
The Effect of Scale Insects on Growth Parameters of cv. Chardonnay and cv. Sauvignon Blanc Grapevines Grown in a Greenhouse

Abstract
:1. Introduction
2. Results
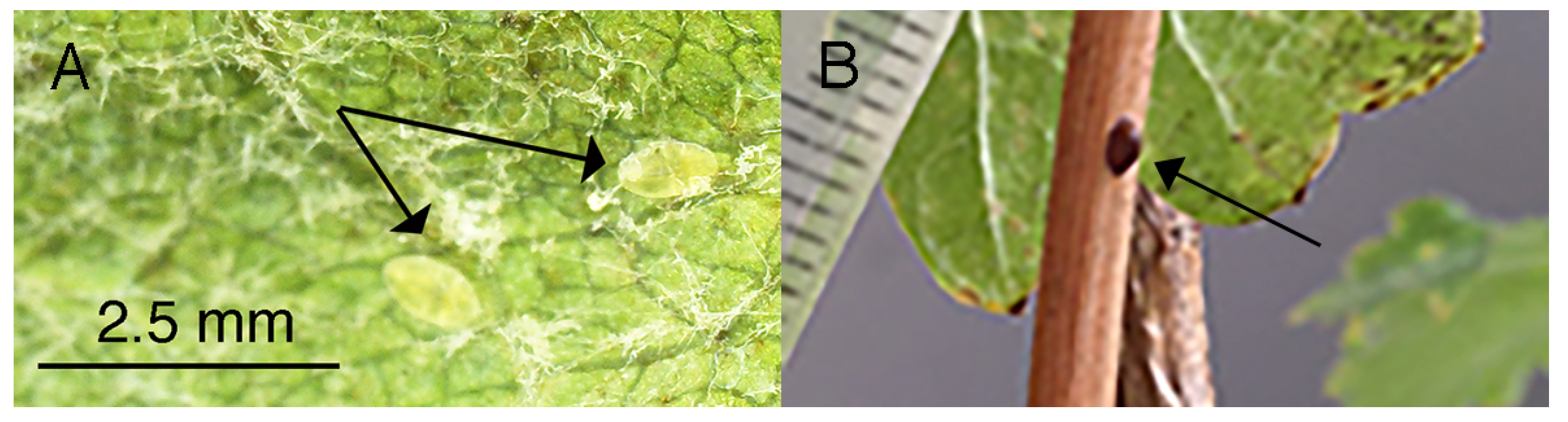
2.1. Scale Number and Plant Phenotypes
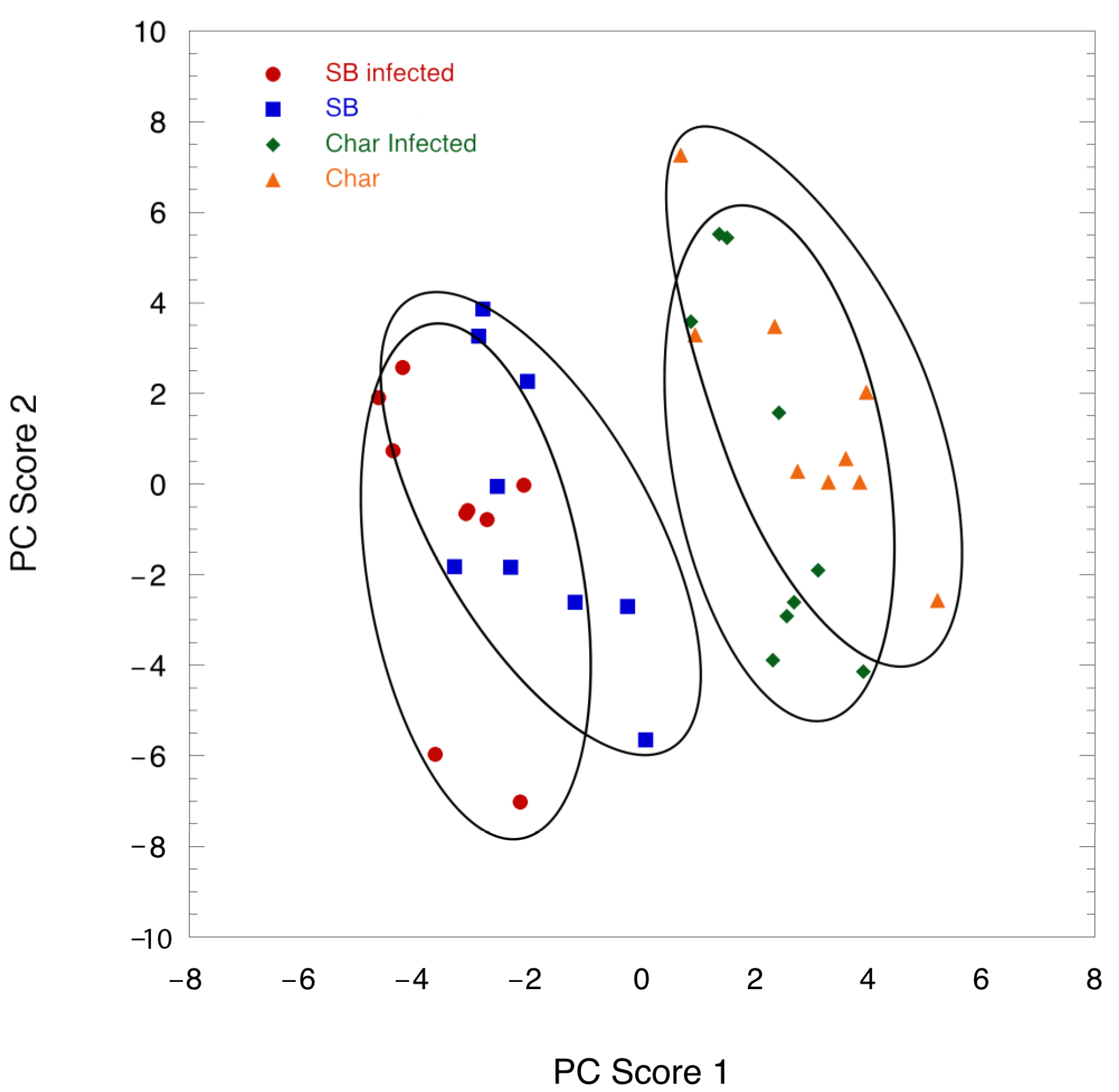
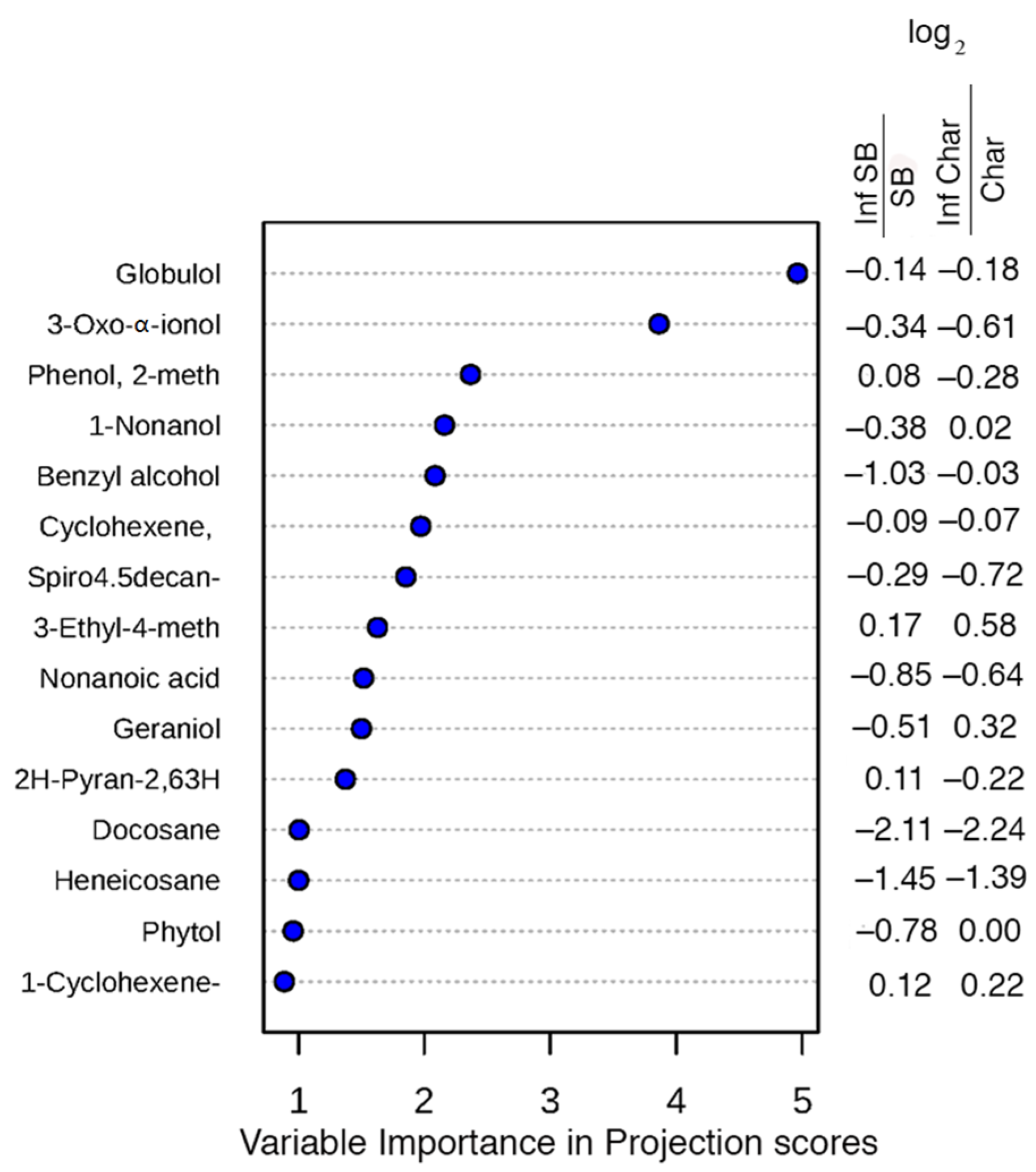
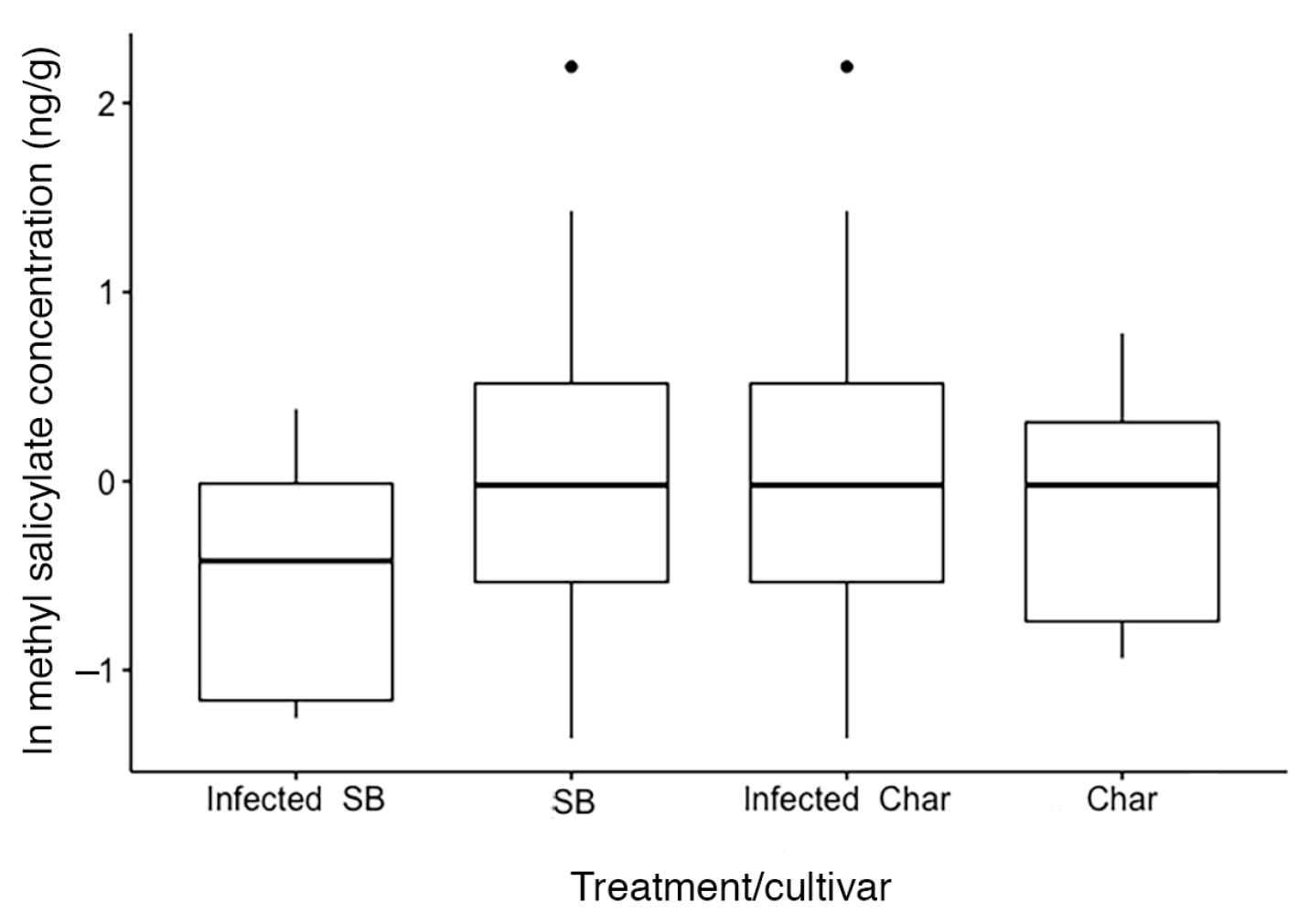
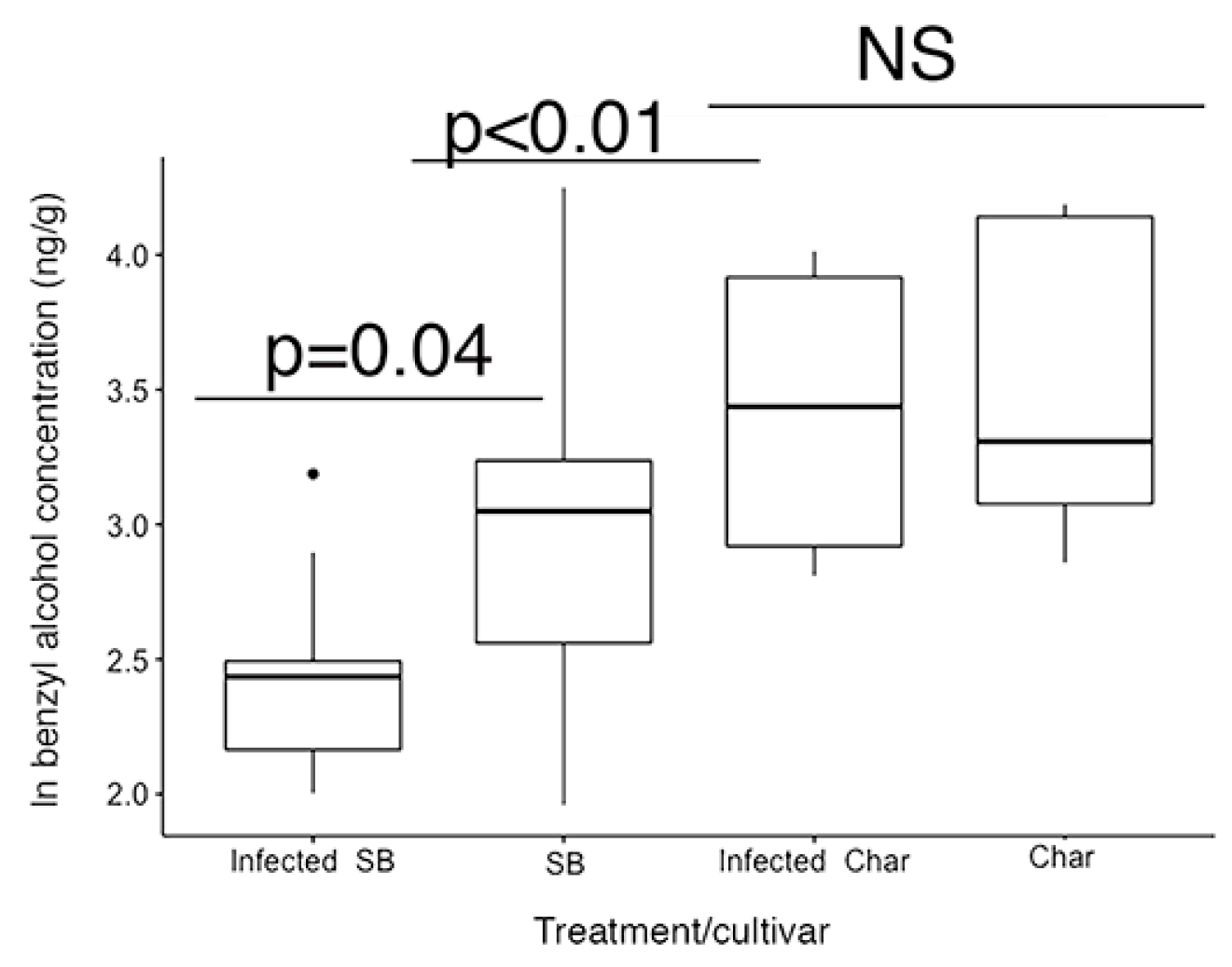
2.2. Leaf Volatile Chemistry
3. Discussion
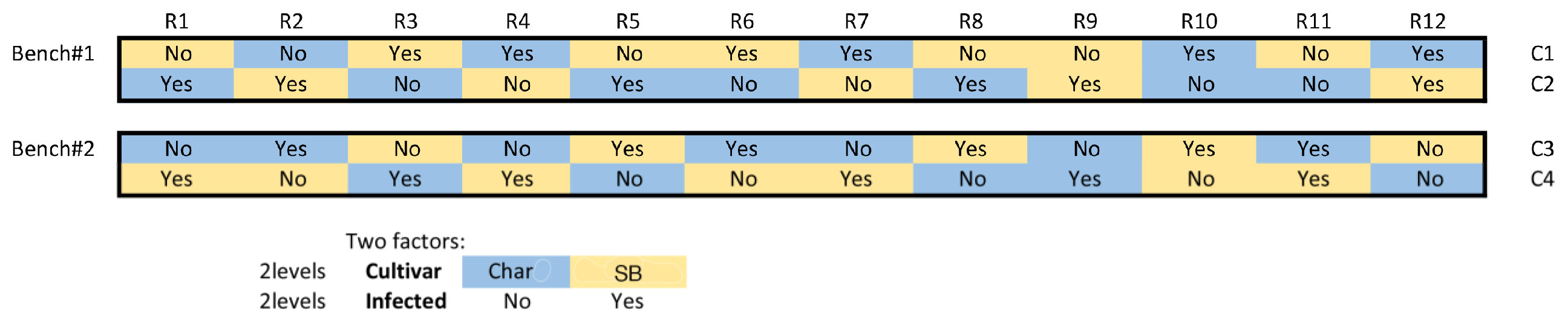
4. Materials and Methods
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Yactayo-Chang, J.P.; Tang, H.V.; Mendoza, J.; Christensen, S.A.; Block, A.K. Plant defense chemicals against insect pests. Agron. Basel 2020, 10, 1156. [Google Scholar] [CrossRef]
- Zogli, P.; Pingault, L.; Grover, S.; Louis, J. Ento(o)mics: The intersection of ‘omic’ approaches to decipher plant defense against sap-sucking insect pests. Curr. Opin. Plant Biol. 2020, 56, 153–161. [Google Scholar] [CrossRef] [PubMed]
- Zvereva, E.L.; Lanta, V.; Kozlov, M.V. Effects of sap-feeding insect herbivores on growth and reproduction of woody plants: A meta-analysis of experimental studies. Oecologia 2010, 163, 949–960. [Google Scholar] [CrossRef] [PubMed]
- Erb, M. Plant defenses against herbivory: Closing the fitness gap. Trends Plant Sci. 2018, 23, 187–194. [Google Scholar] [CrossRef] [PubMed]
- Aljbory, Z.; Chen, M.-S. Indirect plant defense against insect herbivores: A review. Insect. Sci. 2018, 25, 2–23. [Google Scholar] [CrossRef] [PubMed]
- Croteau, R. Biosynthesis of benzaldehyde, benzyl alcohol and benzyl benzoate from benzoic acid in cranberry (Vaccinium macrocarpon). J. Food Biochem. 1978, 1, 317–326. [Google Scholar] [CrossRef]
- Bernays, E.A.; Chapman, R.F. The role of food plants in the survival and development of Chortoicetes terminifera (Walker) under drought conditions. Aust. J. Zool. 1973, 21, 575–592. [Google Scholar] [CrossRef]
- Norkute, M.; Olsson, U.; Ninkovic, V. Aphids-induced plant volatiles affect diel foraging behavior of a ladybird beetle Coccinella septempunctata. Insect. Sci. 2019, 27, 1266–1275. [Google Scholar] [CrossRef] [Green Version]
- Lawo, N.C.; Weingart, G.J.F.; Schuhmacher, R.; Forneck, A. The volatile metabolome of grapevine roots: First insights into the metabolic response upon phylloxera attack. Plant Phys. Biochem. 2011, 49, 1059–1063. [Google Scholar] [CrossRef] [Green Version]
- Filgueiras, C.C.; Martins, A.D.; Pereira, R.V.; Willett, D.S. The ecology of salicylic acid signaling: Primary, secondary and tertiary effects with applications in agriculture. Int. J. Molec. Sci. 2019, 20, 5851. [Google Scholar] [CrossRef]
- This, P.; Lacombe, T.; Thomas, M.R. Historical origins and genetic diversity of wine grapes. Trends Genet. 2006, 22, 511–519. [Google Scholar] [CrossRef] [PubMed]
- Burdziej, A.; Paczkowski, C.; Destrac-Irvine, A.; Richard, T.; Cluzet, S.; Szakiel, A. Triterpenoid profiles of the leaves of wild and domesticated grapevines. Phytochem. Lett. 2019, 30, 302–308. [Google Scholar] [CrossRef]
- Hely, P.C.; Pasfield, G.; Gellatley, J.G. Insect Pests of Fruit and Vegetables in New South Wales; Inkarta Press: Melbourne, Australia; London, UK, 1982; p. 312. [Google Scholar]
- Rakimov, A. Aspects of the Biology, Ecology and Biological Control of Soft Scale Insects (Coccidae) in Australian Vineyards; The University of Melbourne: Melbourne, Australia, 2010. [Google Scholar]
- Buchanan, G. Biological control of grapevine scales. Department of Primary Industries; Biosciences Research Division; DPI: Mildura, Australia, 2008; p. 32. [Google Scholar]
- Simbiken, N.A. Feeding and Ecology of Grapevine Scales, Parthenolecanium persicae and P. pruinosum, on Different Varieties of Grapevine. Ph.D. Thesis, The Australian National University, Canberra, Australia, 2014. [Google Scholar]
- Saleh, M.; Salama, H.S. Main chemical components of the honey dew excreted by the vine mealy bug Planococcus vitis. J. Insect Phys. 1971, 17, 1661–1663. [Google Scholar] [CrossRef]
- Venus, J. Scale in Vineyards-Identification and Control; South Australia Central, Wine Australia Regional Program: Adelaide, South Australia, Australia, 2017; p. 27. Available online: https://www.wineaustralia.com/getmedia/ac2d8790-432e-4f3d-9824-d509b9e903a0/SA-Central-2016_17-AOP-Activity-2-Scale-in-Vineyards-Final-REPORT (accessed on 20 November 2022).
- Cooper, P.D.; Venus, J. Climate change and its influence on scale insects and sooty mould occurrence. In Proceedings of the 17th Australian Wine Industry Technical Conference, Adelaide, Australia, 21–24 July 2019; Beames, K.S., Robinson, E.M.C., Dry, P.R., Johnson, D.L., Eds.; The Australian Wine Industry Technical Conference Inc.: Urrbrae, Australia, 2020; pp. 40–44. [Google Scholar]
- Simbiken, N.A.; Cooper, P.D.; Powell, K.S. Development and feeding effect of frosted scale Parthenolecanium pruinosum Cocquillet (Hemiptera: Coccidae) on selected Vitis vinifera L. cultivars. Aust. J. Grape Wine Res. 2015, 21, 451–457. [Google Scholar] [CrossRef]
- Garcia, L.C.; Eubanks, M.D. Overcompensation for insect herbivory: A review and meta-analysis of the evidence. Ecology 2019, 100, e02585. [Google Scholar] [CrossRef]
- Schaeffer, R.N.; Wang, Z.; Thornber, C.S.; Preisser, E.L.; Orians, C.M. Two invasive herbivores on a shared host: Patterns and consequences of phytohormone induction. Oecologia 2018, 186, 973–982. [Google Scholar] [CrossRef]
- Zhou, S.Q.; Jander, G. Molecular ecology of plant volatiles in interactions with insect herbivores. J. Exp. Bot. 2022, 73, 449–462. [Google Scholar] [CrossRef]
- Kwon, J.K.; Khoshimkhujaev, B.; Lee, J.H.; Yu, I.H.; Park, K.S.; Choi, H.G. Growth and Yield of Tomato and Cucumber Plants in Polycarbonate or Glass Greenhouses. Hortic. Sci. Technol. 2017, 35, 79–87. [Google Scholar] [CrossRef]
- Fernandes de Oliveira, A.; Nieddu, G. Vine growth and physiological performance of two red grape cultivars under natural and reduced UV solar radiation. Aust. J. Grape Wine Res. 2015, 22, 105–114. [Google Scholar] [CrossRef]
- Cooper, P.D. Scale Insects and the Vitas Vinifera cv. Shiraz-Reason for High Incidence of Scales; The Australian National University: Wine, Australia, 2017; Available online: https://www.wineaustralia.com/research/projects/scale-insects-and-the-vitis-vinifera-cv (accessed on 20 November 2022).
- Simbiken, N.A.; Powell, K.S.; Cooper, P.D. Scale back on scale in the vineyard. Aust. NZ Grapegrow. Winemak. 2017, 645, 42–47. [Google Scholar]
- Greathead, D.J. Crawler behaviour and dispersal. In Soft Scale Insects: Their Biology, Natural Enemies and Control; Ben-Dov, Y., Hodgson, C.J., Eds.; Elseveir: Amsterdam, The Netherlands, 1997; Volume B, pp. 339–342. [Google Scholar]
- Murphy, L. Scale Infestation of Grapevine Vitis vinifera Cultivars; Honours; The Australian National University: Canberra, Australia, 2021. [Google Scholar]
- Ding, P.T.; Ding, Y.L. Stories of salicylic acid: A plant defense hormone. Trends Plant Sci. 2020, 25, 549–565. [Google Scholar] [CrossRef]
- Perestrelo, R.; Caldeira, M.; Camara, J.S. Solid phase microextraction as a reliable alternative to conventional extraction techniques to evaluate the pattern of hydrolytically released components in Vitis vinifera L. grapes. Talanta 2012, 95, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Wirth, J.; Guo, W.F.; Baumes, R.; Gunata, Z. Volatile compounds released by enzymatic hydrolysis of glycoconjugates of leaves and grape berries from Vitis vinifera Muscat of Alexandria and Shiraz cultivars. J. Agricul. Food Chem. 2001, 49, 2917–2923. [Google Scholar] [CrossRef] [PubMed]
- Gallon, C.; Martello, R.H.; Cozzer, G.; Rezende, C.A.L.; Calisto, J.F.F.; Floss, P.A.; Oliveira, J.V.; Rezende, R.D.; Dal Magro, J.; Albeny-Simoes, D. Chemistry matters: Biological activity of Eucalyptus essential oils on mosquito larval mortality. Entomol. Exp. Appl. 2020, 168, 407–415. [Google Scholar] [CrossRef]
- Kiran, S.; Prakash, B. Toxicity and biochemical efficacy of chemically characterized Rosmarinus officinalis essential oil against Sitophilus oryzae and Oryzaephilus surinamensis. Indust. Crops Prod. 2015, 74, 817–823. [Google Scholar] [CrossRef]
- Goncalves, G.B.; Silva, C.E.; Mendonca, A.D.; Vanickova, L.; Tomcala, A.; Do Nascimento, R.R. Pheromone communication in Anastrepha obliqua (Diptera: Tephritidae): A comparison of the volatiles and salivary gland extracts of two wild populations. Flor. Entomol. 2013, 96, 1365–1374. [Google Scholar] [CrossRef]
- Mansson, P.E.; Schlyter, F.; Eriksson, C.; Sjodin, K. Nonanoic acid, other alkanoic acids, and related compounds as antifeedants in Hylobius abietis pine weevils. Entomol. Exp. Appl. 2006, 121, 191–201. [Google Scholar] [CrossRef]
- Williams, R.N.; Fickle, D.S.; McGovern, T.P.; Klein, M.G. Development of an attractant for the scarab pest Macrodactylus subspinosus (Coleoptera:Scarabaeidae). J. Econ. Entomol. 2000, 93, 1480–1484. [Google Scholar] [CrossRef]
- Timm, A.E.; Reineke, A. First insights into grapevine transcriptional responses as a result of vine mealybug Planococcus ficus feeding. Arth Plant Inter. 2014, 8, 495–505. [Google Scholar] [CrossRef]
- Naegele, R.P.; Cousins, P.; Daane, K.M. Identification of Vitis cultivars, rootstocks, and species expressing resistance to a Planococcus mealybug. Insects 2020, 11, 86. [Google Scholar] [CrossRef] [Green Version]
- Lenz, M.S.; Isaacs, R.; Flore, J.A.; Howell, G.S. Vegetative growth responses of Pinot gris (Vitis vinifera L.) grapevines to infestation by potato leafhoppers (Empoasca fabae Harris). Amer. J. Enol. Viticult. 2009, 60, 130–137. [Google Scholar] [CrossRef]
- Rivers, J.Y.; Truong, T.T.; Pogson, B.J.; McQuinn, R.P. Volatile apocarotenoid discovery and quantification in Arabidopsis thaliana: Optimized sensitive analysis via HS-SPME-GC/MS. Metabolomics 2019, 15, 79. [Google Scholar] [CrossRef] [PubMed]
- Fiehn, O.; Robertson, D.; Griffin, J.; van der Werf, M.; Nikolau, B.; Morrison, N.; Sumner, L.W.; Goodacre, R.; Hardy, N.W.; Taylor, C.; et al. The metabolomics standards initiative (MSI). Metabolomics 2007, 3, 175–178. [Google Scholar] [CrossRef] [Green Version]
- Schrimpe-Rutledge, A.C.; Codreanu, S.G.; Sherrod, S.D.; McLean, J.A. Untargeted Metabolomics Strategies-Challenges and Emerging Directions. J. Am. Soc. Mass Spectrom. 2016, 27, 1897–1905. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Chardonnay | Sauvignon Blanc | |
|---|---|---|
| Control | 60.6 ± 23.7 a | 31.2 ± 23.7 a |
| Infested | 143.4 ± 23.7 b | 114.0 ± 23.7 b |
| Chardonnay | Sauvignon Blanc | |||
|---|---|---|---|---|
| Number with Grapes | Mean Mass of Grapes (g) | Number with Grapes | Mean Mass of Grapes (g) | |
| Control | 5 | 67 ± 15.6 a | 3 | 23 ± 24.7 b |
| Infested | 1 | 112 ± 32.7 c | 6 | 68 ± 14.5 d |
| Chardonnay | Sauvignon Blanc | |||||
|---|---|---|---|---|---|---|
| Root (g) | Shoot (g) | Shoot:Root | Root (g) | Shoot (g) | Shoot:Root | |
| Control | 170 ± 32.4 a | 39 ± 2.8 c | 0.3 ± 0.06 f | 160 ± 32.4 a | 42 ± 2.8 c | 0.3 ± 0.06 f |
| Infested | 151 ± 32.2 a | 35 ± 2.8 c | 0.3 ± 0.06 f | 216 ± 32.2 b | 51 ± 2.8 d | 0.2 ± 0.06 f |
| Cultivar/Volatile | Fold Change | log2 Fold Change |
|---|---|---|
| Sauvignon Blanc | ||
| Docosane | 0.233 | −2.10 |
| Heneicosane | 0.365 | −1.45 |
| Methyl salicylate | 0.369 | −1.44 |
| Tricosane | 0.416 | −1.27 |
| Benzyl alcohol | 0.490 | −1.03 |
| Nonanoic acid | 0.554 | −0.85 |
| Phytol | 0.584 | −0.78 |
| Chardonnay | ||
| Docosane | 0.218 | −2.20 |
| Tricosane | 0.319 | −1.65 |
| Heneicosane | 0.381 | −1.39 |
| Eicosane | 0.594 | −0.75 |
| Spiro4.5decan-7-one, 1,8-dimethyl-8,9-epoxy-4-isopropyl- | 0.606 | −0.72 |
| Nonanoic acid | 0.642 | −0.64 |
| Methyl salicylate | 1.527 | 0.61 |
| 3-Oxo-α-ionol | 0.655 | −0.61 |
| 2,5-Furandione, 3,4-dimethyl- | 0.663 | −0.59 |
| 2-Octenal, E- | 1.524 | 0.61 |
| 1-Hexanol, 2-ethyl- | 1.501 | 0.59 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cooper, P.D.; Truong, T.T.; Keszei, A.; Neeman, T.; Webster, K.W. The Effect of Scale Insects on Growth Parameters of cv. Chardonnay and cv. Sauvignon Blanc Grapevines Grown in a Greenhouse. Int. J. Mol. Sci. 2023, 24, 1544. https://doi.org/10.3390/ijms24021544
Cooper PD, Truong TT, Keszei A, Neeman T, Webster KW. The Effect of Scale Insects on Growth Parameters of cv. Chardonnay and cv. Sauvignon Blanc Grapevines Grown in a Greenhouse. International Journal of Molecular Sciences. 2023; 24(2):1544. https://doi.org/10.3390/ijms24021544
Chicago/Turabian StyleCooper, Paul D., Thy T. Truong, Andras Keszei, Teresa Neeman, and Kenneth W. Webster. 2023. "The Effect of Scale Insects on Growth Parameters of cv. Chardonnay and cv. Sauvignon Blanc Grapevines Grown in a Greenhouse" International Journal of Molecular Sciences 24, no. 2: 1544. https://doi.org/10.3390/ijms24021544