
Interplay between Inter-Subunit Rotation of the Ribosome and Binding of Translational GTPases


Abstract
:1. Introduction
2. Results
2.1. LepA Stablizes the Non-Rotated Conformation of the Ribosome
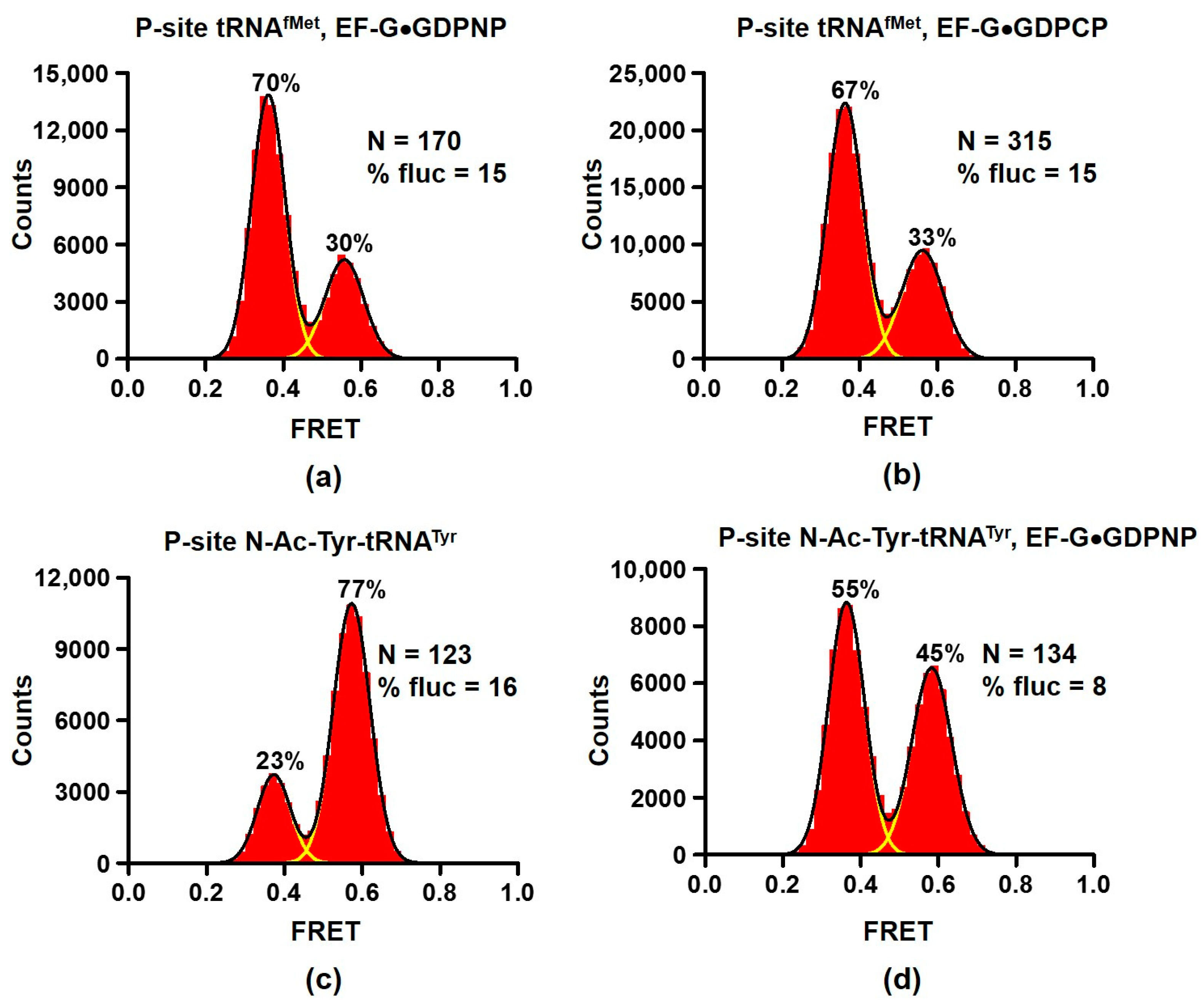
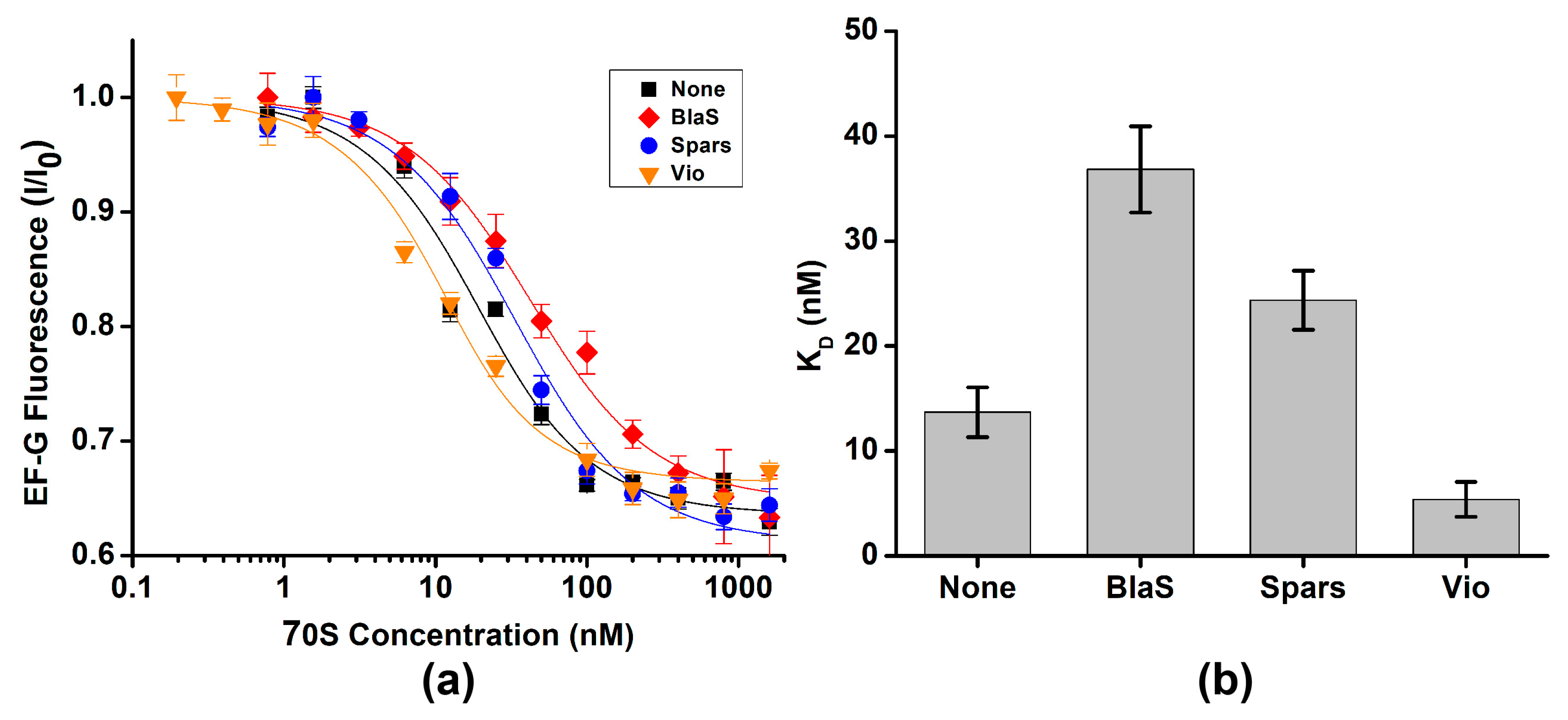
2.2. EF-G interacts with Both the Rotated and Non-Rotated Conformations of the Ribosome
3. Discussion
4. Materials and Methods
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Korostelev, A.A. The Structural Dynamics of Translation. Annu. Rev. Biochem. 2022, 91, 245–267. [Google Scholar] [CrossRef] [PubMed]
- Frank, J.; Agrawal, R.K. A ratchet-like inter-subunit reorganization of the ribosome during translocation. Nature 2000, 406, 318–322. [Google Scholar] [CrossRef] [PubMed]
- Moazed, D.; Noller, H.F. Intermediate states in the movement of transfer RNA in the ribosome. Nature 1989, 342, 142–148. [Google Scholar] [CrossRef] [PubMed]
- Ling, C.; Ermolenko, D.N. Structural insights into ribosome translocation. Wiley Interdiscip. Rev. RNA 2016, 7, 620–636. [Google Scholar] [CrossRef]
- Frank, J.; Gonzalez, R.L., Jr. Structure and dynamics of a processive Brownian motor: The translating ribosome. Annu. Rev. Biochem. 2010, 79, 381–412. [Google Scholar] [CrossRef] [Green Version]
- Cornish, P.V.; Ermolenko, D.N.; Noller, H.F.; Ha, T. Spontaneous intersubunit rotation in single ribosomes. Mol. Cell 2008, 30, 578–588. [Google Scholar] [CrossRef] [Green Version]
- Marshall, R.A.; Dorywalska, M.; Puglisi, J.D. Irreversible chemical steps control intersubunit dynamics during translation. Proc. Natl. Acad. Sci. USA 2008, 105, 15364–15369. [Google Scholar] [CrossRef] [Green Version]
- Sternberg, S.H.; Fei, J.; Prywes, N.; McGrath, K.A.; Gonzalez, R.L., Jr. Translation factors direct intrinsic ribosome dynamics during translation termination and ribosome recycling. Nat. Struct. Mol. Biol. 2009, 16, 861–868. [Google Scholar] [CrossRef] [Green Version]
- Kaledhonkar, S.; Fu, Z.; Caban, K.; Li, W.; Chen, B.; Sun, M.; Gonzalez, R.L., Jr.; Frank, J. Late steps in bacterial translation initiation visualized using time-resolved cryo-EM. Nature 2019, 570, 400–404. [Google Scholar] [CrossRef]
- Dunkle, J.A.; Wang, L.; Feldman, M.B.; Pulk, A.; Chen, V.B.; Kapral, G.J.; Noeske, J.; Richardson, J.S.; Blanchard, S.C.; Cate, J.H. Structures of the bacterial ribosome in classical and hybrid states of tRNA binding. Science 2011, 332, 981–984. [Google Scholar] [CrossRef]
- Ling, C.; Ermolenko, D.N. Initiation factor 2 stabilizes the ribosome in a semirotated conformation. Proc. Natl. Acad. Sci. USA 2015, 112, 15874–15879. [Google Scholar] [CrossRef] [Green Version]
- Adio, S.; Sharma, H.; Senyushkina, T.; Karki, P.; Maracci, C.; Wohlgemuth, I.; Holtkamp, W.; Peske, F.; Rodnina, M.V. Dynamics of ribosomes and release factors during translation termination in E. coli. Elife 2018, 7, e34252. [Google Scholar] [CrossRef]
- Shoji, S.; Janssen, B.D.; Hayes, C.S.; Fredrick, K. Translation factor LepA contributes to tellurite resistance in Escherichia coli but plays no apparent role in the fidelity of protein synthesis. Biochimie 2010, 92, 157–163. [Google Scholar] [CrossRef] [Green Version]
- Dibb, N.J.; Wolfe, P.B. lep operon proximal gene is not required for growth or secretion by Escherichia coli. J. Bacteriol. 1986, 166, 83–87. [Google Scholar] [CrossRef] [Green Version]
- Bauerschmitt, H.; Funes, S.; Herrmann, J.M. The membrane-bound GTPase Guf1 promotes mitochondrial protein synthesis under suboptimal conditions. J. Biol. Chem. 2008, 283, 17139–17146. [Google Scholar] [CrossRef] [Green Version]
- Gao, Y.; Bai, X.; Zhang, D.; Han, C.; Yuan, J.; Liu, W.; Cao, X.; Chen, Z.; Shangguan, F.; Zhu, Z.; et al. Mammalian elongation factor 4 regulates mitochondrial translation essential for spermatogenesis. Nat. Struct. Mol. Biol. 2016, 23, 441–449. [Google Scholar] [CrossRef]
- Qin, Y.; Polacek, N.; Vesper, O.; Staub, E.; Einfeldt, E.; Wilson, D.N.; Nierhaus, K.H. The highly conserved LepA is a ribosomal elongation factor that back-translocates the ribosome. Cell 2006, 127, 721–733. [Google Scholar] [CrossRef] [Green Version]
- Gibbs, M.R.; Moon, K.M.; Chen, M.; Balakrishnan, R.; Foster, L.J.; Fredrick, K. Conserved GTPase LepA (Elongation Factor 4) functions in biogenesis of the 30S subunit of the 70S ribosome. Proc. Natl. Acad. Sci. USA 2017, 114, 980–985. [Google Scholar] [CrossRef] [Green Version]
- March, P.E.; Inouye, M. GTP-binding membrane protein of Escherichia coli with sequence homology to initiation factor 2 and elongation factors Tu and G. Proc. Natl. Acad. Sci. USA 1985, 82, 7500–7504. [Google Scholar] [CrossRef] [Green Version]
- Fishbein, S.R.S.; Tomasi, F.G.; Wolf, I.D.; Dulberger, C.L.; Wang, A.; Keshishian, H.; Wallace, L.; Carr, S.A.; Ioerger, T.R.; Rego, E.H.; et al. The conserved translation factor LepA is required for optimal synthesis of a porin family in Mycobacterium smegmatis. J. Bacteriol. 2020, 203, 13370–13383. [Google Scholar] [CrossRef]
- Gagnon, M.G.; Lin, J.; Bulkley, D.; Steitz, T.A. Crystal structure of elongation factor 4 bound to a clockwise ratcheted ribosome. Science 2014, 345, 684–687. [Google Scholar] [CrossRef] [PubMed]
- Zhang, D.; Yan, K.; Liu, G.; Song, G.; Luo, J.; Shi, Y.; Cheng, E.; Wu, S.; Jiang, T.; Lou, J.; et al. EF4 disengages the peptidyl-tRNA CCA end and facilitates back-translocation on the 70S ribosome. Nat. Struct. Mol. Biol. 2016, 23, 125–131. [Google Scholar] [CrossRef] [PubMed]
- Connell, S.R.; Topf, M.; Qin, Y.; Wilson, D.N.; Mielke, T.; Fucini, P.; Nierhaus, K.H.; Spahn, C.M. A new tRNA intermediate revealed on the ribosome during EF4-mediated back-translocation. Nat. Struct. Mol. Biol. 2008, 15, 910–915. [Google Scholar] [CrossRef] [PubMed]
- Ermolenko, D.N.; Majumdar, Z.K.; Hickerson, R.P.; Spiegel, P.C.; Clegg, R.M.; Noller, H.F. Observation of intersubunit movement of the ribosome in solution using FRET. J. Mol. Biol. 2007, 370, 530–540. [Google Scholar] [CrossRef] [PubMed]
- Fei, J.; Richard, A.C.; Bronson, J.E.; Gonzalez, R.L., Jr. Transfer RNA-mediated regulation of ribosome dynamics during protein synthesis. Nat. Struct. Mol. Biol. 2011, 18, 1043–1051. [Google Scholar] [CrossRef]
- Prabhakar, A.; Capece, M.C.; Petrov, A.; Choi, J.; Puglisi, J.D. Post-termination Ribosome Intermediate Acts as the Gateway to Ribosome Recycling. Cell Rep. 2017, 20, 161–172. [Google Scholar] [CrossRef] [Green Version]
- Valle, M.; Zavialov, A.; Sengupta, J.; Rawat, U.; Ehrenberg, M.; Frank, J. Locking and unlocking of ribosomal motions. Cell 2003, 114, 123–134. [Google Scholar] [CrossRef] [Green Version]
- Zavialov, A.V.; Ehrenberg, M. Peptidyl-tRNA regulates the GTPase activity of translation factors. Cell 2003, 114, 113–122. [Google Scholar] [CrossRef] [Green Version]
- Chen, J.; Petrov, A.; Tsai, A.; O’Leary, S.E.; Puglisi, J.D. Coordinated conformational and compositional dynamics drive ribosome translocation. Nat. Struct. Mol. Biol. 2013, 20, 718–727. [Google Scholar] [CrossRef] [Green Version]
- Munro, J.B.; Wasserman, M.R.; Altman, R.B.; Wang, L.; Blanchard, S.C. Correlated conformational events in EF-G and the ribosome regulate translocation. Nat. Struct. Mol. Biol. 2010, 17, 1470–1477. [Google Scholar] [CrossRef] [Green Version]
- Chen, C.; Stevens, B.; Kaur, J.; Cabral, D.; Liu, H.; Wang, Y.; Zhang, H.; Rosenblum, G.; Smilansky, Z.; Goldman, Y.E.; et al. Single-molecule fluorescence measurements of ribosomal translocation dynamics. Mol. Cell 2011, 42, 367–377. [Google Scholar] [CrossRef] [Green Version]
- Ermolenko, D.N.; Noller, H.F. mRNA translocation occurs during the second step of ribosomal intersubunit rotation. Nat. Struct. Mol. Biol. 2011, 18, 457–462. [Google Scholar] [CrossRef] [Green Version]
- Holtkamp, W.; Cunha, C.E.; Peske, F.; Konevega, A.L.; Wintermeyer, W.; Rodnina, M.V. GTP hydrolysis by EF-G synchronizes tRNA movement on small and large ribosomal subunits. EMBO J. 2014, 33, 1073–1085. [Google Scholar] [CrossRef]
- Walker, S.E.; Shoji, S.; Pan, D.; Cooperman, B.S.; Fredrick, K. Role of hybrid tRNA-binding states in ribosomal translocation. Proc. Natl. Acad. Sci. USA 2008, 105, 9192–9197. [Google Scholar] [CrossRef] [Green Version]
- Holtkamp, W.; Wintermeyer, W.; Rodnina, M.V. Synchronous tRNA movements during translocation on the ribosome are orchestrated by elongation factor G and GTP hydrolysis. Bioessays 2014, 36, 908–918. [Google Scholar] [CrossRef] [Green Version]
- Joseph, S.; Noller, H.F. EF-G-catalyzed translocation of anticodon stem-loop analogs of transfer RNA in the ribosome. EMBO J. 1998, 17, 3478–3483. [Google Scholar] [CrossRef]
- Lancaster, L.; Lambert, N.J.; Maklan, E.J.; Horan, L.H.; Noller, H.F. The sarcin-ricin loop of 23S rRNA is essential for assembly of the functional core of the 50S ribosomal subunit. RNA 2008, 14, 1999–2012. [Google Scholar] [CrossRef] [Green Version]
- Ermolenko, D.N.; Cornish, P.V.; Ha, T.; Noller, H.F. Antibiotics that bind to the A site of the large ribosomal subunit can induce mRNA translocation. RNA 2013, 19, 158–166. [Google Scholar] [CrossRef] [Green Version]
- Svidritskiy, E.; Ling, C.; Ermolenko, D.N.; Korostelev, A.A. Blasticidin S inhibits translation by trapping deformed tRNA on the ribosome. Proc. Natl. Acad. Sci. USA 2013, 110, 12283–12288. [Google Scholar] [CrossRef] [Green Version]
- Ermolenko, D.N.; Spiegel, P.C.; Majumdar, Z.K.; Hickerson, R.P.; Clegg, R.M.; Noller, H.F. The antibiotic viomycin traps the ribosome in an intermediate state of translocation. Nat. Struct. Mol. Biol. 2007, 14, 493–497. [Google Scholar] [CrossRef]
- Modolell, J.; Vazquez. The inhibition of ribosomal translocation by viomycin. Eur. J. Biochem. 1977, 81, 491–497. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Herrmann, C.J.; Simonovic, M.; Szklarczyk, D.; von Mering, C. Version 4.0 of PaxDb: Protein abundance data, integrated across model organisms, tissues, and cell-lines. Proteomics 2015, 15, 3163–3168. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brilot, A.F.; Korostelev, A.A.; Ermolenko, D.N.; Grigorieff, N. Structure of the ribosome with elongation factor G trapped in the pretranslocation state. Proc. Natl. Acad. Sci. USA 2013, 110, 20994–20999. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gao, Y.G.; Selmer, M.; Dunham, C.M.; Weixlbaumer, A.; Kelley, A.C.; Ramakrishnan, V. The structure of the ribosome with elongation factor G trapped in the posttranslocational state. Science 2009, 326, 694–699. [Google Scholar] [CrossRef] [Green Version]
- Ratje, A.H.; Loerke, J.; Mikolajka, A.; Brunner, M.; Hildebrand, P.W.; Starosta, A.L.; Donhofer, A.; Connell, S.R.; Fucini, P.; Mielke, T.; et al. Head swivel on the ribosome facilitates translocation by means of intra-subunit tRNA hybrid sites. Nature 2010, 468, 713–716. [Google Scholar] [CrossRef] [Green Version]
- Carbone, C.E.; Loveland, A.B.; Gamper, H.B., Jr.; Hou, Y.M.; Demo, G.; Korostelev, A.A. Time-resolved cryo-EM visualizes ribosomal translocation with EF-G and GTP. Nat. Commun. 2021, 12, 7236. [Google Scholar] [CrossRef]
- Kumar, V.; Ero, R.; Ahmed, T.; Goh, K.J.; Zhan, Y.; Bhushan, S.; Gao, Y.G. Structure of the GTP Form of Elongation Factor 4 (EF4) Bound to the Ribosome. J. Biol. Chem. 2016, 291, 12943–12950. [Google Scholar] [CrossRef] [Green Version]
- Pohlschroder, M.; Hartmann, E.; Hand, N.J.; Dilks, K.; Haddad, A. Diversity and evolution of protein translocation. Annu. Rev. Microbiol. 2005, 59, 91–111. [Google Scholar] [CrossRef]
- Itoh, Y.; Andrell, J.; Choi, A.; Richter, U.; Maiti, P.; Best, R.B.; Barrientos, A.; Battersby, B.J.; Amunts, A. Mechanism of membrane-tethered mitochondrial protein synthesis. Science 2021, 371, 846–849. [Google Scholar] [CrossRef]
- Rexroad, G.; Donohue, J.P.; Lancaster, L.; Noller, H.F. The role of GTP hydrolysis by EF-G in ribosomal translocation. Proc. Natl. Acad. Sci. USA 2022, 119, e2212502119. [Google Scholar] [CrossRef]
- Salsi, E.; Farah, E.; Dann, J.; Ermolenko, D.N. Following movement of domain IV of elongation factor G during ribosomal translocation. Proc. Natl. Acad. Sci. USA 2014, 111, 15060–15065. [Google Scholar] [CrossRef] [Green Version]
- Walter, J.D.; Hunter, M.; Cobb, M.; Traeger, G.; Spiegel, P.C. Thiostrepton inhibits stable 70S ribosome binding and ribosome-dependent GTPase activation of elongation factor G and elongation factor 4. Nucleic Acids Res. 2012, 40, 360–370. [Google Scholar] [CrossRef]
- Bao, C.; Loerch, S.; Ling, C.; Korostelev, A.A.; Grigorieff, N.; Ermolenko, D.N. mRNA stem-loops can pause the ribosome by hindering A-site tRNA binding. Elife 2020, 9, e55799. [Google Scholar] [CrossRef]
- Roy, S. Fluorescence quenching methods to study protein-nucleic acid interactions. Methods Enzymol. 2004, 379, 175–187. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| P Site | Antibiotic | KD, nM | p Value |
|---|---|---|---|
| Vacant | - | 115 ± 5 | |
| tRNAfMet | - | 16 ± 2 | |
| N-Ac-Met-tRNAfMet | - | 66 ± 2 | 8 × 10−7 |
| tRNAPhe | - | 14 ± 2 | |
| Blasticidin S | 37 ± 4 | 7 × 10−5 | |
| Sparsomycin | 24 ± 3 | 1 × 10−3 | |
| Viomycin | ≤5 | 4 × 10−3 | |
| N-Ac-Phe-tRNAPhe | - | 31 ± 4 | 1 × 10−3 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Das, A.; Adiletta, N.; Ermolenko, D.N. Interplay between Inter-Subunit Rotation of the Ribosome and Binding of Translational GTPases. Int. J. Mol. Sci. 2023, 24, 6878. https://doi.org/10.3390/ijms24086878
Das A, Adiletta N, Ermolenko DN. Interplay between Inter-Subunit Rotation of the Ribosome and Binding of Translational GTPases. International Journal of Molecular Sciences. 2023; 24(8):6878. https://doi.org/10.3390/ijms24086878
Chicago/Turabian StyleDas, Ananya, Nichole Adiletta, and Dmitri N. Ermolenko. 2023. "Interplay between Inter-Subunit Rotation of the Ribosome and Binding of Translational GTPases" International Journal of Molecular Sciences 24, no. 8: 6878. https://doi.org/10.3390/ijms24086878