
Phloem Sap Composition: What Have We Learnt from Metabolomics?
 , ,
, ,
Abstract
:1. Introduction
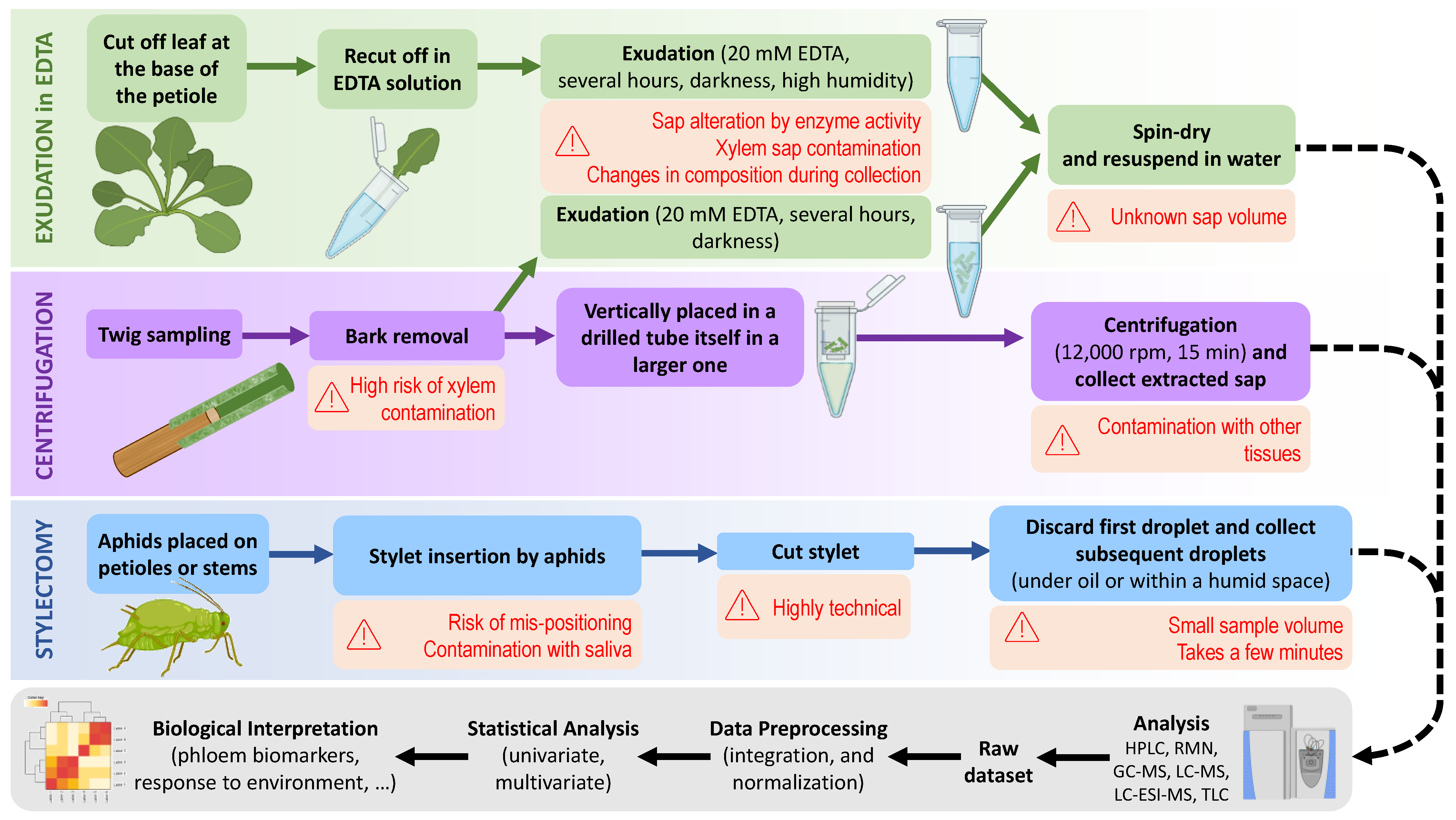
2. Phloem Sap Sampling Methods for Metabolomics: Overview
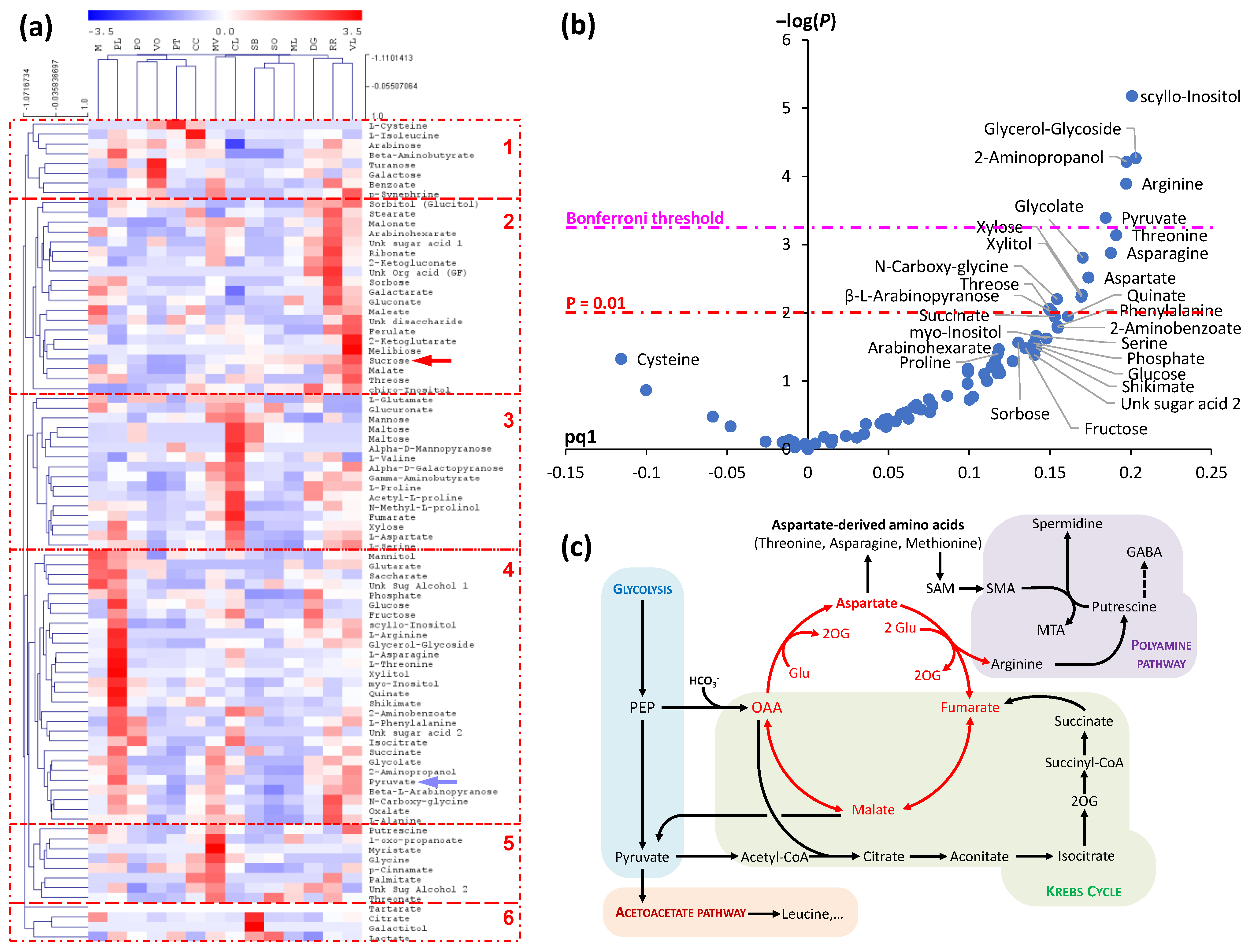
3. Phloem Sap Metabolites

{kind=link}
{kind=link}
{kind=link}
| Sap Sampling Method | Analytical Technique | Species | Observed Metabolites | References |
|---|---|---|---|---|
| Bark sampling | GC-MS | Picea abies | Carbohydrates | [73] |
| Bark sampling | GC-MS | Quercus cerris | Carbohydrates, fatty acids and organic acids | [74] |
| Bark sampling and EDTA | GC-MS | Vitis sp. | Amino acids, carbohydrates, organic acids and sugar alcohol | [64] |
| Centrifugation | GC-MS | Morus multicaulis | Amino acids, carbohydrates, organic acids, phytohormones and sugar alcohol | [75] |
| Centrifugation | GC-MS | Prunus sp. | Amino acids, carbohydrates, organic acids and sugar alcohol | [68] |
| Centrifugation | Raman spectroscopy | Quercus rubra | Carbohydrates | [76] |
| Centrifugation | GC-MS | Citrus sp. | Amino acids, fatty acids, organic acids and sugar alcohol | [77] |
| Centrifugation | GC-MS | Citrus sp. | Amino acids, carbohydrates, fatty acids, organic acids, phytohormones, sugar acids and sugar alcohol | [59] |
| Centrifugation | GC-MS | Citrus sp. | Fatty acids | [78] |
| EDTA | GC-MS | Arabidopsis thaliana | Amino acids, carbohydrates, organic acids and sugar alcohol | [65] |
| EDTA | HPLC | Arabidopsis thaliana and Sinapis alba | Amino acids | [79] |
| EDTA | LC-MS | Arabidopsis thaliana | Amino acids, carbohydrates, fatty acids and organic acids | [80] |
| EDTA | HPLC | Solanum tuberosum | Amino acids and carbohydrates | [60] |
| EDTA | Chromatography | Chenopodium rubrum, Perilla crispa and Pharbitis nil | Carbohydrates and sugar alcohols | [44] |
| EDTA | Radio-labelling | Anemone sylvestris, Centranthus ruber, Digitalis purpurea and Pulsatilla vulgaris | Carbohydrates | [71] |
| EDTA | Automated amino acid analyser | Picea abies, Fagus sylvatica | Amino acids and carbohydrates | [81] |
| EDTA | GC-MS and UHPLC-FLD | Plantago major and Poa annua | Amino acids, carbohydrates, organic acids and sugar alcohols | [63] |
| EDTA | HPLC | Solanum lycopersicum | Amino acids | [47] |
| EDTA | GC-MS | Zea mays | Amino acids, carbohydrates, organic acids and sugar alcohols | [62] |
| EDTA and centrifugation | GC-MS | Citrus sinensis | Amino acids, carbohydrates, fatty acids, organic acids and sugar alcohols | [45] |
| EDTA and isotope composition | HPLC and automated amino acid analyser | Fagus sylvatica | Amino acids and carbohydrates | [82] |
| EDTA-HEPES | Plate reader (enzymatic assay) | Solanum lycopersicum | Carbohydrates | [83] |
| Incision | GC-MS | Cucurbita maxima | Amino acids, carbohydrates and organic acids | [66] |
| Incision | Chromatography, FCR and amino acid analyser | Ricinus communis | Amino acids, carbohydrates and phytohormones | [55] |
| Incision | Refractometry and amino acid analyser | Nicotina glauca | Amino acids and carbohydrates | [84] |
| Incision | HPLC | Eucalyptus globulus | Amino acids, carbohydrates and organic acids | [58] |
| Incision | HPLC and refractometry | Ricinus communis | Amino acids, carbohydrates and organic acids | [21] |
| Incision | GC-MS | Cucumis sativus | Amino acids, carbohydrates, organic acids and sugar alcohols | [67] |
| Incision | GC-MS | Cucurbita maxima | Fatty acids | [85] |
| Incision | Chromatography | Robinia pseudoacacia, Quercus borealis, Quercus robur and Fraxinus americana | Amino acids and carbohydrates | [86] |
| Stylectomy | Automated amino acid analyser | Medicago sativa | Amino acids | [87] |
| Stylectomy | HPLC | Triticum aestivum | Amino acids and carbohydrates | [54] |
| Stylectomy | HPLC and fluospectrometry | Oryza sativa | Amino acids, carbohydrates and nucleotides | [52] |
| Stylectomy | CE-LIF | Arabidopsis thaliana | Amino acids | [88] |
| Stylectomy | HPLC | Zea mays | Amino acids | [89] |
| Stylectomy | HPLC | Brassica napus | Amino acids and carbohydrates | [51] |
| Stylectomy | HPLC | Plantago major, Plantago maritima, Prunus persica and Apium graveolens | Carbohydrates and sugar alcohols | [72] |
| Stylectomy | HPLC | Zea mays | Amino acids, carbohydrates and nucleotides | [50] |
| Stylectomy | HPLC | Quercus robur and Fraxinus excelsior | Carbohydrates and sugar alcohols | [56] |
| Stylectomy | GC-MS | Triticum aestivum | Amino acids, carbohydrates and organic acids | [69] |
| Stylectomy | HPLC | Trifolium pratense, Medicago sativa, Vicia faba and Pisum satiuum | Amino acids | [90] |
| Stylectomy | HPLC | Brassica napus | Amino acids and carbohydrates | [91] |
| Stylectomy | Non-aqueous fractionation | Hordeum vulgare | Amino acids and carbohydrates | [53] |
| Stylectomy | Enzymatic assay and LC | Beta vulgaris | Amino acids and carbohydrates | [61] |
| Stylectomy | HPLC | Oryza sativa | Phytohormones | [92] |
| Stylectomy and 11C radiotracing | HPLC | Zea mays | Amino acids and carbohydrates | [93] |
| Stylectomy and EDTA | HPLC | Fagus sylvatica, Magnolia kobus and Gnetum gnemon | Carbohydrates and sugar alcohols | [57] |
| Stylectomy and EDTA | Chromatography, amino acid analyser and SDS-PAGE | Lactuca sativa | Amino acids and carbohydrates | [94] |
4. Metabolic Pathways Reflected by Phloem Sap Metabolome
5. Phloem Sap Metabolites and Plant Resistance to Environmental Cues
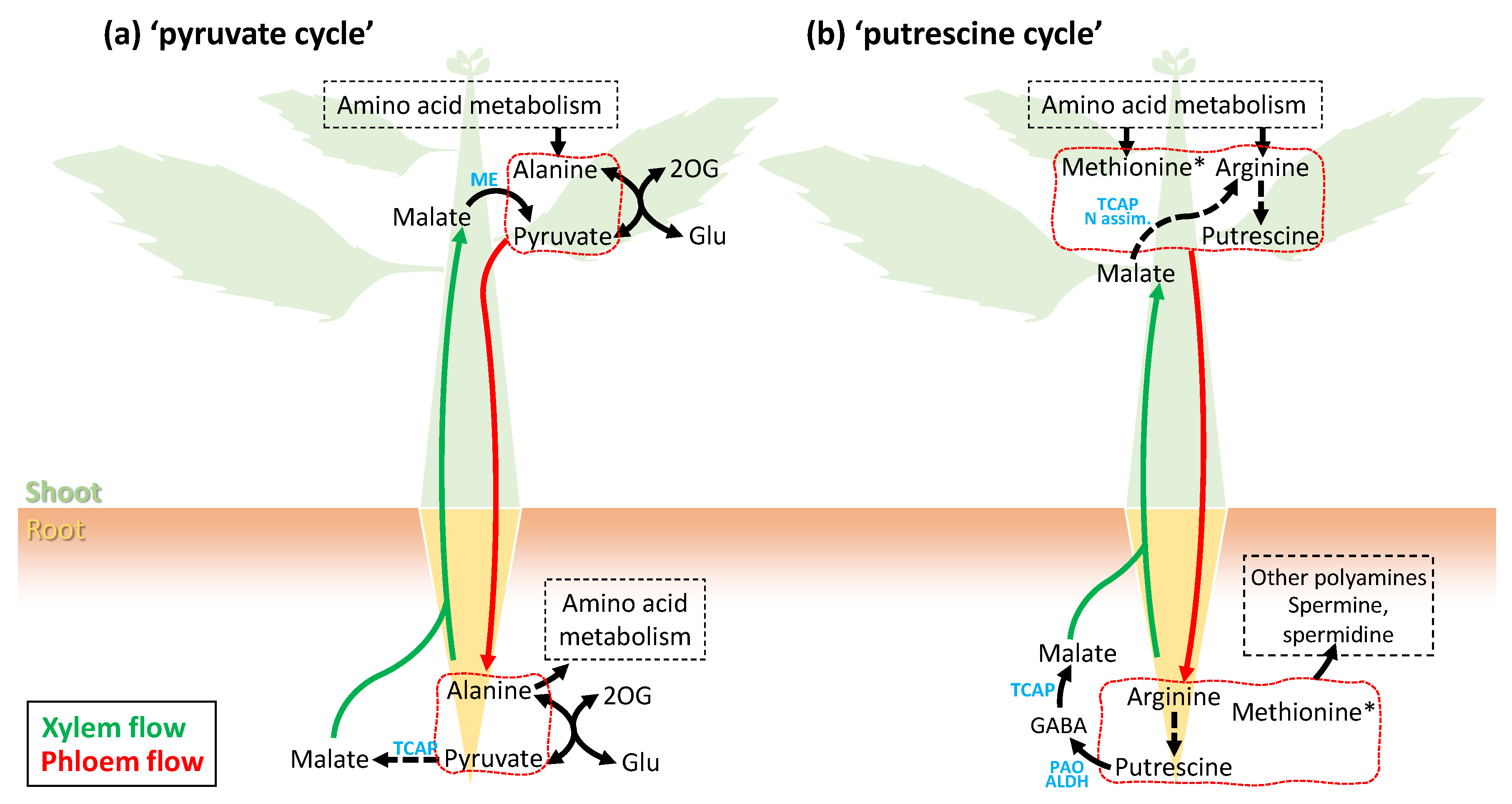
6. Phloem Sap Metabolome: Unforeseen Whole-Plant Metabolic Cycles?
7. Conclusions and Perspectives
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Dinant, S.; Lemoine, R. The phloem pathway: New issues and old debates. Comptes Rendus Biol. 2010, 333, 307–319. [Google Scholar] [CrossRef] [PubMed]
- Evert, R.F. Phloem Structure and Histochemistry. Annu. Rev. Plant Physiol. 1977, 28, 199–222. [Google Scholar] [CrossRef]
- Cronshaw, J. Phloem Structure and Function. Annu. Rev. Plant Physiol. 1981, 32, 465–484. [Google Scholar] [CrossRef]
- Golecki, B.; Schulz, A.; Thompson, G.A. Translocation of Structural P Proteins in the Phloem. Plant Cell 1999, 11, 127–140. [Google Scholar] [CrossRef] [Green Version]
- Ernst, A.M.; Jekat, S.B.; Zielonka, S.; Muller, B.; Neumann, U.; Ruping, B.; Twyman, R.M.; Krzyzanek, V.; Prufer, D.; Noll, G.A. Sieve element occlusion (SEO) genes encode structural phloem proteins involved in wound sealing of the phloem. Proc. Natl. Acad. Sci. USA 2012, 109, E1980–E1989. [Google Scholar] [CrossRef] [Green Version]
- Sellier, D.; Harrington, J.J. Phloem transport in trees: A generic surface model. Ecol. Model. 2014, 290, 102–109. [Google Scholar] [CrossRef]
- Holbrook, N.M.; Knoblauch, M. Editorial overview: Physiology and metabolism: Phloem: A supracellular highway for the transport of sugars, signals, and pathogens. Curr. Opin. Plant Biol. 2018, 43, iii–vii. [Google Scholar] [CrossRef]
- Weiler, E.W. Determination of phytohormones in phloem exudate from tree species by radioimmunoassay. Planta 1981, 152, 168–170. [Google Scholar] [CrossRef]
- Simpson, R.J. Nitrogen Redistribution during Grain Growth in Wheat (Triticum aestivum L.) 1: IV. Development of a Quantitative Model of the Translocation of Nitrogen to the Grain. Plant Physiol. 1983, 71, 7–14. [Google Scholar] [CrossRef] [Green Version]
- Rennenberg, H. Analysis of uptake and allocation of nitrogen and sulphur compounds by trees in the field. J. Exp. Bot. 1996, 47, 1491–1498. [Google Scholar] [CrossRef]
- Anderson, J.W. Sulphur Distribution and Redistribution in Vegetative and Generative Plants. In Sulphur in Plants; Springer: Dordrecht, The Netherlands, 2003; pp. 113–134. [Google Scholar] [CrossRef]
- Youssefi, F.; Brown, P.; Weinbaum, S. Relationship between tree nitrogen status, xylem and phloem sap amino acid concentrations, and apparent soil nitrogen uptake by almond trees (Prunus dulcis). J. Hortic. Sci. Biotechnol. 2015, 75, 62–68. [Google Scholar] [CrossRef]
- Pate, J.S.; Jeschke, W.D. Mineral uptake and transport in xylem and phloem of the proteaceous tree, Banksia prionotes. Plant Soil 1993, 155, 273–276. [Google Scholar] [CrossRef]
- Hoffmann-Benning, S.; Gage, D.A.; McIntosh, L.; Kende, H.; Zeevaart, J.A. Comparison of peptides in the phloem sap of flowering and non-flowering Perilla and lupine plants using microbore HPLC followed by matrix-assisted laser desorption/ionization time-of-flight mass spectrometry. Planta 2002, 216, 140–147. [Google Scholar] [CrossRef]
- Rodriguez-Celma, J.; Ceballos-Laita, L.; Grusak, M.A.; Abadia, J.; Lopez-Millan, A.F. Plant fluid proteomics: Delving into the xylem sap, phloem sap and apoplastic fluid proteomes. Biochim. Biophys. Acta 2016, 1864, 991–1002. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cankui, Z.; Turgeon, R. Mechanisms of Phloem Loading. Curr. Opin. Plant Biol. 2018, 43, 71–75. [Google Scholar] [CrossRef]
- Turgeon, R.; Wolf, S. Phloem transport: Cellular pathways and molecular trafficking. Annu. Rev. Plant Biol. 2009, 60, 207–221. [Google Scholar] [CrossRef] [Green Version]
- Dinant, S.; Kehr, J. Sampling and analysis of phloem sap. Methods Mol. Biol. 2013, 953, 185–194. [Google Scholar] [CrossRef]
- Hall, S.M.; Baker, D.A.; Milburn, J.A. Phloem transport of 14C-labelled assimilates in Ricinus. Planta 1971, 100, 200–207. [Google Scholar] [CrossRef] [PubMed]
- Meller, S.; Frossard, E.; Luster, J. Phosphorus Allocation to Leaves of Beech Saplings Reacts to Soil Phosphorus Availability. Front. Plant Sci. 2019, 10, 744. [Google Scholar] [CrossRef] [Green Version]
- Peuke, A. Correlations in concentrations, xylem and phloem flows, and partitioning of elements and ions in intact plants. A summary and statistical re-evaluation of modelling experiments in Ricinus communis. J. Exp. Bot. 2010, 61, 635–655. [Google Scholar] [CrossRef] [Green Version]
- Zhang, B.; Tolstikov, V.; Turnbull, C.; Hicks, L.M.; Fiehn, O. Divergent metabolome and proteome suggest functional independence of dual phloem transport systems in cucurbits. Proc. Natl. Acad. Sci. USA 2010, 107, 13532–13537. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brandt, S.; Kehr, J.; Walz, C.; Imlau, A.; Willmitzer, L.; Fisahn, J. Technical Advance: A rapid method for detection of plant gene transcripts from single epidermal, mesophyll and companion cells of intact leaves. Plant J. 1999, 20, 245–250. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Raps, A.; Kehr, J.; Gugerli, P.; Moar, W.J.; Bigler, F.; Hilbeck, A. Immunological analysis of phloem sap of Bacillus thuringiensis corn and of the nontarget herbivore Rhopalosiphum padi (Homoptera: Aphididae) for the presence of Cry1Ab. Mol. Ecol. 2001, 10, 525–533. [Google Scholar] [CrossRef] [PubMed]
- Asano, T.; Masumura, T.; Kusano, H.; Kikuchi, S.; Kurita, A.; Shimada, H.; Kadowaki, K. Construction of a specialized cDNA library from plant cells isolated by laser capture microdissection: Toward comprehensive analysis of the genes expressed in the rice phloem. Plant J. 2002, 32, 401–408. [Google Scholar] [CrossRef] [PubMed]
- Yu, Y.; Lashbrook, C.C.; Hannapel, D.J. Tissue integrity and RNA quality of laser microdissected phloem of potato. Planta 2007, 226, 797–803. [Google Scholar] [CrossRef] [PubMed]
- Deeken, R.; Ache, P.; Kajahn, I.; Klinkenberg, J.; Bringmann, G.; Hedrich, R. Identification of Arabidopsis thaliana phloem RNAs provides a search criterion for phloem-based transcripts hidden in complex datasets of microarray experiments. Plant J. 2008, 55, 746–759. [Google Scholar] [CrossRef] [Green Version]
- Brady, S.M.; Orlando, D.A.; Lee, J.-Y.; Wang, J.Y.; Koch, J.; Dinneny, J.R.; Mace, D.; Ohler, U.; Benfey, P.N. A High-Resolution Root Spatiotemporal Map Reveals Dominant Expression Patterns. Science 2007, 318, 801–806. [Google Scholar] [CrossRef]
- Iyer-Pascuzzi, A.S.; Benfey, P.N. Fluorescence-Activated Cell Sorting in Plant Developmental Biology. In Plant Developmental Biology: Methods and Protocols; Hennig, L., Köhler, C., Eds.; Methods in Molecular Biology; Humana Press: Totowa, NJ, USA, 2010; pp. 313–319. [Google Scholar]
- Biddulph, O.; Cory, R. An Analysis of Translocation in the Phloem of the Bean Plant Using Tho, P32, And C14. Plant Physiol. 1957, 32, 608. [Google Scholar] [CrossRef] [Green Version]
- Bourgis, F.; Roje, S.; Nuccio, M.L.; Fisher, D.B.; Tarczynski, M.C.; Li, C.; Herschbach, C.; Rennenberg, H.; Pimenta, M.J.; Shen, T.L.; et al. S-methylmethionine plays a major role in phloem sulfur transport and is synthesized by a novel type of methyltransferase. Plant Cell 1999, 11, 1485–1498. [Google Scholar] [CrossRef] [Green Version]
- Minchin, P.E.H.; Thorpe, M.R. Measurement of Unloading and Reloading of Photo-assimilate within the Stem of Bean. J. Exp. Bot. 1987, 38, 211–220. [Google Scholar] [CrossRef]
- Kennedy, J.S.; Mittler, T.E. A Method of obtaining Phloem Sap via the Mouth-parts of Aphids. Nature 1953, 171, 528. [Google Scholar] [CrossRef]
- Dinant, S.; Bonnemain, J.-L.; Girousse, C.; Kehr, J. Phloem sap intricacy and interplay with aphid feeding. Comptes Rendus Biol. 2010, 333, 504–515. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- van Bel, A.J.; Knoblauch, M.; Furch, A.C.; Hafke, J.B. Questions on phloem biology. 1. Electropotential waves, Ca2+ fluxes and cellular cascades along the propagation pathway. Plant Sci. 2011, 181, 210–218. [Google Scholar] [CrossRef] [PubMed]
- Pompon, J.; Quiring, D.; Goyer, C.; Giordanengo, P.; Pelletier, Y. A phloem-sap feeder mixes phloem and xylem sap to regulate osmotic potential. J. Insect Physiol. 2011, 57, 1317–1322. [Google Scholar] [CrossRef]
- Prado, E.; Tjallingii, W.F. Aphid activities during sieve element punctures. Entomol. Exp. Appl. 1994, 72, 157–165. [Google Scholar] [CrossRef]
- Tjallingii, W.F. Membrane potentials as an indication for plant cell penetration by aphid stylets. Entomol. Exp. Appl. 1985, 38, 187–193. [Google Scholar] [CrossRef]
- Tjallingii, W.F.; Esch, T.H. Fine structure of aphid stylet routes in plant tissues in correlation with EPG signals. Physiol. Entomol. 1993, 18, 317–328. [Google Scholar] [CrossRef]
- Tjallingii, W.F. Electronic Recording of Penetration Behaviour by Aphids. Entomol. Exp. Appl. 1978, 24, 721–730. [Google Scholar] [CrossRef]
- McLean, D.L.; Kinsey, M.G. A Technique for Electronically Recording Aphid Feeding and Salivation. Nature 1964, 202, 1358–1359. [Google Scholar] [CrossRef]
- Spiller, N.j.; Koenders, L.; Tjallingii, W.f. Xylem ingestion by aphids—A strategy for maintaining water balance. Entomol. Exp. Appl. 1990, 55, 101–104. [Google Scholar] [CrossRef]
- Seo, B.Y.; Kwon, Y.-H.; Jung, J.K.; Kim, G.-H. Electrical penetration graphic waveforms in relation to the actual positions of the stylet tips of Nilaparvata lugens in rice tissue. J. Asia-Pac. Entomol. 2009, 12, 89–95. [Google Scholar] [CrossRef]
- King, R.W.; Zeevaart, J.A.D. Enhancement of Phloem Exudation from Cut Petioles by Chelating Agents 1. Plant Physiol. 1974, 53, 96–103. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hijaz, F.; Killiny, N. Collection and chemical composition of phloem sap from Citrus sinensis L. Osbeck (sweet orange). PLoS ONE 2014, 9, e101830. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Killiny, N. Collection of the phloem sap, pros and cons. Plant Signal. Behav. 2019, 14, 1618181. [Google Scholar] [CrossRef] [PubMed]
- Valle, E.M.; Boggio, S.B.; Heldt, H.W. Free Amino Acid Composition of Phloem Sap and Growing Fruit of Lycopersicon esculentum. Plant Cell Physiol. 1998, 39, 458–461. [Google Scholar] [CrossRef] [Green Version]
- Pahlow, S.; Ostendorp, A.; Krussel, L.; Kehr, J. Phloem Sap Sampling from Brassica napus for 3D-PAGE of Protein and Ribonucleoprotein Complexes. J. Vis. Exp. 2018, 2018, e57097. [Google Scholar] [CrossRef] [Green Version]
- Lohaus, G. Review primary and secondary metabolites in phloem sap collected with aphid stylectomy. J. Plant Physiol. 2022, 271, 153645. [Google Scholar] [CrossRef]
- Ohshima, T.; Hayashi, H.; Chino, M. Collection and Chemical Composition of Pure Phloem Sap from Zea mays L. Plant Cell Physiol. 1990, 31, 735–737. [Google Scholar] [CrossRef]
- Lohaus, G.; Moellers, C. Phloem transport of amino acids in two Brassica napus L. genotypes and one B. carinata genotype in relation to their seed protein content. Planta 2000, 211, 833–840. [Google Scholar] [CrossRef]
- Hayashi, H.; Chino, M. Chemical Composition of Phloem Sap from the Uppermost Internode of the Rice Plant. Plant Cell Physiol. 1990, 31, 247–251. [Google Scholar] [CrossRef] [Green Version]
- Winter, H.; Lohaus, G.; Heldt, H.W. Phloem Transport of Amino Acids in Relation to their Cytosolic Levels in Barley Leaves 1. Plant Physiol. 1992, 99, 996–1004. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hayashi, H.; Chino, M. Collection of Pure Phloem Sap from Wheat and its Chemical Composition. Plant Cell Physiol. 1986, 27, 1387–1393. [Google Scholar] [CrossRef]
- Hall, S.M.; Baker, D.A. The chemical composition of Ricinus phloem exudate. Planta 1972, 106, 131–140. [Google Scholar] [CrossRef] [PubMed]
- Oner-Sieben, S.; Lohaus, G. Apoplastic and symplastic phloem loading in Quercus robur and Fraxinus excelsior. J. Exp. Bot. 2014, 65, 1905–1916. [Google Scholar] [CrossRef] [Green Version]
- Fink, D.; Dobbelstein, E.; Barbian, A.; Lohaus, G. Ratio of sugar concentrations in the phloem sap and the cytosol of mesophyll cells in different tree species as an indicator of the phloem loading mechanism. Planta 2018, 248, 661–673. [Google Scholar] [CrossRef]
- Pate, J.; Shedley, E.; Arthur, D.; Adams, M. Spatial and temporal variations in phloem sap composition of plantation-grown Eucalyptus globulus. Oecologia 1998, 117, 312–322. [Google Scholar] [CrossRef]
- Killiny, N. Metabolite signature of the phloem sap of fourteen citrus varieties with different degrees of tolerance to Candidatus liberibacter asiaticus. Physiol. Mol. Plant Pathol. 2017, 97, 20–29. [Google Scholar] [CrossRef]
- Heineke, D.; Sonnewald, U.; Büssis, D.; Günter, G.; Leidreiter, K.; Wilke, I.; Raschke, K.; Willmitzer, L.; Heldt, H.W. Apoplastic Expression of Yeast-Derived Invertase in Potato: Effects on Photosynthesis, Leaf Solute Composition, Water Relations, and Tuber Composition. Plant Physiol. 1992, 100, 301–308. [Google Scholar] [CrossRef] [Green Version]
- Winzer, T.; Lohaus, G.; Heldt, H.-W. Influence of phloem transport, N-fertilization and ion accumulation on sucrose storage in the taproots of fodder beet and sugar beet. J. Exp. Bot. 1996, 47, 863–870. [Google Scholar] [CrossRef] [Green Version]
- Yesbergenova-Cuny, Z.; Dinant, S.; Martin-Magniette, M.-L.; Quilleré, I.; Armengaud, P.; Monfalet, P.; Lea, P.J.; Hirel, B. Genetic variability of the phloem sap metabolite content of maize (Zea mays L.) during the kernel-filling period. Plant Sci. 2016, 252, 347–357. [Google Scholar] [CrossRef]
- Stallmann, J.; Schweiger, R. Effects of Arbuscular Mycorrhiza on Primary Metabolites in Phloem Exudates of Plantago major and Poa annua and on a Generalist Aphid. Int. J. Mol. Sci. 2021, 22, 13086. [Google Scholar] [CrossRef] [PubMed]
- Tedesco, S.; Erban, A.; Gupta, S.; Kopka, J.; Fevereiro, P.; Kragler, F.; Pina, A. The Impact of Metabolic Scion-Rootstock Interactions in Different Grapevine Tissues and Phloem Exudates. Metabolites 2021, 11, 349. [Google Scholar] [CrossRef] [PubMed]
- Chardon, F.; De Marco, F.; Marmagne, A.; Le Hir, R.; Vilaine, F.; Bellini, C.; Dinant, S. Natural variation in the long-distance transport of nutrients and photoassimilates in response to N availability. J. Plant Physiol. 2022, 273, 153707. [Google Scholar] [CrossRef] [PubMed]
- Fiehn, O. Metabolic networks of Cucurbita maxima phloem. Phytochemistry 2003, 62, 875–886. [Google Scholar] [CrossRef]
- Sun, J.; Li, Q.; Xu, H.; Zhang, W. Analysis of Metabolomic Changes in Xylem and Phloem Sap of Cucumber under Phosphorus Stresses. Metabolites 2022, 12, 361. [Google Scholar] [CrossRef]
- Gallinger, J.; Gross, J. Phloem Metabolites of Prunus sp. Rather than Infection with Candidatus phytoplasma prunorum Influence Feeding Behavior of Cacopsylla pruni Nymphs. J. Chem. Ecol. 2020, 46, 756–770. [Google Scholar] [CrossRef] [Green Version]
- Palmer, L.J.; Dias, D.A.; Boughton, B.; Roessner, U.; Graham, R.D.; Stangoulis, J.R. Metabolite profiling of wheat (Triticum aestivum L.) phloem exudate. Plant Methods 2014, 10, 27. [Google Scholar] [CrossRef] [Green Version]
- Xie, H.; Shi, F.; Li, J.; Yu, M.; Yang, X.; Li, Y.; Fan, J. The Reciprocal Effect of Elevated CO2 and Drought on Wheat-Aphid Interaction System. Front. Plant Sci. 2022, 13, 853220. [Google Scholar] [CrossRef]
- Liu, D.D.; Chao, W.M.; Turgeon, R. Transport of sucrose, not hexose, in the phloem. J. Exp. Bot. 2012, 63, 4315–4320. [Google Scholar] [CrossRef] [Green Version]
- Nadwodnik, J.; Lohaus, G. Subcellular concentrations of sugar alcohols and sugars in relation to phloem translocation in Plantago major, Plantago maritima, Prunus persica, and Apium graveolens. Planta 2008, 227, 1079–1089. [Google Scholar] [CrossRef] [Green Version]
- Jyske, T.; Keinänen, S.; Hölttä, T.; Lintunen, A.; Pranovich, A.; Laakso, T.; Suuronen, J.-P.; da Silva Viana, G.; Pakkala, T. Pilot Study of Sap Properties of Norway Spruce (Picea abies (L.) Karst.) Trees Used and Not Used for Sap-Feeding by Three-Toed Woodpeckers (Picoides tridactylus). Forests 2022, 13, 1681. [Google Scholar] [CrossRef]
- Sen, A.; Miranda, I.; Santos, S.; Grac, J.; Pereira, H. The chemical composition of cork and phloem in the rhytidome of Quercus cerris bark. Ind. Crops Prod. 2010, 31, 417–422. [Google Scholar] [CrossRef]
- Gai, Y.-P.; Han, X.-J.; Li, Y.-Q.; Yuan, C.-Z.; Mo, Y.-Y.; Guo, F.-Y.; Liu, Q.-X.; Ji, X.-L. Metabolomic analysis reveals the potential metabolites and pathogenesis involved in mulberry yellow dwarf disease. Plant Cell Environ. 2014, 37, 1474–1490. [Google Scholar] [CrossRef] [PubMed]
- Gersony, J.T.; McClelland, A.; Holbrook, N.M. Raman spectroscopy reveals high phloem sugar content in leaves of canopy red oak trees. New Phytol. 2021, 232, 418–424. [Google Scholar] [CrossRef] [PubMed]
- Killiny, N.; Hijaz, F. Amino acids implicated in plant defense are higher in Candidatus liberibacter asiaticus-tolerant Citrus varieties. Plant Signal. Behav. 2016, 11, e1171449. [Google Scholar] [CrossRef] [Green Version]
- Valim, M.F.; Killiny, N. Occurrence of free fatty acids in the phloem sap of different citrus varieties. Plant Signal. Behav. 2017, 12, e1327497. [Google Scholar] [CrossRef]
- Corbesier, L.; Havelange, A.; Lejeune, P.; Bernier, G.; Périlleux, C. N content of phloem and xylem exudates during the transition to flowering in Sinapis alba and Arabidopsis thaliana. Plant Cell Environ. 2001, 24, 367–375. [Google Scholar] [CrossRef]
- Guelette, B.S.; Benning, U.F.; Hoffmann-Benning, S. Identification of lipids and lipid-binding proteins in phloem exudates from Arabidopsis thaliana. J. Exp. Bot. 2012, 63, 3603–3616. [Google Scholar] [CrossRef] [Green Version]
- Schneider, S.; GessLer, A.; Weber, P.; Sengbusch, D.; Hanemann, U.; Rennenberg, H. Soluble N compounds in trees exposed to high loads of N: A comparison of spruce (Picea abies) and beech (Fagus sylvatica) grown under field conditions. New Phytol. 1996, 134, 103–114. [Google Scholar] [CrossRef]
- Gessler, A.; Rennenberg, H.; Keitel, C. Stable isotope composition of organic compounds transported in the phloem of European beech--evaluation of different methods of phloem sap collection and assessment of gradients in carbon isotope composition during leaf-to-stem transport. Plant Biol. 2004, 6, 721–729. [Google Scholar] [CrossRef]
- Lombardo, F.; Gramazio, P.; Ezura, H. Increase in Phloem Area in the Tomato hawaiian skirt Mutant Is Associated with Enhanced Sugar Transport. Genes 2021, 12, 932. [Google Scholar] [CrossRef] [PubMed]
- Hocking, P.J. The Composition of Phloem Exudate and Xylem Sap from Tree Tobacco (Nicotiana glauca Grah.). Ann. Bot. 1980, 45, 633–643. [Google Scholar] [CrossRef]
- Tombuloglu, H.; Anıl, I.; Akhtar, S.; Turumtay, H.; Sabit, H.; Slimani, Y.; Almessiere, M.; Baykal, A. Iron oxide nanoparticles translocate in pumpkin and alter the phloem sap metabolites related to oil metabolism. Sci. Hortic. 2020, 265, 109223. [Google Scholar] [CrossRef]
- Ziegler, H. Untersuchungen Über die Leitung und Sekretion der Assimilate. Planta 1956, 47, 447–500. [Google Scholar] [CrossRef]
- Girousse, C.; Bournoville, R.; Bonnemain, J.L. Water Deficit-Induced Changes in Concentrations in Proline and Some Other Amino Acids in the Phloem Sap of Alfalfa. Plant Physiol. 1996, 111, 109–113. [Google Scholar] [CrossRef] [Green Version]
- Hunt, E.; Gattolin, S.; Newbury, H.J.; Bale, J.S.; Tseng, H.-M.; Barrett, D.A.; Pritchard, J. A mutation in amino acid permease AAP6 reduces the amino acid content of the Arabidopsis sieve elements but leaves aphid herbivores unaffected. J. Exp. Bot. 2010, 61, 55–64. [Google Scholar] [CrossRef]
- Lohaus, G.; Büker, M.; Hußmann, M.; Soave, C.; Heldt, H.-W. Transport of amino acids with special emphasis on the synthesis and transport of asparagine in the Illinois Low Protein and Illinois High Protein strains of maize. Planta 1998, 205, 181–188. [Google Scholar] [CrossRef]
- Sandström, J.; Pettersson, J. Amino acid composition of phloem sap and the relation to intraspecific variation in pea aphid (Acyrthosiphon pisum) performance. J. Insect Physiol. 1994, 40, 947–955. [Google Scholar] [CrossRef]
- Tilsner, J.; Kassner, N.; Struck, C.; Lohaus, G. Amino acid contents and transport in oilseed rape (Brassica napus L.) under different nitrogen conditions. Planta 2005, 221, 328–338. [Google Scholar] [CrossRef]
- Yokota, T.; Nakayama, M.; Harasawa, I.; Sato, M.; Katsuhara, M.; Kawabe, S. Polyamines, indole-3-acetic acid and abscisic acid in rice phloem sap. Plant Growth Regul. 1994, 15, 125–128. [Google Scholar] [CrossRef]
- Babst, B.A.; Braun, D.M.; Karve, A.A.; Frank Baker, R.; Tran, T.M.; Kenny, D.J.; Rohlhill, J.; Knoblauch, J.; Knoblauch, M.; Lohaus, G.; et al. Sugar loading is not required for phloem sap flow in maize plants. Nat. Plants 2022, 8, 171–180. [Google Scholar] [CrossRef]
- van Helden, M.; Tjallingh, W.F.; van Beek, T.A. Phloem sap collection from lettuce (Lactuca sativa L.): Chemical comparison among collection methods. J. Chem. Ecol. 1994, 20, 3191–3206. [Google Scholar] [CrossRef]
- Touraine, B.; Muller, B.; Grignon, C. Effect of Phloem-Translocated Malate on NO3− Uptake by Roots of Intact Soybean Plants. Plant Physiol. 1992, 99, 1118–1123. [Google Scholar] [CrossRef] [Green Version]
- Benzioni, A.; Vaadia, Y.; Herman Lips, S. Correlations between Nitrate Reduction, Protein Synthesis and Malate Accumulation. Physiol. Plant. 1970, 23, 1039–1047. [Google Scholar] [CrossRef]
- Hoffmann-Benning, S. Collection and Analysis of Phloem Lipids. Methods Mol. Biol. 2021, 2295, 351–361. [Google Scholar] [CrossRef] [PubMed]
- Gaufichon, L.; Reisdorf-Cren, M.; Rothstein, S.; Chardon, F.; Suzuki, A. Biological functions of asparagine synthetase in plants. Plant Sci. 2010, 179, 141–153. [Google Scholar] [CrossRef]
- Gaufichon, L.; Masclaux-Daubresse, C.; Tcherkez, G.; Reisdorf-Cren, M.; Sakakibara, Y.; Hase, T.; Clement, G.; Avice, J.C.; Grandjean, O.; Marmagne, A.; et al. Arabidopsis thaliana ASN2 encoding asparagine synthetase is involved in the control of nitrogen assimilation and export during vegetative growth. Plant Cell Environ. 2013, 36, 328–342. [Google Scholar] [CrossRef] [PubMed]
- Kennecke, M.; Ziegler, H.; de Fekete, M.A. Enzyme activities in the sieve tube sap of Robinia pseudoacacia L. and of other tree species. Planta 1971, 98, 330–356. [Google Scholar] [CrossRef]
- Knoblauch, M. Sieve Tubes in Action. Plant Cell 1998, 10, 35–50. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Vasina, V.V.; Kraner, M.E.; Peters, W.S.; Sonnewald, U.; Knoblauch, M. Proteomics of isolated sieve tubes from Nicotiana tabacum: Sieve element-specific proteins reveal differentiation of the endomembrane system. Proc. Natl. Acad. Sci. USA 2022, 119, e2112755119. [Google Scholar] [CrossRef]
- Cui, J.; Peuke, A.D.; Limami, A.M.; Tcherkez, G. Why is phloem sap nitrate kept low? Plant Cell Environ. 2021, 44, 2838–2843. [Google Scholar] [CrossRef]
- Cui, J.; Abadie, C.; Carroll, A.; Lamade, E.; Tcherkez, G. Responses to K deficiency and waterlogging interact via respiratory and nitrogen metabolism. Plant Cell Environ. 2019, 42, 647–658. [Google Scholar] [CrossRef] [PubMed]
- Gajdanowicz, P.; Michard, E.; Sandmann, M.; Rocha, M.; Corrêa, L.G.G.; Ramírez-Aguilar, S.J.; Gomez-Porras, J.L.; González, W.; Thibaud, J.-B.; van Dongen, J.T. Potassium (K+) gradients serve as a mobile energy source in plant vascular tissues. Proc. Natl. Acad. Sci. USA 2011, 108, 864–869. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lothier, J.; Diab, H.; Cukier, C.; Limami, A.M.; Tcherkez, G. Metabolic Responses to Waterlogging Differ between Roots and Shoots and Reflect Phloem Transport Alteration in Medicago truncatula. Plants 2020, 9, 1373. [Google Scholar] [CrossRef] [PubMed]
- van Beusichem, M.L.; Nelemans, O.A.; Hinnen, M.G.J. Nitrogen: Nitrogen cycling in plant species differing in shoot/root reduction of nitrate. J. Plant Nutr. 1987, 10, 1723–1731. [Google Scholar] [CrossRef]
- Marschner, P.; Crowley, D.E.; Higashi, R.M. Root exudation and physiological status of a root-colonizing fluorescent pseudomonad in mycorrhizal and non-mycorrhizal pepper (Capsicum annuum L.). Plant Soil 1997, 189, 11–20. [Google Scholar] [CrossRef]
- Teskey, R.O.; Saveyn, A.; Steppe, K.; McGuire, M.A. Origin, fate and significance of CO2 in tree stems. New Phytol. 2008, 177, 17–32. [Google Scholar] [CrossRef]
- Trumbore, S.E.; Angert, A.; Kunert, N.; Muhr, J.; Chambers, J.Q. What’s the flux? Unraveling how CO2 fluxes from trees reflect underlying physiological processes. New Phytol. 2013, 197, 353–355. [Google Scholar] [CrossRef] [Green Version]
- Cui, J.; Pottosin, I.; Lamade, E.; Tcherkez, G. What is the role of putrescine accumulated under potassium deficiency? Plant Cell Environ. 2020, 43, 1331–1347. [Google Scholar] [CrossRef]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Broussard, L.; Abadie, C.; Lalande, J.; Limami, A.M.; Lothier, J.; Tcherkez, G. Phloem Sap Composition: What Have We Learnt from Metabolomics? Int. J. Mol. Sci. 2023, 24, 6917. https://doi.org/10.3390/ijms24086917
Broussard L, Abadie C, Lalande J, Limami AM, Lothier J, Tcherkez G. Phloem Sap Composition: What Have We Learnt from Metabolomics? International Journal of Molecular Sciences. 2023; 24(8):6917. https://doi.org/10.3390/ijms24086917
Chicago/Turabian StyleBroussard, Louis, Cyril Abadie, Julie Lalande, Anis M. Limami, Jérémy Lothier, and Guillaume Tcherkez. 2023. "Phloem Sap Composition: What Have We Learnt from Metabolomics?" International Journal of Molecular Sciences 24, no. 8: 6917. https://doi.org/10.3390/ijms24086917