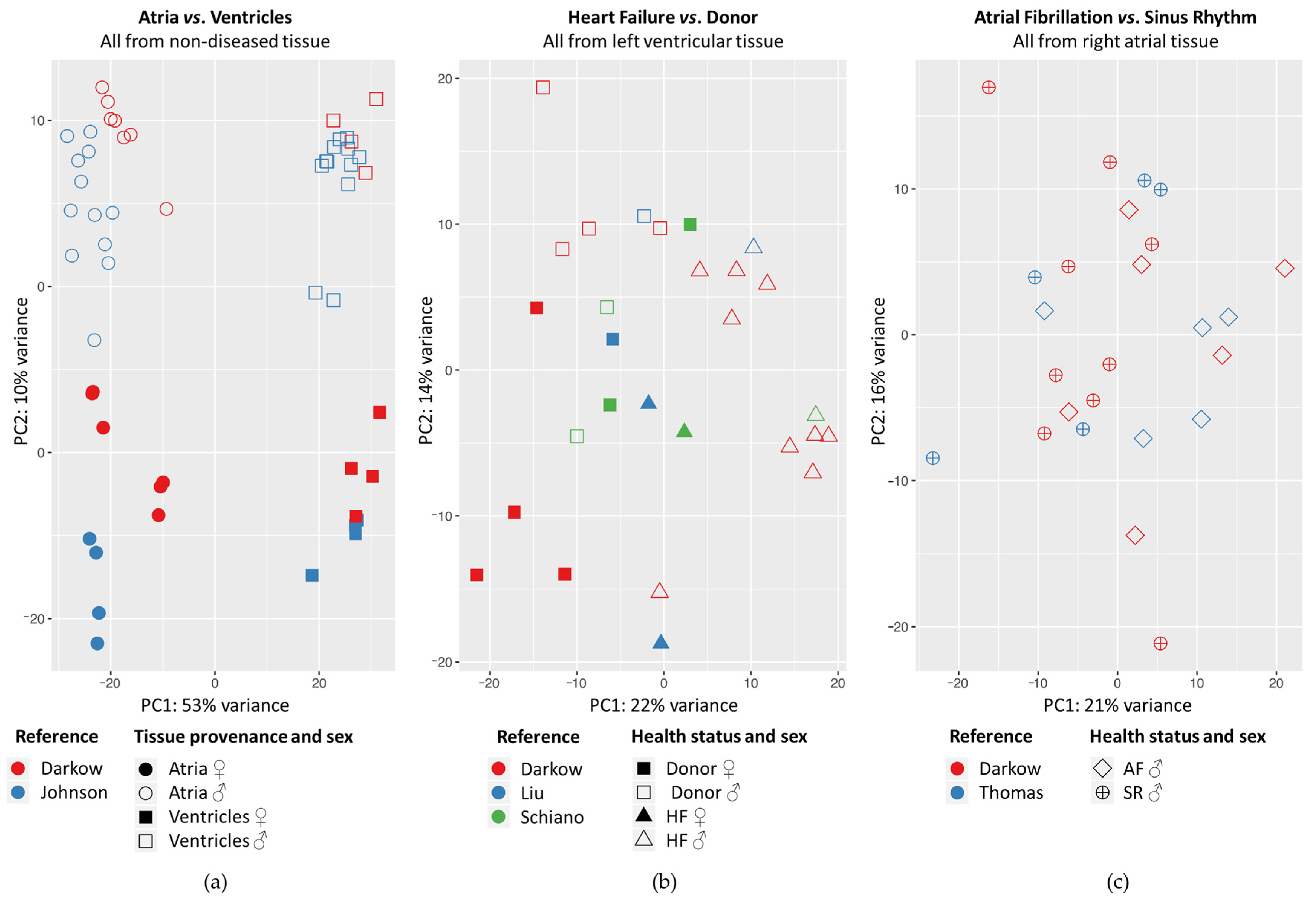
2.1. Principal Component Analysis
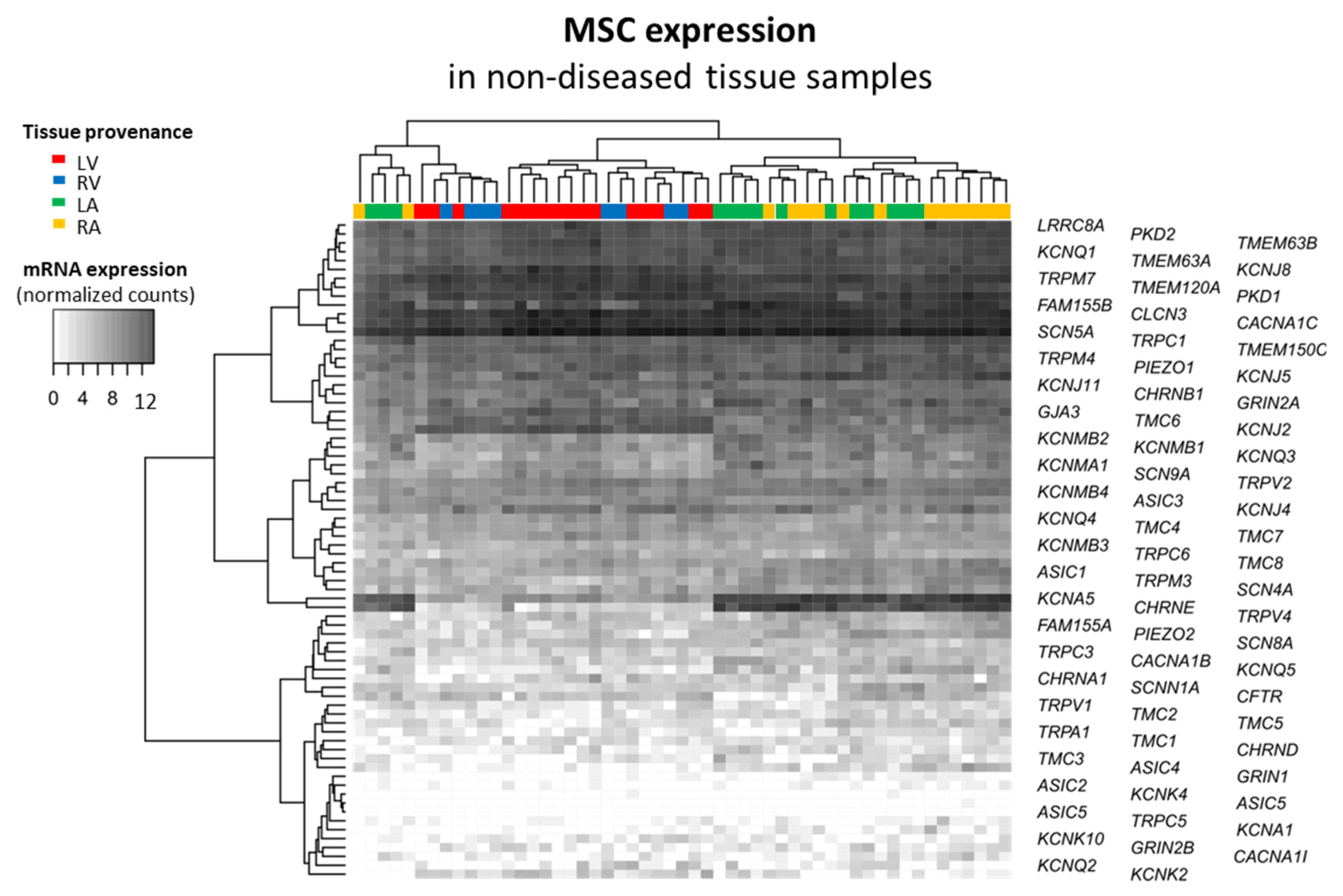
2.2. Mechano-Sensitive Ion Channels and Their Cardiac mRNA Expression
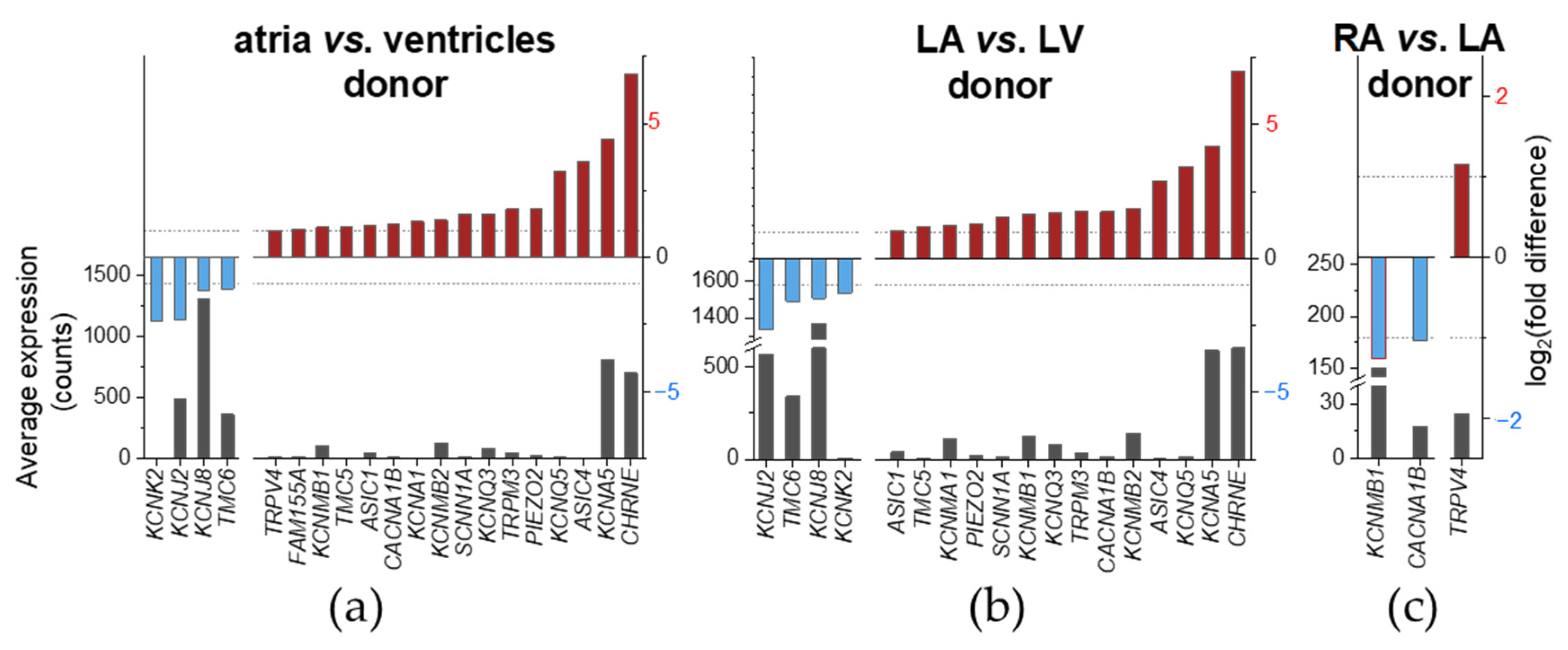
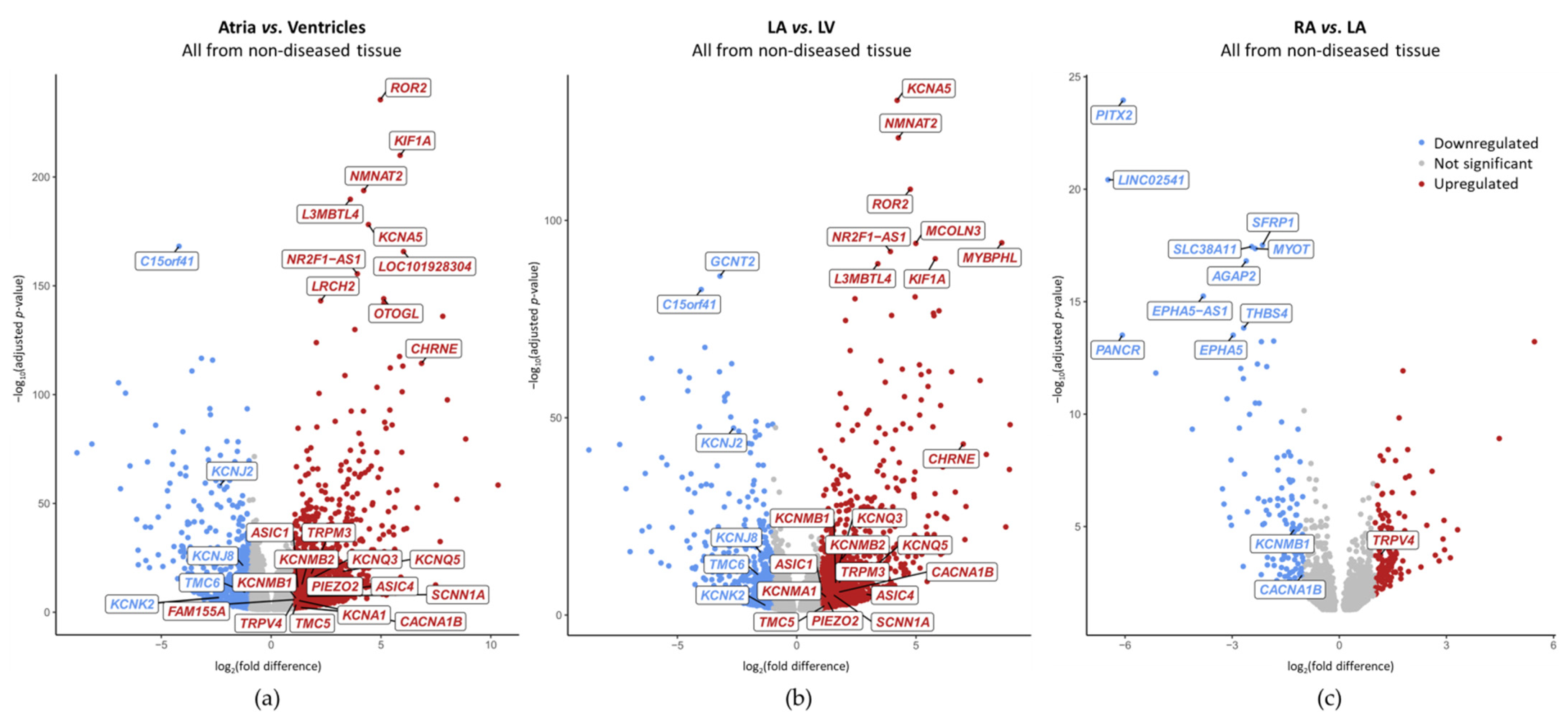
2.3. Chamber-Preferential MSC mRNA Expression
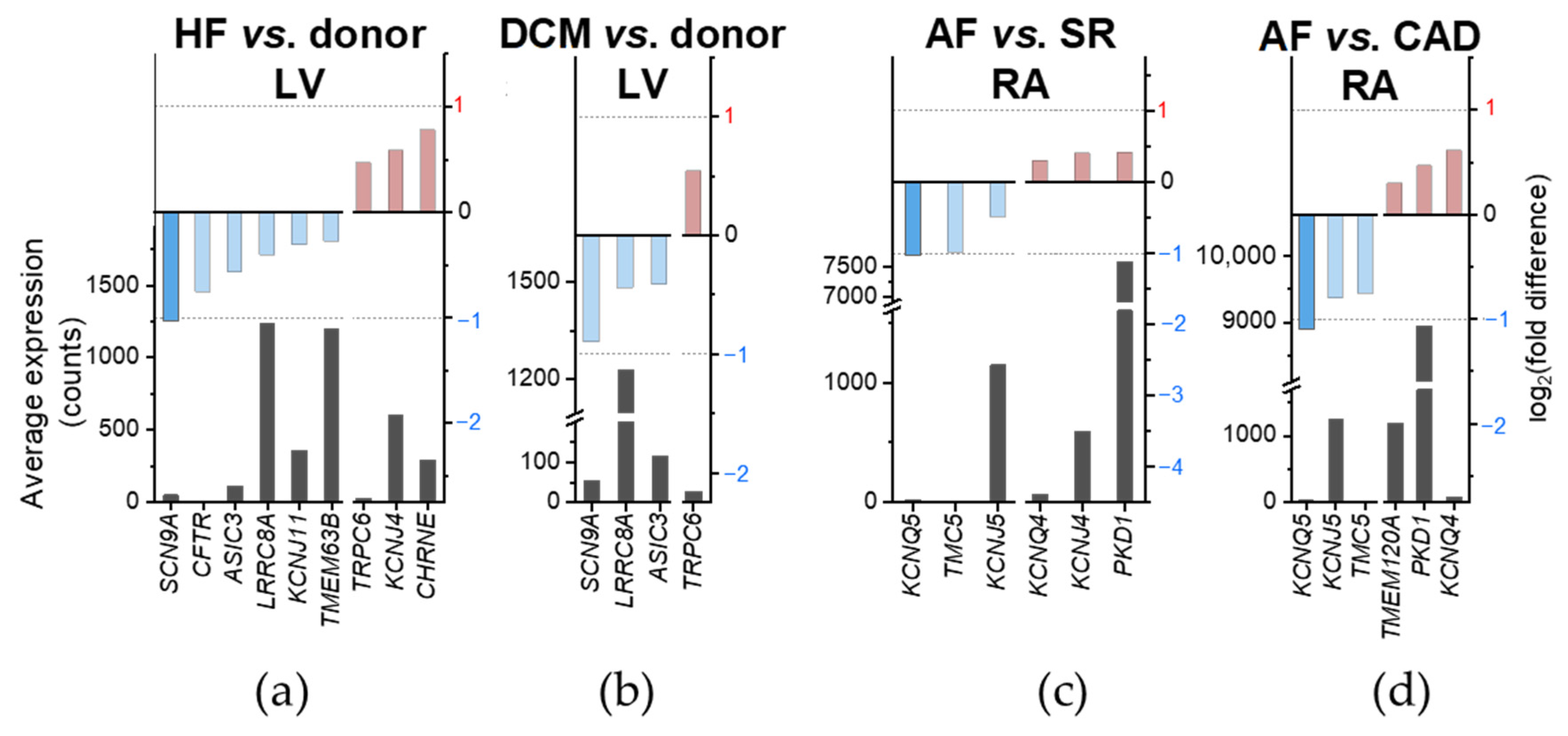
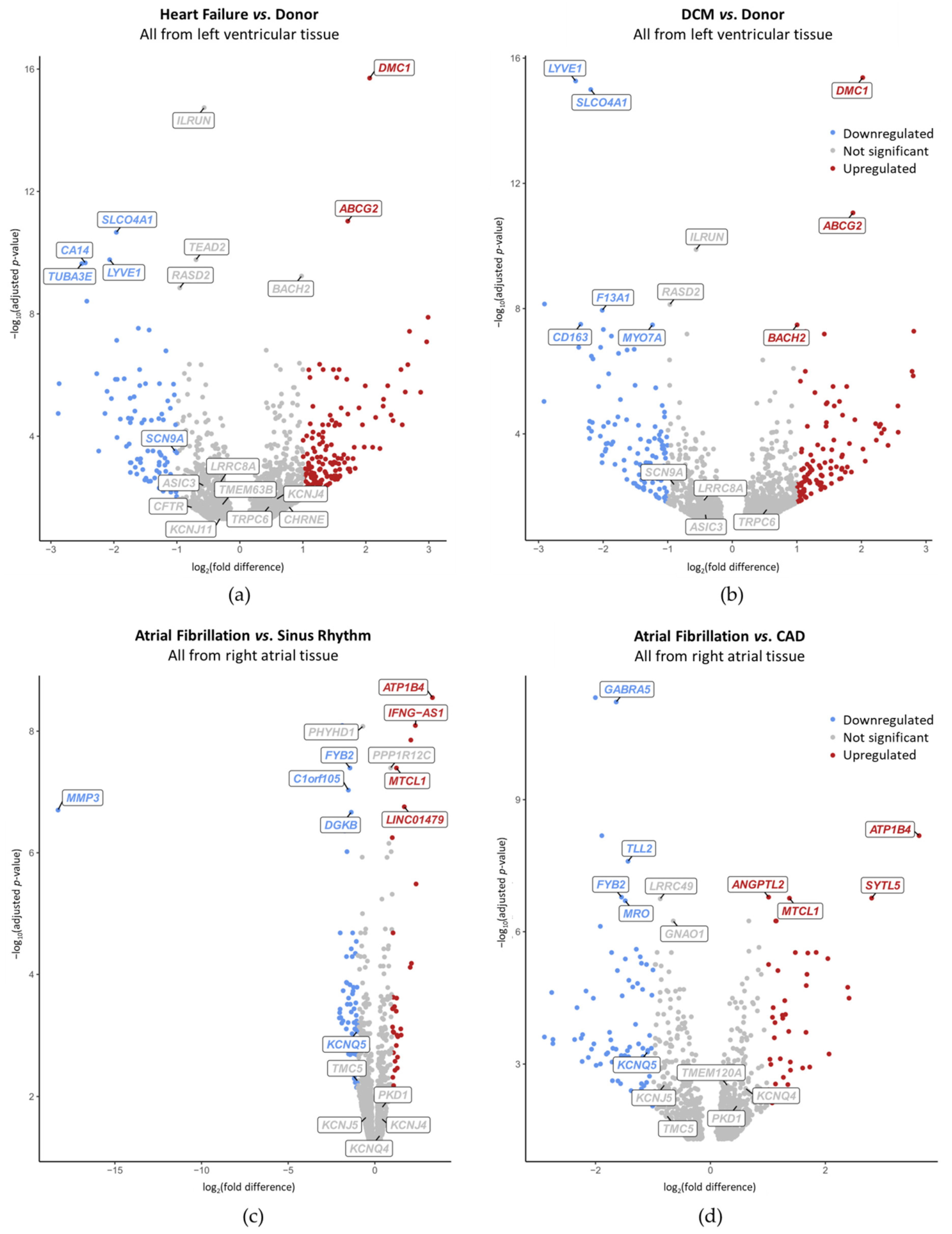
2.4. Disease-Preferential MSC mRNA Expression
3. Discussion
4. Materials and Methods
4.1. Studies Considered for the RNA Sequencing Meta-Analysis
4.2. RNA Sequencing Data Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| AF | Atrial fibrillation |
| Ca2+ | Calcium |
| CAD | Coronary artery disease |
| Cl− | Chloride |
| DCM | Dilated cardiomyopathy |
| HF | Heart failure |
| HVD | Heart valve disease |
| ICM | Ischemic cardiomyopathy |
| K+ | Potassium |
| LA | Left atrium |
| LV | Left ventricle |
| mRNA | Messenger RNA |
| MMC | Mechanically modulated ion channel |
| MSC | Mechano-sensitive ion channel |
| Na+ | Sodium |
| PC | Principal component |
| RA | Right atrium |
| RNA | Ribonucleic acid |
| RV | Right ventricle |
| SAC | Stretch-activated ion channel |
| SACK | K+-selective SAC |
| SACNS | Cation non-selective SAC |
| SEM | Standard error to the (arithmetic) mean |
| SR | Sinus rhythm |
| SRA | Sequence read archive |
| TRP | Transient receptor potential ion channel |
| VAC | Volume-activated ion channel |
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| ↗ | LV | RV | LA | RA | |||
| ↘ | |||||||
| LV | ASIC1,4 CACNA1B CHRNE KCNA5 KCNMA1 KCNMB1,2 KCNQ3,5 PIEZO2 SCNN1A TMC5 TRPM3 | Atrial-preferential expression | |||||
| ASIC1,4 CACNA1B CHRNE FAM155A KCNA1,5 KCNMB1,2 KCNQ3,5 PIEZO2 SCNN1A TMC5 TRPM3 TRPV4 | |||||||
| RV | |||||||
| LA | KCNJ2,8 KCNK2 TMC6 | Ventricular-prefer. expression | TRPV4 | Left/right-prefer. atrial expression | |||
| KCNJ2,8 KCNK2 TMC6 | |||||||
| RA | CACNA1B KCNMB1 | ||||||
References
- Gasparski, A.N.; Beningo, K.A. Mechanoreception at the cell membrane: More than the integrins. Arch. Biochem. Biophys. 2015, 586, 20–26. [Google Scholar] [CrossRef] [PubMed]
- Murthy, S.E.; Dubin, A.E.; Patapoutian, A. Piezos thrive under pressure: Mechanically activated ion channels in health and disease. Nat. Rev. Mol. Cell Biol. 2017, 18, 771–783. [Google Scholar] [CrossRef] [PubMed]
- Gaudet, P.; Livstone, M.S.; Lewis, S.E.; Thomas, P.D. Phylogenetic-based propagation of functional annotations within the Gene Ontology consortium. Brief. Bioinform. 2011, 12, 449–462. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- AgBase; BHF-UCL.; DictyBase; UK-UCL.; Parkinson’s; HGNC.; Roslin Institute; FlyBase; UniProtKB. go-site/goref-0000024.md at master · geneontology/go-site · GitHub. 2019. Available online: https://github.com/geneontology/go-site/blob/master/metadata/gorefs/goref-0000024.md (accessed on 1 October 2021).
- Lyon, R.C.; Zanella, F.; Omens, J.H.; Sheikh, F. Mechanotransduction in cardiac hypertrophy and failure. Circ. Res. 2015, 116, 1462–1476. [Google Scholar] [CrossRef] [Green Version]
- Du, J.; Xie, J.; Zhang, Z.; Tsujikawa, H.; Fusco, D.; Silverman, D.; Liang, B.; Yue, L. TRPM7-mediated Ca2+ signals confer fibrogenesis in human atrial fibrillation. Circ. Res. 2010, 106, 992–1003. [Google Scholar] [CrossRef] [Green Version]
- Guinamard, R.; Chatelier, A.; Demion, M.; Potreau, D.; Patri, S.; Rahmati, M.; Bois, P. Functional characterization of a Ca2+-activated non-selective cation channel in human atrial cardiomyocytes. J. Physiol. 2004, 558, 75–83. [Google Scholar] [CrossRef]
- Kruse, M.; Schulze-Bahr, E.; Corfield, V.; Beckmann, A.; Stallmeyer, B.; Kurtbay, G.; Ohmert, I.; Schulze-Bahr, E.; Brink, P.; Pongs, O. Impaired endocytosis of the ion channel TRPM4 is associated with human progressive familial heart block type I. J. Clin. Investig. 2009, 119, 2737–2744. [Google Scholar] [CrossRef]
- Stallmeyer, B.; Zumhagen, S.; Denjoy, I.; Duthoit, G.; Hébert, J.-L.; Ferrer, X.; Maugenre, S.; Schmitz, W.; Kirchhefer, U.; Schulze-Bahr, E.; et al. Mutational spectrum in the Ca2+-activated cation channel gene TRPM4 in patients with cardiac conductance disturbances. Hum. Mutat. 2012, 33, 109–117. [Google Scholar] [CrossRef]
- Kuwahara, K.; Nakao, K. New molecular mechanisms for cardiovascular disease: Transcriptional pathways and novel therapeutic targets in heart failure. J. Pharmacol. Sci. 2011, 116, 337–342. [Google Scholar] [CrossRef] [Green Version]
- Kuwahara, K.; Wang, Y.; McAnally, J.; Richardson, J.A.; Bassel-Duby, R.; Hill, J.A.; Olson, E.N. TRPC6 fulfills a calcineurin signaling circuit during pathologic cardiac remodeling. J. Clin. Investig. 2006, 116, 3114–3126. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Morley, M.; Brandimarto, J.; Hannenhalli, S.; Hu, Y.; Ashley, E.A.; Tang, W.H.W.; Moravec, C.S.; Margulies, K.B.; Cappola, T.P.; et al. RNA-Seq identifies novel myocardial gene expression signatures of heart failure. Genomics 2015, 105, 83–89. [Google Scholar] [CrossRef]
- Schiano, C.; Costa, V.; Aprile, M.; Grimaldi, V.; Maiello, C.; Esposito, R.; Soricelli, A.; Colantuoni, V.; Donatelli, F.; Ciccodicola, A.; et al. Heart failure: Pilot transcriptomic analysis of cardiac tissue by RNA-sequencing. Cardiol. J. 2017, 24, 539–553. [Google Scholar] [CrossRef] [Green Version]
- Johnson, E.K.; Matkovich, S.J.; Nerbonne, J.M. Regional Differences in mRNA and lncRNA Expression Profiles in Non-Failing Human Atria and Ventricles. Sci. Rep. 2018, 8, 13919. [Google Scholar] [CrossRef] [Green Version]
- Thomas, A.M.; Cabrera, C.P.; Finlay, M.; Lall, K.; Nobles, M.; Schilling, R.J.; Wood, K.; Mein, C.A.; Barnes, M.R.; Munroe, P.B.; et al. Differentially expressed genes for atrial fibrillation identified by rna sequencing from paired human left and right atrial appendages. Physiol. Genom. 2019, 51, 323–332. [Google Scholar] [CrossRef]
- Darkow, E.; Nguyen, T.T.; Stolina, M.; Kari, F.A.; Schmidt, C.; Wiedmann, F.; Baczkó, I.; Kohl, P.; Rajamani, S.; Ravens, U.; et al. Small Conductance Ca2+-Activated K+ (SK) Channel mRNA Expression in Human Atrial and Ventricular Tissue: Comparison Between Donor, Atrial Fibrillation and Heart Failure Tissue. Front. Physiol. 2021, 12, 342. [Google Scholar] [CrossRef]
- Martinac, B. The ion channels to cytoskeleton connection as potential mechanism of mechanosensitivity. Biochim. Biophys. Acta—Biomembr. 2014, 1838, 682–691. [Google Scholar] [CrossRef] [Green Version]
- Peyronnet, R.; Nerbonne, J.M.; Kohl, P. Cardiac Mechano-Gated Ion Channels and Arrhythmias. Circ. Res. 2016, 118, 311–329. [Google Scholar] [CrossRef] [Green Version]
- Ashburner, M.; Ball, C.A.; Blake, J.A.; Botstein, D.; Butler, H.; Cherry, J.M.; Davis, A.P.; Dolinski, K.; Dwight, S.S.; Eppig, J.T.; et al. Gene ontology: Tool for the unification of biology. Nat. Genet. 2000, 25, 25–29. [Google Scholar] [CrossRef] [Green Version]
- Carbon, S.; Douglass, E.; Good, B.M.; Unni, D.R.; Harris, N.L.; Mungall, C.J.; Basu, S.; Chisholm, R.L.; Dodson, R.J.; Hartline, E.; et al. The Gene Ontology resource: Enriching a GOld mine. Nucleic Acids Res. 2021, 49, D325–D334. [Google Scholar] [CrossRef]
- Barth, D.; Fronius, M. Shear force modulates the activity of acid-sensing ion channels at low pH or in the presence of non-proton ligands. Sci. Rep. 2019, 9, 6781. [Google Scholar] [CrossRef] [Green Version]
- Calabrese, B.; Tabarean, I.V.; Juranka, P.; Morris, C.E. Mechanosensitivity of N-type calcium channel currents. Biophys. J. 2002, 83, 2560–2574. [Google Scholar] [CrossRef] [Green Version]
- Langton, P.D. Calcium channel currents recorded from isolated myocytes of rat basilar artery are stretch sensitive. J. Physiol. 1993, 471, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Basson, M.D.; Zeng, B.; Downey, C.; Sirivelu, M.P.; Tepe, J.J. Increased extracellular pressure stimulates tumor proliferation by a mechanosensitive calcium channel and PKC-β. Mol. Oncol. 2015, 9, 513–526. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.K.; Wang, D.; Duan, Y.; Loy, M.M.T.; Chan, H.C.; Huang, P. Mechanosensitive gating of CFTR. Nat. Cell Biol. 2010, 12, 507–512. [Google Scholar] [CrossRef]
- Pan, N.C.; Ma, J.J.; Peng, H.B. Mechanosensitivity of nicotinic receptors. Pflügers Arch.—Eur. J. Physiol. 2012, 464, 193–203. [Google Scholar] [CrossRef] [Green Version]
- Duan, D.; Winter, C.; Hume, J.R.; Horowitz, B. Molecular identification of a volume-regulated chloride channel. Nature 1997, 390, 417–421. [Google Scholar] [CrossRef]
- Bao, L.; Sachs, F.; Dahl, G. Connexins are mechanosensitive. Am. J. Physiol.—Cell Physiol. 2004, 287, C1389-95. [Google Scholar] [CrossRef] [Green Version]
- Kloda, A.; Lua, L.; Hall, R.; Adams, D.J.; Martinac, B. Liposome reconstitution and modulation of recombinant N-methyl-D-aspartate receptor channels by membrane stretch. Proc. Natl. Acad. Sci. USA 2007, 104, 1540–1545. [Google Scholar] [CrossRef] [Green Version]
- Singh, P.; Doshi, S.; Spaethling, J.M.; Hockenberry, A.J.; Patel, T.P.; Geddes-Klein, D.M.; Lynch, D.R.; Meaney, D.F. N-methyl-D-aspartate receptor mechanosensitivity is governed by C terminus of NR2B subunit. J. Biol. Chem. 2012, 287, 4348–4359. [Google Scholar] [CrossRef] [Green Version]
- Hao, J.; Padilla, F.; Dandonneau, M.; Lavebratt, C.; Lesage, F.; Noël, J.; Delmas, P. Kv1.1 channels act as mechanical brake in the senses of touch and pain. Neuron 2013, 77, 899–914. [Google Scholar] [CrossRef] [Green Version]
- Milton, A.O.; Wang, T.; Li, W.; Guo, J.; Zhang, S.; Shipston, M. Mechanical stretch increases Kv1.5 current through an interaction between the S1-S2 linker and N-terminus of the channel. J. Biol. Chem. 2020, 295, 4723–4732. [Google Scholar] [CrossRef] [PubMed]
- He, Y.; Xiao, J.; Yang, Y.; Zhou, Q.; Zhang, Z.; Pan, Q.; Liu, Y.; Chen, Y. Stretch-induced alterations of human Kir2.1 channel currents. Biochem. Biophys. Res. Commun. 2006, 351, 462–467. [Google Scholar] [CrossRef] [PubMed]
- Dyachenko, V.; Husse, B.; Rueckschloss, U.; Isenberg, G. Mechanical deformation of ventricular myocytes modulates both TRPC6 and Kir2.3 channels. Cell Calcium 2009, 45, 38–54. [Google Scholar] [CrossRef] [PubMed]
- Ji, S.; John, S.A.; Lu, Y.; Weiss, J.N. Mechanosensitivity of the Cardiac Muscarinic Potassium Channel. J. Biol. Chem. 1998, 273, 1324–1328. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Wagoner, D.R. Mechanosensitive gating of atrial ATP-sensitive potassium channels. Circ. Res. 1993, 72, 973–983. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Patel, A.J.; Honoré, E.; Maingret, F.; Lesage, F.; Fink, M.; Duprat, F.; Lazdunski, M. A mammalian two pore domain mechano-gated S-like K+ channel. EMBO J. 1998, 17, 4283–4290. [Google Scholar] [CrossRef]
- Maingret, F.; Fosset, M.; Lesage, F.; Lazdunski, M.; Honoré, E. TRAAK is a mammalian neuronal mechano-gated K+ channel. J. Biol. Chem. 1999, 274, 1381–1387. [Google Scholar] [CrossRef] [Green Version]
- Lesage, F.; Terrenoire, C.; Romey, G.; Lazdunski, M. Human TREK2, a 2P Domain Mechano-sensitive K+ Channel with Multiple Regulations by Polyunsaturated Fatty Acids, Lysophospholipids, and Gs, Gi, and Gq Protein-coupled Receptors. J. Biol. Chem. 2000, 275, 28398–28405. [Google Scholar] [CrossRef] [Green Version]
- Naruse, K.; Tang, Q.Y.; Sokabe, M. Stress-Axis Regulated Exon (STREX) in the C terminus of BKCa channels is responsible for the stretch sensitivity. Biochem. Biophys. Res. Commun. 2009, 385, 634–639. [Google Scholar] [CrossRef]
- Kubota, T.; Horie, M.; Takano, M.; Yoshida, H.; Otani, H.; Sasayama, S. Role of KCNQ1 in the cell swelling-induced enhancement of the slowly activating delayed rectifier K+ current. Jpn. J. Physiol. 2002, 52, 31–39. [Google Scholar] [CrossRef] [Green Version]
- Schütze, S.; Orozco, I.J.; Jentsch, T.J. KCNQ potassium channels modulate sensitivity of skin Down-hair (D-hair) mechanoreceptors. J. Biol. Chem. 2016, 291, 5566–5575. [Google Scholar] [CrossRef] [Green Version]
- Grunnet, M.; Jespersen, T.; MacAulay, N.; Jørgensen, N.K.; Schmitt, N.; Pongs, O.; Olesen, S.P.; Klaerke, D.A. KCNQ1 channels sense small changes in cell volume. J. Physiol. 2003, 549, 419–427. [Google Scholar] [CrossRef]
- Jensen, H.S.; Callø, K.; Jespersen, T.; Jensen, B.S.; Olesen, S.P. The KCNQ5 potassium channel from mouse: A broadly expressed M-current like potassium channel modulated by zinc, pH, and volume changes. Mol. Brain Res. 2005, 139, 52–62. [Google Scholar] [CrossRef]
- Voss, F.K.; Ullrich, F.; Münch, J.; Lazarow, K.; Lutte, D.; Mah, N.; Andrade-Navarro, M.A.; Von Kries, J.P.; Stauber, T.; Jentsch, T.J. Identification of LRRC8 heteromers as an essential component of the volume-regulated anion channel VRAC. Science 2014, 344, 634–638. [Google Scholar] [CrossRef] [Green Version]
- Syeda, R.; Qiu, Z.; Dubin, A.E.; Murthy, S.E.; Florendo, M.N.; Mason, D.E.; Mathur, J.; Cahalan, S.M.; Peters, E.C.; Montal, M.; et al. LRRC8 Proteins Form Volume-Regulated Anion Channels that Sense Ionic Strength. Cell 2016, 164, 499–511. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Coste, B.; Mathur, J.; Schmidt, M.; Earley, T.J.; Ranade, S.; Petrus, M.J.; Dubin, A.E.; Patapoutian, A. Piezo1 and Piezo2 are essential components of distinct mechanically activated cation channels. Science 2010, 330, 55–60. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sharif-Naeini, R.; Folgering, J.H.A.; Bichet, D.; Duprat, F.; Lauritzen, I.; Arhatte, M.; Jodar, M.; Dedman, A.; Chatelain, F.C.; Schulte, U.; et al. Polycystin-1 and -2 Dosage Regulates Pressure Sensing. Cell 2009, 139, 587–596. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aboualaiwi, W.A.; Takahashi, M.; Mell, B.R.; Jones, T.J.; Ratnam, S.; Kolb, R.J.; Nauli, S.M. Ciliary polycystin-2 is a mechanosensitive calcium channel involved in nitric oxide signaling cascades. Circ. Res. 2009, 104, 860–869. [Google Scholar] [CrossRef]
- Tabarean, I.V.; Juranka, P.; Morris, C.E. Membrane stretch affects gating modes of a skeletal muscle sodium channel. Biophys. J. 1999, 77, 758–774. [Google Scholar] [CrossRef] [Green Version]
- Beyder, A.; Rae, J.L.; Bernard, C.; Strege, P.R.; Sachs, F.; Farrugia, G. Mechanosensitivity of Nav1.5, a voltage-sensitive sodium channel. J. Physiol. 2010, 588, 4969–4985. [Google Scholar] [CrossRef]
- Wang, J.A.; Lin, W.; Morris, T.; Banderali, U.; Juranka, P.F.; Morris, C.E. Membrane trauma and Na+ leak from Nav1.6 channels. Am. J. Physiol.—Cell Physiol. 2009, 297, C823–C834. [Google Scholar] [CrossRef] [Green Version]
- Körner, J.; Meents, J.; Machtens, J.P.; Lampert, A. β1 subunit stabilises sodium channel Nav1.7 against mechanical stress. J. Physiol. 2018, 596, 2433–2445. [Google Scholar] [CrossRef] [Green Version]
- Kizer, N.; Guo, X.L.; Hruska, K. Reconstitution of stretch-activated cation channels by expression of the α-subunit of the epithelial sodium channel cloned from osteoblasts. Proc. Natl. Acad. Sci. USA 1997, 94, 1013–1018. [Google Scholar] [CrossRef] [Green Version]
- Kawashima, Y.; Géléoc, G.S.G.; Kurima, K.; Labay, V.; Lelli, A.; Asai, Y.; Makishima, T.; Wu, D.K.; Della Santina, C.C.; Holt, J.R.; et al. Mechanotransduction in mouse inner ear hair cells requires transmembrane channel-like genes. J. Clin. Investig. 2011, 121, 4796–4809. [Google Scholar] [CrossRef]
- Beaulieu-Laroche, L.; Christin, M.; Donoghue, A.M.; Agosti, F.; Yousefpour, N.; Petitjean, H.; Davidova, A.; Stanton, C.; Khan, U.; Dietz, C.; et al. TACAN is an essential component of the mechanosensitive ion channel responsible for pain sensing. bioRxiv 2018. [Google Scholar] [CrossRef]
- Hong, G.S.; Lee, B.; Wee, J.; Chun, H.; Kim, H.; Jung, J.; Cha, J.Y.; Riew, T.R.; Kim, G.H.; Kim, I.B.; et al. Tentonin 3/TMEM150c Confers Distinct Mechanosensitive Currents in Dorsal-Root Ganglion Neurons with Proprioceptive Function. Neuron 2016, 91, 107–118. [Google Scholar] [CrossRef]
- Murthy, S.E.; Dubin, A.E.; Whitwam, T.; Jojoa-Cruz, S.; Cahalan, S.M.; Mousavi, S.A.R.; Ward, A.B.; Patapoutian, A. OSCA/TMEM63 are an evolutionarily conserved family of mechanically activated ion channels. Elife 2018, 7, e41844. [Google Scholar] [CrossRef]
- Moparthi, L.; Zygmunt, P.M. Human TRPA1 is an inherently mechanosensitive bilayer-gated ion channel. Cell Calcium 2020, 91, 102255. [Google Scholar] [CrossRef]
- Maroto, R.; Raso, A.; Wood, T.G.; Kurosky, A.; Martinac, B.; Hamill, O.P. TRPC1 forms the stretch-activated cation channel in vertebrate cells. Nat. Cell Biol. 2005, 7, 179–185. [Google Scholar] [CrossRef]
- Quick, K.; Zhao, J.; Eijkelkamp, N.; Linley, J.E.; Rugiero, F.; Cox, J.J.; Raouf, R.; Gringhuis, M.; Sexton, J.E.; Abramowitz, J.; et al. TRPC3 and TRPC6 are essential for normal mechanotransduction in subsets of sensory neurons and cochlear hair cells. Open Biol. 2012, 2, 120068. [Google Scholar] [CrossRef] [Green Version]
- Gomis, A.; Soriano, S.; Belmonte, C.; Viana, F. Hypoosmotic- and pressure-induced membrane stretch activate TRPC5 channels. J. Physiol. 2008, 586, 5633–5649. [Google Scholar] [CrossRef] [Green Version]
- Spassova, M.A.; Hewavitharana, T.; Xu, W.; Soboloff, J.; Gill, D.L. A common mechanism underlies stretch activation and activation of TRPC6 channels. Proc. Natl. Acad. Sci. USA 2006, 103, 16586–16591. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grimm, C.; Kraft, R.; Sauerbruch, S.; Schultz, G.; Harteneck, C. Molecular and functional characterization of the melastatin-related cation channel TRPM3. J. Biol. Chem. 2003, 278, 21493–21501. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morita, H.; Honda, A.; Inoue, R.; Ito, Y.; Abe, K.; Nelson, M.T.; Brayden, J.E. Membrane Stretch-Induced Activation of a TRPM4-Like Nonselective Cation Channel in Cerebral Artery Myocytes. J. Pharmacol. Sci. 2007, 103, 417–426. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Numata, T.; Shimizu, T.; Okada, Y. Direct Mechano-Stress Sensitivity of TRPM7 Channel. Cell. Physiol. Biochem. 2007, 19, 1–8. [Google Scholar] [CrossRef]
- Feng, N.H.; Lee, H.H.; Shiang, J.C.; Ma, M.C. Transient receptor potential vanilloid type 1 channels act as mechanoreceptors and cause substance P release and sensory activation in rat kidneys. Am. J. Physiol.—Ren. Physiol. 2008, 294, F316–F325. [Google Scholar] [CrossRef] [Green Version]
- Muraki, K.; Iwata, Y.; Katanosaka, Y.; Ito, T.; Ohya, S.; Shigekawa, M.; Imaizumi, Y. TRPV2 Is a Component of Osmotically Sensitive Cation Channels in Murine Aortic Myocytes. Circ. Res. 2003, 93, 829–838. [Google Scholar] [CrossRef] [Green Version]
- Köttgen, M.; Buchholz, B.; Garcia-Gonzalez, M.A.; Kotsis, F.; Fu, X.; Doerken, M.; Boehlke, C.; Steffl, D.; Tauber, R.; Wegierski, T.; et al. TRPP2 and TRPV4 form a polymodal sensory channel complex. J. Cell Biol. 2008, 182, 437–447. [Google Scholar] [CrossRef]
- Wang, Z.; Fermini, B.; Nattel, S. Sustained depolarization-induced outward current in human atrial myocytes: Evidence for a novel delayed rectifier K+ current similar to Kv1.5 cloned channel currents. Circ. Res. 1993, 73, 1061–1076. [Google Scholar] [CrossRef] [Green Version]
- Peyronnet, R.; Ravens, U. Atria-selective antiarrhythmic drugs in need of alliance partners. Pharmacol. Res. 2019, 145, 104262. [Google Scholar] [CrossRef]
- Pabel, S.; Ahmad, S.; Tirilomis, P.; Stehle, T.; Mustroph, J.; Knierim, M.; Dybkova, N.; Bengel, P.; Holzamer, A.; Hilker, M.; et al. Inhibition of NaV1.8 prevents atrial arrhythmogenesis in human and mice. Basic Res. Cardiol. 2020, 115, 20. [Google Scholar] [CrossRef]
- Ellinghaus, P.; Scheubel, R.J.; Dobrev, D.; Ravens, U.; Holtz, J.; Huetter, J.; Nielsch, U.; Morawietz, H. Comparing the global mRNA expression profile of human atrial and ventricular myocardium with high-density oligonucleotide arrays. J. Thorac. Cardiovasc. Surg. 2005, 129, 1383–1390. [Google Scholar] [CrossRef] [Green Version]
- Gaborit, N.; Le Bouter, S.; Szuts, V.; Varro, A.; Escande, D.; Nattel, S.; Demolombe, S. Regional and tissue specific transcript signatures of ion channel genes in the non-diseased human heart. J. Physiol. 2007, 582, 675–693. [Google Scholar] [CrossRef]
- Barth, A.S.; Merk, S.; Arnoldi, E.; Zwermann, L.; Kloos, P.; Gebauer, M.; Steinmeyer, K.; Bleich, M.; Kääb, S.; Pfeufer, A.; et al. Functional profiling of human atrial and ventricular gene expression. Pflugers Arch. Eur. J. Physiol. 2005, 450, 201–208. [Google Scholar] [CrossRef]
- Wong, K.R.; Trezise, A.E.O.; Crozatier, B.; Vandenberg, J.I. Loss of the normal epicardial to endocardial gradient of cftr mRNA expression in the hypertrophied rabbit left ventricle. Biochem. Biophys. Res. Commun. 2000, 278, 144–149. [Google Scholar] [CrossRef]
- Solbach, T.F.; Paulus, B.; Weyand, M.; Eschenhagen, T.; Zolk, O.; Fromm, M.F. ATP-binding cassette transporters in human heart failure. Naunyn. Schmiedebergs. Arch. Pharmacol. 2008, 377, 231–243. [Google Scholar] [CrossRef]
- Fedorov, V.V.; Glukhov, A.V.; Ambrosi, C.M.; Kostecki, G.; Chang, R.; Janks, D.; Schuessler, R.B.; Moazami, N.; Nichols, C.G.; Efimov, I.R. Effects of KATP channel openers diazoxide and pinacidil in coronary-perfused atria and ventricles from failing and non-failing human hearts. J. Mol. Cell. Cardiol. 2011, 51, 215–225. [Google Scholar] [CrossRef] [Green Version]
- Roselli, C.; Chaffin, M.D.; Weng, L.C.; Aeschbacher, S.; Ahlberg, G.; Albert, C.M.; Almgren, P.; Alonso, A.; Anderson, C.D.; Aragam, K.G.; et al. Multi-ethnic genome-wide association study for atrial fibrillation. Nat. Genet. 2018, 50, 1225–1233. [Google Scholar] [CrossRef] [Green Version]
- Kääb, S.; Barth, A.S.; Margerie, D.; Dugas, M.; Gebauer, M.; Zwermann, L.; Merk, S.; Pfeufer, A.; Steinmeyer, K.; Bleich, M.; et al. Global gene expression in human myocardium—Oligonucleotide microarray analysis of regional diversity and transcriptional regulation in heart failure. J. Mol. Med. 2004, 82, 308–316. [Google Scholar] [CrossRef]
- Peyronnet, R.; Martins, J.R.; Duprat, F.; Demolombe, S.; Arhatte, M.; Jodar, M.; Tauc, M.; Duranton, C.; Paulais, M.; Teulon, J.; et al. Piezo1-dependent stretch-activated channels are inhibited by Polycystin-2 in renal tubular epithelial cells. EMBO Rep. 2013, 14, 1143–1148. [Google Scholar] [CrossRef] [Green Version]
- Jakob, D.; Klesen, A.; Allegrini, B.; Darkow, E.; Aria, D.; Emig, R.; Chica, A.S.; Rog-Zielinska, E.A.; Guth, T.; Beyersdorf, F.; et al. Piezo1 and BKCa channels in human atrial fibroblasts: Interplay and remodelling in atrial fibrillation. J. Mol. Cell. Cardiol. 2021, 158, 49–62. [Google Scholar] [CrossRef]
- Altamirano, F.; Schiattarella, G.G.; French, K.M.; Kim, S.Y.; Engelberger, F.; Kyrychenko, S.; Villalobos, E.; Tong, D.; Schneider, J.W.; Ramirez-Sarmiento, C.A.; et al. Polycystin-1 Assembles With Kv Channels to Govern Cardiomyocyte Repolarization and Contractility. Circulation 2019, 140, 921–936. [Google Scholar] [CrossRef] [PubMed]
- Mukherjee, R.; Akar, J.G.; Wharton, J.M.; Adams, D.K.; McClure, C.D.; Stroud, R.E.; Rice, A.D.; Desantis, S.M.; Spinale, F.G.; Gold, M.R. Plasma profiles of matrix metalloproteinases and tissue inhibitors of the metalloproteinases predict recurrence of atrial fibrillation following cardioversion. J. Cardiovasc. Transl. Res. 2013, 6, 528–535. [Google Scholar] [CrossRef] [PubMed]
- Aloui, S.; Zidi, W.; Fourty, N.; Sanhaji, H.; Mourali, M.S.; Ouali, S.; Feki, M.; Allal-Elasmi, M. Circulating Stromelysin-1 (MMP-3): A potential predictor for atrial fibrillation in patients with mitral stenosis. Arch. Cardiovasc. Dis. Suppl. 2020, 12, 93. [Google Scholar] [CrossRef]
- De With, R.R.; Artola Arita, V.; Nguyen, B.O.; Linz, D.; Ten Cate, H.; Spronk, H.; Schotten, U.; Jan Van Zonneveld, A.; Erküner, Ö.; Bayón, M.A.; et al. Different circulating biomarkers in women and men with paroxysmal atrial fibrillation: Results from the AF-RISK and RACE V studies. Europace 2022, 24, 193–201. [Google Scholar] [CrossRef] [PubMed]
- Boycott, H.E.; Barbier, C.S.M.; Eichel, C.A.; Costa, K.D.; Martins, R.P.; Louault, F.; Dilanian, G.; Coulombe, A.; Hatem, S.N.; Balse, E. Shear stress triggers insertion of voltage-gated potassium channels from intracellular compartments in atrial myocytes. Proc. Natl. Acad. Sci. USA 2013, 110, E3955–E3964. [Google Scholar] [CrossRef] [Green Version]
- Patel, K.K.; Venkatesan, C.; Abdelhalim, H.; Zeeshan, S.; Arima, Y.; Linna-Kuosmanen, S.; Ahmed, Z. Genomic approaches to identify and investigate genes associated with atrial fibrillation and heart failure susceptibility. Hum. Genomics 2023, 17, 47. [Google Scholar] [CrossRef]
- Leinonen, R.; Sugawara, H.; Shumway, M. The sequence read archive. Nucleic Acids Res. 2011, 39, D19. [Google Scholar] [CrossRef] [Green Version]
- Afgan, E.; Baker, D.; Batut, B.; Van Den Beek, M.; Bouvier, D.; Ech, M.; Chilton, J.; Clements, D.; Coraor, N.; Grüning, B.; et al. The Galaxy platform for accessible, reproducible and collaborative biomedical analyses: 2018 update. Nucleic Acids Res. 2018, 46, W537–W544. [Google Scholar] [CrossRef] [Green Version]
- Batut, B.; Hiltemann, S.; Bagnacani, A.; Baker, D.; Bhardwaj, V.; Blank, C.; Bretaudeau, A.; Brillet-Guéguen, L.; Čech, M.; Chilton, J.; et al. Community-Driven Data Analysis Training for Biology. Cell Syst. 2018, 6, 752–758.e1. [Google Scholar] [CrossRef] [Green Version]
- Batut, B.; Freeberg, M.A.; Heydarian, M.; Erxleben, A.; Videm, P.; Blank, C.; Doyle, M.; Soranzo, N.; van Heusden, P. Reference-Based RNA-Seq Data Analysis (Galaxy Training Materials). 2020. Available online: https://training.galaxyproject.org/training-material/topics/transcriptomics/tutorials/ref-based/tutorial.html#citing-this-tutorial (accessed on 5 November 2020).
- Grüning, B. GitHub—Bgruening/Galaxytools: Galaxy Tool Wrappers. Available online: https://github.com/bgruening/galaxytools (accessed on 30 September 2021).
- Andrews, S. FastQC A Quality Control Tool for High Throughput Sequence Data. 2010. Available online: http://www.bioinformatics.babraham.ac.uk/projects/fastqc/ (accessed on 7 October 2021).
- Ewels, P.; Magnusson, M.; Lundin, S.; Käller, M. MultiQC: Summarize analysis results for multiple tools and samples in a single report. Bioinformatics 2016, 32, 3047–3048. [Google Scholar] [CrossRef] [Green Version]
- Marcel, M. Cutadapt removes adapter sequences from high-throughput sequencing reads. EMBnet. J. 2011, 17, 10–12. [Google Scholar] [CrossRef]
- Dobin, A.; Davis, C.A.; Schlesinger, F.; Drenkow, J.; Zaleski, C.; Jha, S.; Batut, P.; Chaisson, M.; Gingeras, T.R. STAR: Ultrafast universal RNA-seq aligner. Bioinformatics 2012, 29, 15–21. [Google Scholar] [CrossRef]
- Robinson, J.T.; Thorvaldsdóttir, H.; Winckler, W.; Guttman, M.; Lander, E.S.; Getz, G.; Mesirov, J.P. Integrative genomics viewer. Nat. Biotechnol. 2011, 29, 24–26. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.; Wang, S.; Li, W. RSeQC: Quality control of RNA-seq experiments. Bioinformatics 2012, 28, 2184–2185. [Google Scholar] [CrossRef] [Green Version]
- Liao, Y.; Smyth, G.K.; Shi, W. featureCounts: An efficient general purpose program for assigning sequence reads to genomic features. Bioinformatics 2013, 30, 923–930. [Google Scholar] [CrossRef] [Green Version]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef] [Green Version]
- Core R Team, X. R: A Language and Environment for Statistical Computing. 2021. Available online: https://www.r-project.org/ (accessed on 14 November 2021).
- Wickham, H. ggplot2: Elegant Graphics for Data Analysis; Springer: New York, NY, USA, 2009. [Google Scholar] [CrossRef]
- Ritchie, M.E.; Phipson, B.; Wu, D.; Hu, Y.; Law, C.W.; Shi, W.; Smyth, G.K. Limma powers differential expression analyses for RNA-sequencing and microarray studies. Nucleic Acids Res. 2015, 43, e47. [Google Scholar] [CrossRef] [Green Version]
- Stephens, M. False discovery rates: A new deal. Biostatistics 2017, 18, 275–294. [Google Scholar] [CrossRef] [Green Version]






| Gene | Protein | Category | Reference |
|---|---|---|---|
| ASIC1,2,3,4,5 | Acid-sensing ion channel 1,2,3,4,5 | MMC (Na+) | [21] |
| CACNA1B,C,I | Voltage-dependent N,L,T-type Ca2+ channel subunit α-1B,C,I or Cav2.2, 1.2, 3.3 | MMC (Ca2+) | [22,23,24] |
| CFTR | Cystic fibrosis transmembrane conductance receptor | MMC (Cl−) | [25] |
| CHRNA1,B1,D,E | Acetylcholine receptor nicotinic subunit α1,β1,δ,ε | MMC | [26] |
| CLCN3 | H+/Cl- exchange transporter 3 or CLC-3 | MMC (Cl−) | [27] |
| FAM155A,B | Transmembrane protein FAM155A,B | candidate | [3] |
| GJA3 | Gap junction α-3 protein or Connexin 46 | MMC | [28] |
| GRIN1, 2(A,B) | Ionotropic glutamate receptor NMDA type subunit 1, 2 | [29,30] | |
| KCNA1,5 | K+ voltage-gated channel subfamily A member 1,5 or Kv1.1,1.5 | SACK | [31,32] |
| KCNJ2,4 | Inward rectifier K+ channel 2,4, or Kir2.1,2.3 | [33,34] | |
| KCNJ5 | G protein-activated inward rectifier K+ channel 4 or Kir3.4 or GIRK4 | [35] | |
| KCNJ8,11 | ATP-sensitive inward rectifier K+ channel 8,11 or Kir6.1,6.2 | [36] | |
| KCNK2,4,10 | K+ channel subfamily K member 2,4,10 or TREK-1, TRAAK, TREK-2 | [37,38,39] | |
| KCNMA1 | Ca2+-activated K+ channel subunit α-1 or KCa1.1 or BKCa α | SACK, MMC (K+) | [40] |
| KCNMB1,2,3,4 | Ca2+-activated K+ channel subunit β-1,2,3,4 or BKβ1,2,3,4 | ||
| KCNQ1,2,3,4,5 | K+ voltage-gated channel subfamily KQT member 1,2,3,4,5 or Kv7.1,7.2,7.3,7.4,7.5 | VAC | [41,42,43,44] |
| LRRC8A | Leucine rich repeat containing 8 VRAC subunit A or SWELL1 | [45,46] | |
| PIEZO1,2 | Piezo-type mechano-sensitive ion channel component 1,2 | SACNS | [47] |
| PKD1,2 | Polycystin-1,2 or Transient receptor potential cation channel subfamily P member 1,2 or TRPP1,2 | MMC (Ca2+) | [48,49] |
| SCN(4,5,8,9)A | Na+ channel protein type 4,5,8,9 subunit α or Nav1.4,1.5,1.6,1.7 | SACNS, MMC (Na+) | [50,51,52,53] |
| SCNN1A | Na+ channel epithelial 1 subunit α or ENaCα | SACNS | [54] |
| TMC1,2,3,4,5,6,7,8 | Transmembrane channel like protein 1,2,3,4,5,6,7,8 | MMC (Cl−), candidate | [3,55] |
| TMEM120A | TACAN | SACNS | [56] |
| TMEM150C | Transmembrane protein 150C or Tentonin 3 | MMC | [57] |
| TMEM63A,B | CSC-1 like protein 1,2 | SACNS, candidate | [4,58] |
| TRPA1 | Transient receptor potential cation channel subfamily A member 1 | SACNS | [59] |
| TRPC1,3,5,6 | Short transient receptor potential channel 1,3,5,6 | [60,61,62,63] | |
| TRPM3,4,7 | Transient receptor potential cation channel subfamily M member 3,4,7 | [64,65,66] | |
| TRPV1,2,4 | Transient receptor potential cation channel subfamily V member 1,2,4 | [67,68,69] |
| Gene | Atria vs. Ventricles | LA vs. LV | RA vs. LA |
|---|---|---|---|
| ASIC1 | ↗ | ↗ | − |
| ASIC4 | ↑ | ↑ | − |
| CACNA1B | ↗ | ↑ | ↘ |
| CHRNE | ↑ | ↑ | − |
| FAM155A | ↗ | − | − |
| KCNA1 | ↗ | − | − |
| KCNA5 | ↑ | ↑ | − |
| KCNJ2 | ↓ | ↓ | − |
| KCNJ8 | ↘ | ↘ | − |
| KCNK2 | ↓ | ↘ | − |
| KCNMA1 | − | ↗ | − |
| KCNMB1 | ↗ | ↗ | ↘ |
| KCNMB2 | ↗ | ↑ | − |
| KCNQ3 | ↗ | ↑ | − |
| KCNQ5 | ↑ | ↑ | − |
| PIEZO2 | ↑ | ↗ | − |
| SCNN1A | ↗ | ↗ | − |
| TMC5 | ↗ | ↗ | − |
| TMC6 | ↘ | ↘ | − |
| TRPM3 | ↑ | ↑ | − |
| TRPV4 | ↗ | − | ↗ |
| Gene | HF vs. Donor | DCM vs. Donor | AF vs. SR | AF vs. CAD |
|---|---|---|---|---|
| ASIC3 | ↘ | ↘ | − | − |
| CFTR | ↘ | − | − | − |
| CHRNE | ↗ | − | − | − |
| KCNJ4 | ↗ | − | ↗ | − |
| KCNJ5 | − | − | ↘ | ↘ |
| KCNJ11 | ↘ | − | − | − |
| KCNQ4 | − | − | ↗ | ↗ |
| KCNQ5 | − | − | ↘ | ↘ |
| LRRC8A | ↘ | ↘ | − | − |
| PKD1 | − | − | ↗ | ↗ |
| SCN9A | ↘ | ↘ | − | − |
| TMC5 | − | − | ↘ | ↘ |
| TMEM120A | − | − | − | ↗ |
| TMEM63B | ↘ | − | − | − |
| TRPC6 | ↗ | ↗ | − | − |
| Reference | SRA Accession Number | Platform | Health Status | Tissue Provenance | Structural Heart Disease | Sample Size (n/N) | |
|---|---|---|---|---|---|---|---|
| [12] | PRJNA246308 | Illumina HiSeq 2000 | Non-diseased | LV | No | 2/2 | |
| HF | DCM ICM | LV | Yes | 2/2 | |||
| 1/1 | |||||||
| [13] | PRJNA291619 | Illumina HiSeq 2000 | Non-diseased | LV | No | 4/4 | |
| HF | DCM | LV | Yes | 2/2 | |||
| [14] | PRJNA445706 | Illumina HiSeq 2500 | Non-diseased | LV RV LA RA | No | 32/8 | |
| [15] | PRJNA526687 | Illumina NextSeq 500 | SR | CAD | LA, RA | Yes | 8/4 |
| HVD | RA | 1/1 | |||||
| AF | LA RA | Yes | 10/5 | ||||
| [16] | PRJEB42485 | Illumina HiSeq 4000 | Non-diseased | LV | No | 21/8 | |
| LA | |||||||
| RA | |||||||
| HF | DCM | LV | Yes | 5/5 | |||
| ICM | 4/4 | ||||||
| SR | CAD | RA | Yes | 6/6 | |||
| HVD | 4/4 | ||||||
| AF | RA | Yes | 6/6 | ||||
| Tissue Provenance | LV | RV | LA | RA | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Health Status | Non- Diseased | DCM | ICM | Non- Diseased | Non- Diseased | CAD | AF | Non- Diseased | CAD | HVD | AF |
| Sample size | 22 | 9 | 5 | 8 | 14 | 4 | 5 | 15 | 10 | 5 | 11 |
| Sex (♂/♀) | 13/9 | 7/2 | 4/1 | 6/2 | 9/5 | 4/0 | 5/0 | 10/5 | 10/0 | 5/0 | 11/0 |
| Age (mean ± SEM) | 51.2 ± 2.6 | 52.6 ± 3.8 | 54.2 ± 4.1 | 47.4 ± 4.4 | 53.2 ± 3.2 | 62.5 ± 4.0 | 73.4 ± 2.4 | 54.3 ± 3.0 | 64.2 ± 3.1 | 53.4 ± 7.2 | 70.2 ± 1.6 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Darkow, E.; Yusuf, D.; Rajamani, S.; Backofen, R.; Kohl, P.; Ravens, U.; Peyronnet, R. Meta-Analysis of Mechano-Sensitive Ion Channels in Human Hearts: Chamber- and Disease-Preferential mRNA Expression. Int. J. Mol. Sci. 2023, 24, 10961. https://doi.org/10.3390/ijms241310961
Darkow E, Yusuf D, Rajamani S, Backofen R, Kohl P, Ravens U, Peyronnet R. Meta-Analysis of Mechano-Sensitive Ion Channels in Human Hearts: Chamber- and Disease-Preferential mRNA Expression. International Journal of Molecular Sciences. 2023; 24(13):10961. https://doi.org/10.3390/ijms241310961
Chicago/Turabian StyleDarkow, Elisa, Dilmurat Yusuf, Sridharan Rajamani, Rolf Backofen, Peter Kohl, Ursula Ravens, and Rémi Peyronnet. 2023. "Meta-Analysis of Mechano-Sensitive Ion Channels in Human Hearts: Chamber- and Disease-Preferential mRNA Expression" International Journal of Molecular Sciences 24, no. 13: 10961. https://doi.org/10.3390/ijms241310961