
Identification and Functional Characterization of CsMYCs in Cucumber Glandular Trichome Development
 , , and
, , and
Abstract
:1. Introduction
2. Results
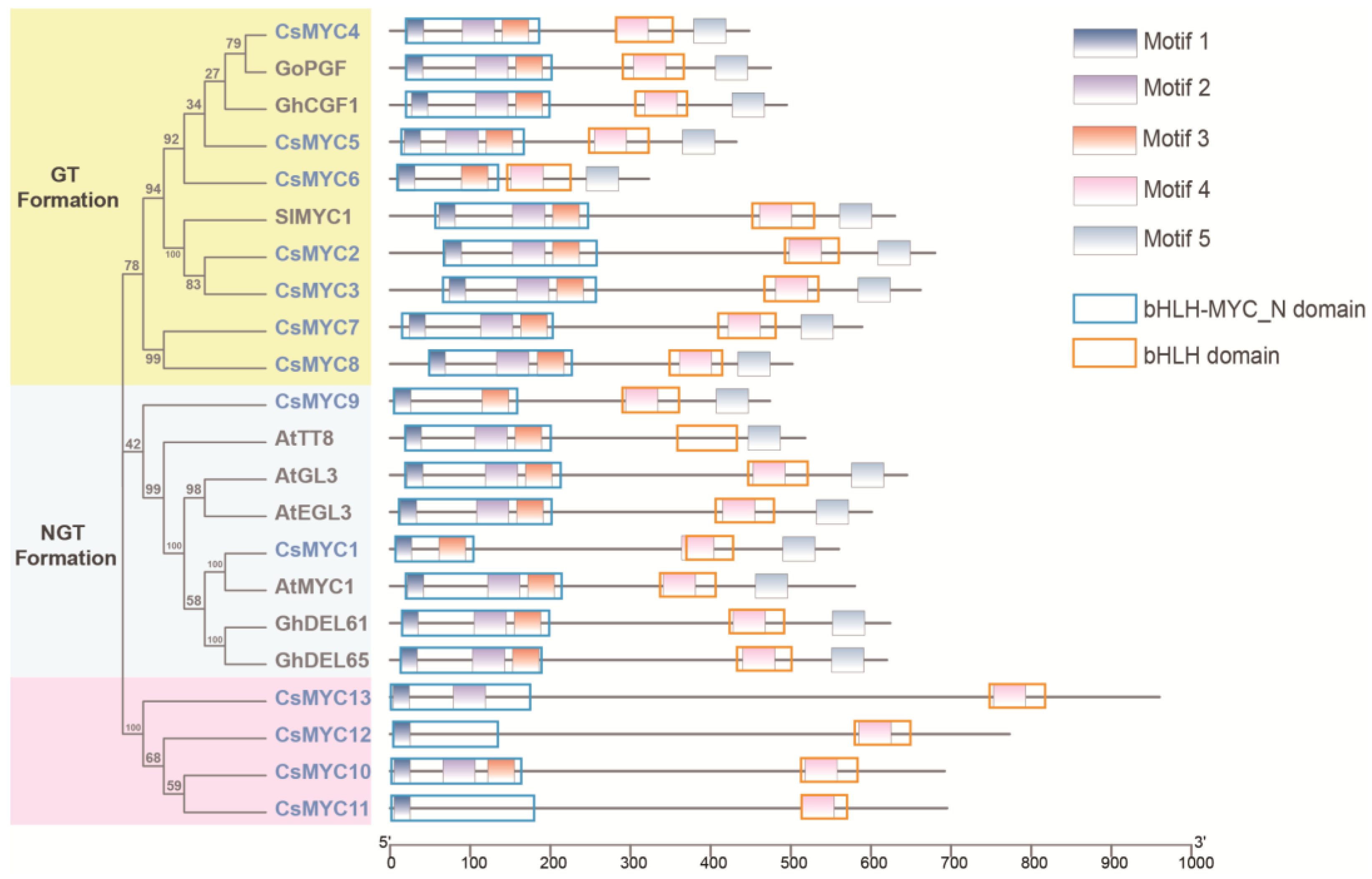
2.1. Genome-Wide Identification and Phylogenetic Analysis of Cucumber MYC Transcription Factors
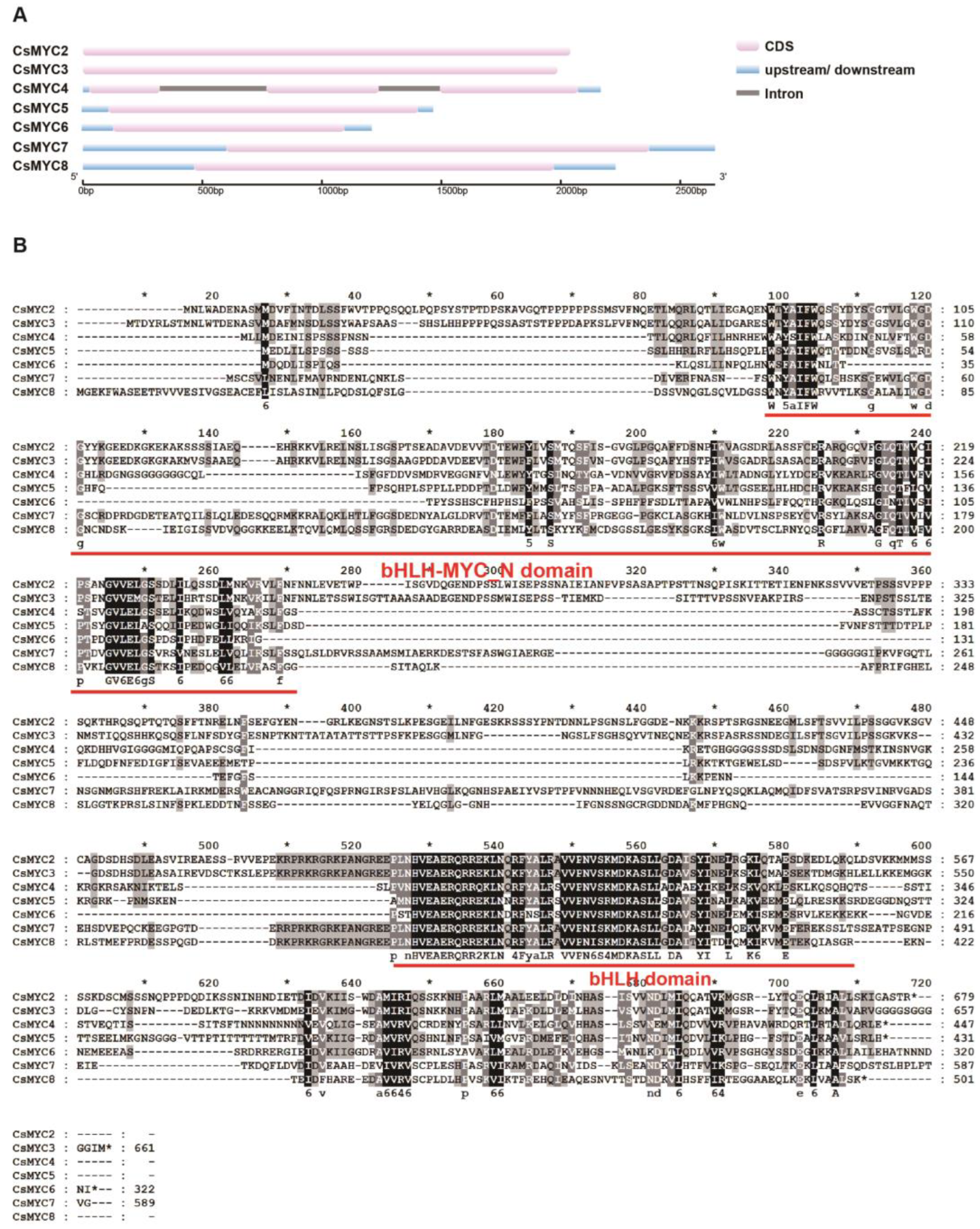
2.2. Identification of Candidate CsMYCs in Cucumber GT Development
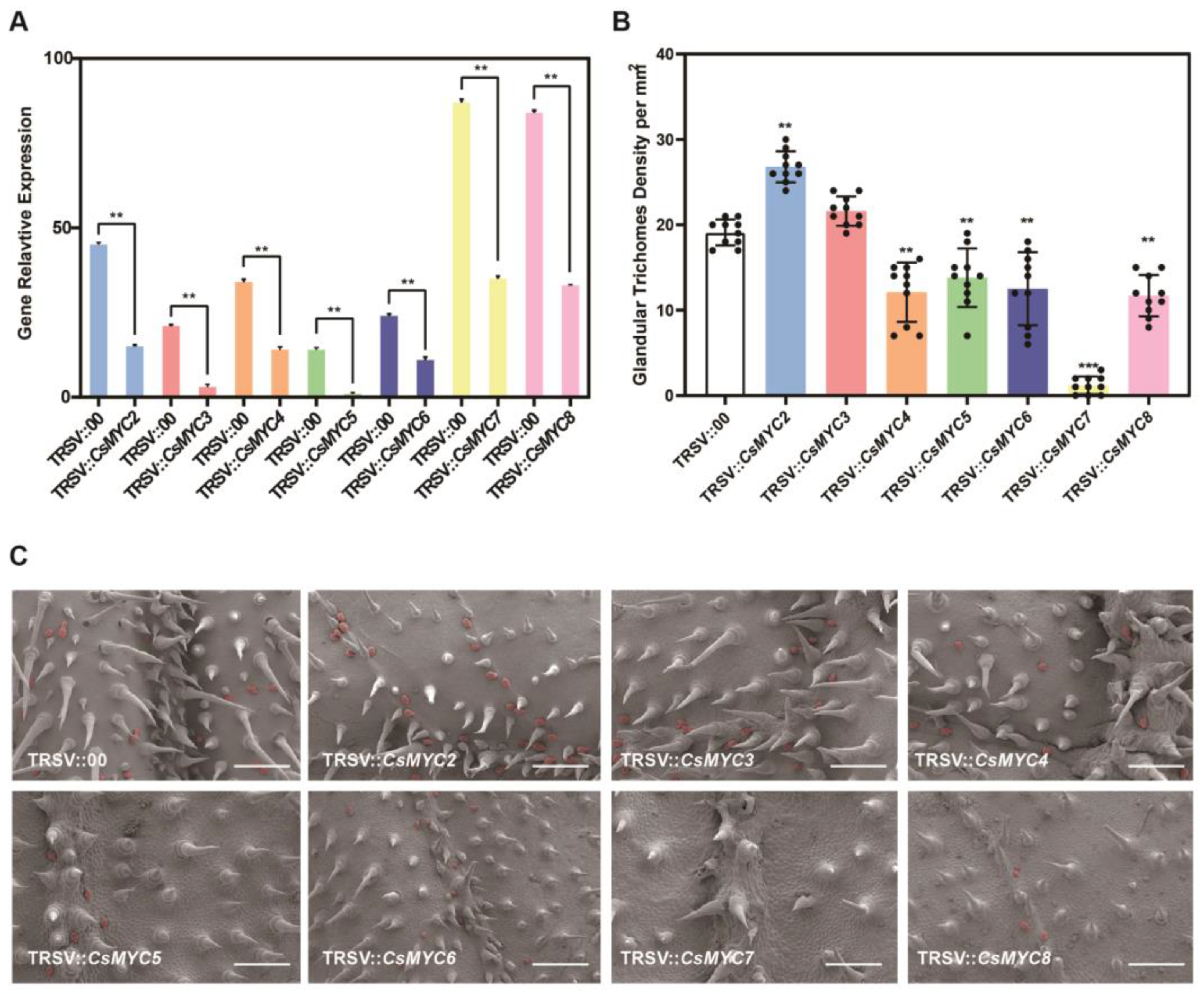
2.3. Candidate CsMYCs May Regulate the Formation of Cucumber GTs
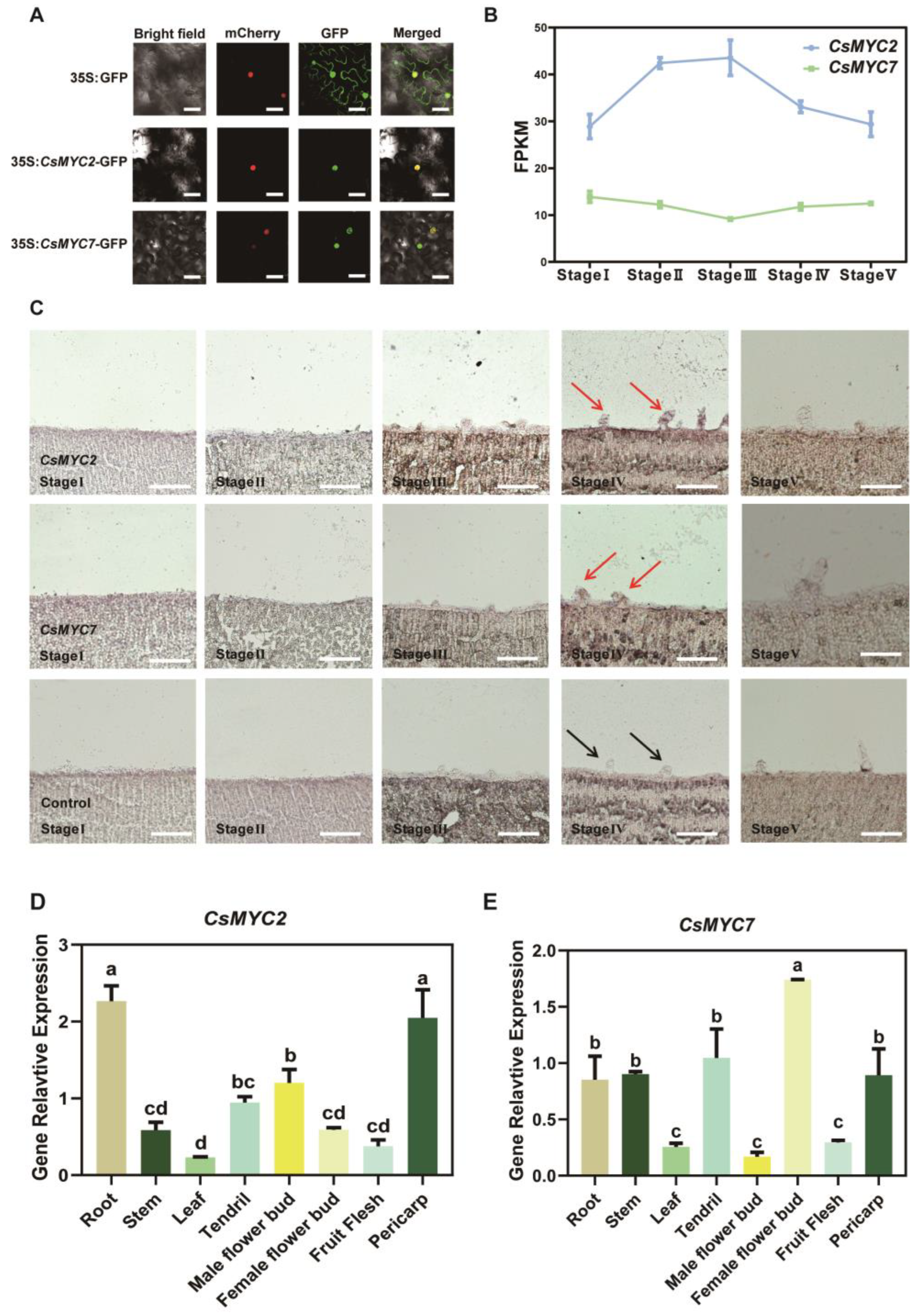
2.4. CsMYC2 and CsMYC7 Show Similar Expression Patterns in Cucumber
3. Discussion
4. Materials and Methods
4.1. Gene Identification
4.2. Phylogenetic Analysis
4.3. Gene Conserved Motif Identification
4.4. Gene Cloning
4.5. Gene Structure and Protein Conserved Domain Alignment Analysis
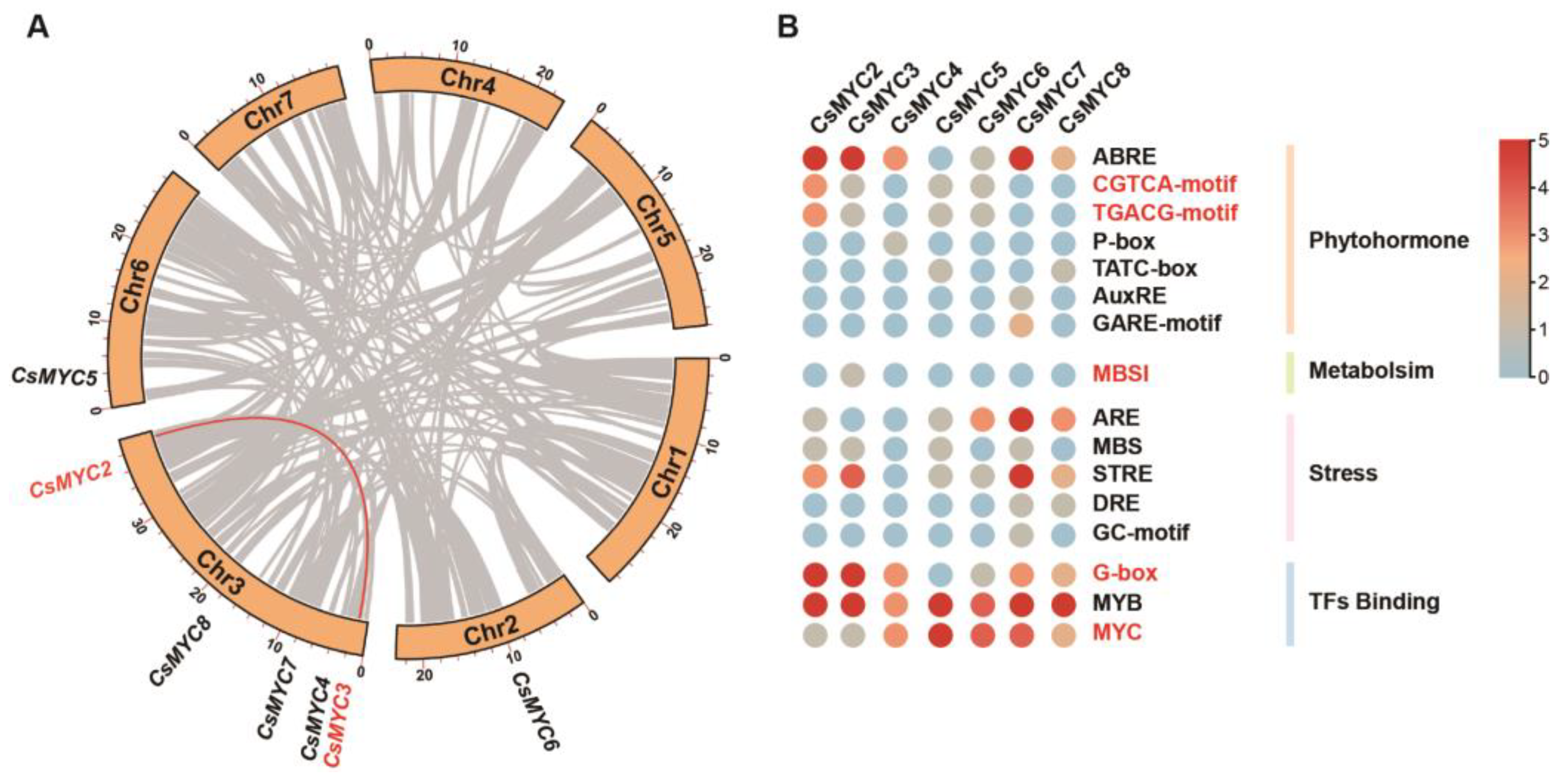
4.6. Collinearity Analysis
4.7. Cis-Regulatory Elements Analysis
4.8. Plant Materials
4.9. RNA Extraction and Real-Time Quantitative PCR (RT-qPCR) Analysis
4.10. VIGS Assay and Phenotypic Observation
4.11. Scanning Electron Microscopy (SEM)
4.12. Subcellular Localization
4.13. In Situ Hybridization
4.14. Accession Numbers
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Singh, S.P.; Upadhyay, S.K.; Pandey, A.; Kumar, S. Molecular Approaches in Plant Biology and Environmental Challenges; Springer: Singapore, 2019; ISBN 9789811506895. [Google Scholar]
- Fahn, A. Structure and Function of Secretory Cells. Adv. Bot. Res. 2000, 31, 37–75. [Google Scholar] [CrossRef]
- Huchelmann, A.; Boutry, M.; Hachez, C. Plant Glandular Trichomes: Natural Cell Factories of High Biotechnological Interest. Plant Physiol. 2017, 175, 6–22. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, C.; Srividya, N.; Parrish, A.N.; Yue, W.; Shan, M.; Wu, Q.; Lange, B.M. Morphology of Glandular Trichomes of Japanese Catnip (Schizonepeta tenuifolia Briquet) and Developmental Dynamics of Their Secretory Activity. Phytochemistry 2018, 150, 23–30. [Google Scholar] [CrossRef] [PubMed]
- Crang, R.; Lyons-Sobaski, S.; Wise, R. Secretory Structures. In Plant Anatomy; Springer International Publishing: Cham, Switzerland, 2018; pp. 443–476. ISBN 978-3-319-77208-0. [Google Scholar]
- Tanney, C.A.S.; Backer, R.; Geitmann, A.; Smith, D.L. Cannabis Glandular Trichomes: A Cellular Metabolite Factory. Front. Plant Sci. 2021, 12, 721986. [Google Scholar] [CrossRef] [PubMed]
- Chalvin, C.; Drevensek, S.; Dron, M.; Bendahmane, A.; Boualem, A. Genetic Control of Glandular Trichome Development. Trends Plant Sci. 2020, 25, 477–487. [Google Scholar] [CrossRef] [PubMed]
- Werker, E. Trichome Diversity and Development. Adv. Bot. Res. 2000, 31, 1–35. [Google Scholar] [CrossRef]
- Xue, S.; Dong, M.; Liu, X.; Xu, S.; Pang, J.; Zhang, W.; Weng, Y.; Ren, H. Classification of Fruit Trichomes in Cucumber and Effects of Plant Hormones on Type II Fruit Trichome Development. Planta 2019, 249, 407–416. [Google Scholar] [CrossRef]
- Dong, M.; Xue, S.; Bartholomew, E.S.; Zhai, X.; Sun, L.; Xu, S.; Zhang, Y.; Yin, S.; Ma, W.; Chen, S.; et al. Transcriptomic and Functional Analysis Provides Molecular Insights into Multicellular Trichome Development. Plant Physiol. 2022, 189, 301–314. [Google Scholar] [CrossRef]
- Cui, J.-Y.; Miao, H.; Ding, L.-H.; Wehner, T.C.; Liu, P.-N.; Wang, Y.; Zhang, S.-P.; Gu, X.-F. A New Glabrous Gene (Csgl3) Identified in Trichome Development in Cucumber (Cucumis sativus L.). PLoS ONE 2016, 11, e0148422. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.-L.; Nie, J.; Chen, H.-M.; Guo, C.; Pan, J.; He, H.-L.; Pan, J.-S.; Cai, R. Identification and Mapping of Tril, a Homeodomain-Leucine Zipper Gene Involved in Multicellular Trichome Initiation in Cucumis sativus. Theor. Appl. Genet. 2016, 129, 305–316. [Google Scholar] [CrossRef]
- Chen, C.; Liu, M.; Jiang, L.; Liu, X.; Zhao, J.; Yan, S.; Yang, S.; Ren, H.; Liu, R.; Zhang, X. Transcriptome Profiling Reveals Roles of Meristem Regulators and Polarity Genes during Fruit Trichome Development in Cucumber (Cucumis sativus L.). J. Exp. Bot. 2014, 65, 4943–4958. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Q.; Cao, C.; Zhang, C.; Zheng, S.; Wang, Z.; Wang, L.; Ren, Z. The Identification of Cucumis sativus Glabrous 1 (CsGL1) Required for the Formation of Trichomes Uncovers a Novel Function for the Homeodomain-Leucine Zipper I Gene. J. Exp. Bot. 2015, 66, 2515–2526. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.; Shen, J.; Bartholomew, E.S.; Dong, M.; Chen, S.; Yin, S.; Zhai, X.; Feng, Z.; Ren, H.; Liu, X. TINY BRANCHED HAIR Functions in Multicellular Trichome Development through an Ethylene Pathway in Cucumis sativus L. Plant J. 2021, 106, 753–765. [Google Scholar] [CrossRef]
- Zhao, J.-L.; Pan, J.-S.; Guan, Y.; Zhang, W.-W.; Bie, B.-B.; Wang, Y.-L.; He, H.-L.; Lian, H.-L.; Cai, R. Micro-Trichomeas a Class I Homeodomain-Leucine Zipper Gene Regulates Multicellular Trichome Development in Cucumis sativus. J. Integr. Plant Biol. 2015, 57, 925–935. [Google Scholar] [CrossRef]
- Chen, C.; Yin, S.; Liu, X.; Liu, B.; Yang, S.; Xue, S.; Cai, Y.; Black, K.; Liu, H.; Dong, M.; et al. The WD-Repeat Protein CsTTG1 Regulates Fruit Wart Formation through Interaction with the Homeodomain-Leucine Zipper I Protein Mict. Plant Physiol. 2016, 171, 1156. [Google Scholar] [CrossRef] [Green Version]
- Goossens, J.; Mertens, J.; Goossens, A. Role and Functioning of bHLH Transcription Factors in Jasmonate Signalling. J. Exp. Bot. 2016, 68, 1333–1347. [Google Scholar] [CrossRef] [PubMed]
- Sun, X.; Wang, Y.; Sui, N. Transcriptional Regulation of bHLH during Plant Response to Stress. Biochem. Biophys. Res. Commun. 2018, 503, 397–401. [Google Scholar] [CrossRef]
- Li, J.; Wang, T.; Han, J.; Ren, Z. Genome-Wide Identification and Characterization of Cucumber bHLH Family Genes and the Functional Characterization of CsbHLH041 in NaCl and ABA Tolerance in Arabidopsis and Cucumber. BMC Plant Biol. 2020, 20, 272. [Google Scholar] [CrossRef]
- Ludwig, S.R.; Habera, L.F.; Dellaporta, S.L.; Wessler, S.R. Lc, a Member of the Maize R Gene Family Responsible for Tissue-Specific Anthocyanin Production, encodes a Protein Similar to Transcriptional Activators and Contains the Myc-Homology Region. Proc. Natl. Acad. Sci. USA 1989, 86, 7092–7096. [Google Scholar] [CrossRef] [Green Version]
- Chen, S.; Zhao, H.; Luo, T.; Liu, Y.; Nie, X.; Li, H. Characteristics and Expression Pattern of MYC Genes in Triticum aestivum, Oryza sativa, and Brachypodium distachyon. Plants 2019, 8, 274. [Google Scholar] [CrossRef] [Green Version]
- Li, X.; Duan, X.; Jiang, H.; Sun, Y.; Tang, Y.; Yuan, Z.; Guo, J.; Liang, W.; Chen, L.; Yin, J.; et al. Genome-Wide Analysis of basic/Helix-Loop-Helix Transcription Factor Family in Rice and Arabidopsis. Plant Physiol. 2006, 141, 1167–1184. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fernández-Calvo, P.; Chini, A.; Fernández-Barbero, G.; Chico, J.-M.; Gimenez-Ibanez, S.; Geerinck, J.; Eeckhout, D.; Schweizer, F.; Godoy, M.; Franco-Zorrilla, J.M.; et al. The Arabidopsis bHLH Transcription Factors MYC3 and MYC4 Are Targets of JAZ Repressors and Act Additively with MYC2 in the Activation of Jasmonate Responses. Plant Cell 2011, 23, 701–715. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pires, N.; Dolan, L. Origin and Diversification of basic-Helix-Loop-Helix Proteins in Plants. Mol. Biol. Evol. 2010, 27, 862–874. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heim, M.A. The Basic Helix-Loop-Helix Transcription Factor Family in Plants: A Genome-Wide Study of Protein Structure and Functional Diversity. Mol. Biol. Evol. 2003, 20, 735–747. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fonseca, S.; Fernández-Calvo, P.; Fernández, G.M.; Díez-Díaz, M.; Gimenez-Ibanez, S.; López-Vidriero, I.; Godoy, M.; Fernández-Barbero, G.; Van Leene, J.; De Jaeger, G.; et al. bHLH003, bHLH013 and bHLH017 Are New Targets of JAZ Repressors Negatively Regulating JA Responses. PLoS ONE 2014, 9, e86182. [Google Scholar] [CrossRef] [Green Version]
- Huang, H.; Gao, H.; Liu, B.; Fan, M.; Wang, J.; Wang, C.; Tian, H.; Wang, L.; Xie, C.; Wu, D.; et al. bHLH13 Regulates Jasmonate-Mediated Defense Responses and Growth. Evol. Bioinform. 2018, 14, 117693431879026. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nakata, M.; Ohme-Takagi, M. Two bHLH-Type Transcription Factors, JA-ASSOCIATED MYC2-LIKE2 and JAM3, Are Transcriptional Repressors and Affect Male Fertility. Plant Signal. Behav. 2013, 8, e26473. [Google Scholar] [CrossRef] [Green Version]
- Yadav, V.; Mallappa, C.; Gangappa, S.N.; Bhatia, S.; Chattopadhyay, S. A Basic Helix-Loop-Helix Transcription Factor in Arabidopsis, MYC2, Acts as a Repressor of Blue Light–Mediated Photomorphogenic Growth. Plant Cell 2005, 17, 1953–1966. [Google Scholar] [CrossRef] [Green Version]
- Sasaki-Sekimoto, Y.; Saito, H.; Masuda, S.; Shirasu, K.; Ohta, H. Comprehensive Analysis of Protein Interactions between JAZ Proteins and bHLH Transcription Factors That Negatively Regulate Jasmonate Signaling. Plant Signal. Behav. 2014, 9, e27639. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Du, M.; Deng, L.; Shen, J.; Fang, M.; Chen, Q.; Lu, Y.; Wang, Q.; Li, C.; Zhai, Q. MYC2 Regulates the Termination of Jasmonate Signaling via an Autoregulatory Negative Feedback Loop. Plant Cell 2019, 31, 106–127. [Google Scholar] [CrossRef] [Green Version]
- Symonds, V.V.; Hatlestad, G.; Lloyd, A.M. Natural Allelic Variation Defines a Role for ATMYC1: Trichome Cell Fate Determination. PLoS Genet. 2011, 7, e1002069. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maes, L.; Inzé, D.; Goossens, A. Functional Specialization of the TRANSPARENT TESTA GLABRA1 Network Allows Differential Hormonal Control of Laminal and Marginal Trichome Initiation in Arabidopsis Rosette Leaves. Plant Physiol. 2008, 148, 1453–1464. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Payne, C.T.; Zhang, F.; Lloyd, A.M. GL3 Encodes a bHLH Protein That Regulates Trichome Development in Arabidopsis through Interaction with GL1 and TTG1. Genetics 2000, 156, 1349–1362. [Google Scholar] [CrossRef] [PubMed]
- Zhang, F.; Gonzalez, A.; Zhao, M.; Payne, C.T.; Lloyd, A. A Network of Redundant bHLH Proteins Functions in All TTG1-Dependent Pathways of Arabidopsis. Development 2003, 130, 4859–4869. [Google Scholar] [CrossRef] [Green Version]
- Feng, Z.; Bartholomew, E.S.; Liu, Z.; Cui, Y.; Dong, Y.; Li, S.; Wu, H.; Ren, H.; Liu, X. Glandular Trichomes: New Focus on Horticultural Crops. Hortic. Res. 2021, 8, 158. [Google Scholar] [CrossRef]
- Xu, J.; van Herwijnen, Z.O.; Dräger, D.B.; Sui, C.; Haring, M.A.; Schuurink, R.C. SlMYC1 Regulates Type VI Glandular Trichome Formation and Terpene Biosynthesis in Tomato Glandular Cells. Plant Cell 2018, 30, 2988–3005. [Google Scholar] [CrossRef] [Green Version]
- Hua, B.; Chang, J.; Wu, M.; Xu, Z.; Zhang, F.; Yang, M.; Xu, H.; Wang, L.; Chen, X.; Wu, S. Mediation of JA Signalling in Glandular Trichomes by the Woolly/SlMYC1 Regulatory Module Improves Pest Resistance in Tomato. Plant Biotechnol. J. 2021, 19, 375–393. [Google Scholar] [CrossRef]
- Ma, D.; Hu, Y.; Yang, C.; Liu, B.; Fang, L.; Wan, Q.; Liang, W.; Mei, G.; Wang, L.; Wang, H.; et al. Genetic Basis for Glandular Trichome Formation in Cotton. Nat. Commun. 2016, 7, 10456. [Google Scholar] [CrossRef] [Green Version]
- Janga, M.R.; Pandeya, D.; Campbell, L.M.; Konganti, K.; Villafuerte, S.T.; Puckhaber, L.; Pepper, A.; Stipanovic, R.D.; Scheffler, J.A.; Rathore, K.S. Genes Regulating Gland Development in the Cotton Plant. Plant Biotechnol. J. 2019, 17, 1142–1153. [Google Scholar] [CrossRef]
- Kazan, K.; Manners, J.M. MYC2: The Master in Action. Mol. Plant 2013, 6, 686–703. [Google Scholar] [CrossRef] [Green Version]
- Ferguson, A.C.; Pearce, S.; Band, L.R.; Yang, C.; Ferjentsikova, I.; King, J.; Yuan, Z.; Zhang, D.; Wilson, Z.A. Biphasic Regulation of the Transcription Factor ABORTED MICROSPORES (AMS) Is Essential for Tapetum and Pollen Development in Arabidopsis. New Phytol. 2017, 213, 778–790. [Google Scholar] [CrossRef] [PubMed]
- Wendrich, J.R.; Yang, B.; Vandamme, N.; Verstaen, K.; Smet, W.; Van de Velde, C.; Minne, M.; Wybouw, B.; Mor, E.; Arents, H.E.; et al. Vascular Transcription Factors Guide Plant Epidermal Responses to Limiting Phosphate Conditions. Science 2020, 370, 1783–1786. [Google Scholar] [CrossRef] [PubMed]
- Wang, G.; Zhao, G.-H.; Jia, Y.-H.; Du, X.-M. Identification and Characterization of Cotton Genes Involved in Fuzz-Fiber Development. J. Integr. Plant Biol. 2013, 55, 619–630. [Google Scholar] [CrossRef] [PubMed]
- Shangguan, X.-X.; Yang, C.-Q.; Zhang, X.-F.; Wang, L.-J. Functional Characterization of a basic Helix-Loop-Helix (bHLH) Transcription Factor GhDEL65 from Cotton (Gossypium hirsutum). Physiol. Plant. 2016, 158, 200–212. [Google Scholar] [CrossRef] [PubMed]
- Zhang, B.; Chopra, D.; Schrader, A.; Hülskamp, M. Evolutionary Comparison of Competitive Protein-Complex Formation of MYB, bHLH, and WDR Proteins in Plants. J. Exp. Bot. 2019, 70, 3197–3209. [Google Scholar] [CrossRef] [Green Version]
- Schweizer, F.; Fernández-Calvo, P.; Zander, M.; Diez-Diaz, M.; Fonseca, S.; Glauser, G.; Lewsey, M.G.; Ecker, J.R.; Solano, R.; Reymond, P. Arabidopsis Basic Helix-Loop-Helix Transcription Factors MYC2, MYC3, and MYC4 Regulate Glucosinolate Biosynthesis, Insect Performance, and Feeding Behavior. Plant Cell 2013, 25, 3117–3132. [Google Scholar] [CrossRef] [Green Version]
- Bernhardt, C.; Lee, M.M.; Gonzalez, A.; Zhang, F.; Lloyd, A.; Schiefelbein, J. The bHLH Genes GLABRA3 (GL3) and ENHANCER OF GLABRA3 (EGL3) Specify Epidermal Cell Fate in the Arabidopsis Root. Development 2003, 130, 6431–6439. [Google Scholar] [CrossRef] [Green Version]
- Min, D.; Li, F.; Cui, X.; Zhou, J.; Li, J.; Ai, W.; Shu, P.; Zhang, X.; Li, X.; Meng, D.; et al. SlMYC2 Are Required for Methyl Jasmonate-Induced Tomato Fruit Resistance to Botrytis Cinerea. Food Chem. 2020, 310, 125901. [Google Scholar] [CrossRef]
- Yang, J.; Duan, G.; Li, C.; Liu, L.; Han, G.; Zhang, Y.; Wang, C. The Crosstalks Between Jasmonic Acid and Other Plant Hormone Signaling Highlight the Involvement of Jasmonic Acid as a Core Component in Plant Response to Biotic and Abiotic Stresses. Front. Plant Sci. 2019, 10, 1349. [Google Scholar] [CrossRef] [Green Version]
- Huang, H.; Liu, B.; Liu, L.; Song, S. Jasmonate Action in Plant Growth and Development. J. Exp. Bot. 2017, 68, 1349–1359. [Google Scholar] [CrossRef] [Green Version]
- Yan, T.; Chen, M.; Shen, Q.; Li, L.; Fu, X.; Pan, Q.; Tang, Y.; Shi, P.; Lv, Z.; Jiang, W.; et al. HOMEODOMAIN PROTEIN 1 Is Required for Jasmonate-mediated Glandular Trichome Initiation in Artemisia annua. New Phytol. 2017, 213, 1145–1155. [Google Scholar] [CrossRef] [PubMed]
- Hua, B.; Chang, J.; Xu, Z.; Han, X.; Xu, M.; Yang, M.; Yang, C.; Ye, Z.; Wu, S. HOMEODOMAIN PROTEIN 8 Mediates Jasmonate-triggered Trichome Elongation in Tomato. New Phytol. 2021, 230, 1063–1077. [Google Scholar] [CrossRef] [PubMed]
- Mistry, J.; Chuguransky, S.; Williams, L.; Qureshi, M.; Salazar, G.A.; Sonnhammer, E.L.L.; Tosatto, S.C.E.; Paladin, L.; Raj, S.; Richardson, L.J.; et al. Pfam: The Protein Families Database in 2021. Nucleic Acids Res. 2021, 49, D412–D419. [Google Scholar] [CrossRef]
- Huang, S.; Li, R.; Zhang, Z.; Li, L.; Gu, X.; Fan, W.; Lucas, W.J.; Wang, X.; Xie, B.; Ni, P.; et al. The Genome of the Cucumber, Cucumis sativus L. Nat. Genet. 2009, 41, 1275–1281. [Google Scholar] [CrossRef] [Green Version]
- Lu, S.; Wang, J.; Chitsaz, F.; Derbyshire, M.K.; Geer, R.C.; Gonzales, N.R.; Gwadz, M.; Hurwitz, D.I.; Marchler, G.H.; Song, J.S.; et al. CDD/SPARCLE: The Conserved Domain Database in 2020. Nucleic Acids Res. 2020, 48, D265–D268. [Google Scholar] [CrossRef] [Green Version]
- Letunic, I.; Khedkar, S.; Bork, P. SMART: Recent Updates, New Developments and Status in 2020. Nucleic Acids Res. 2021, 49, D458–D460. [Google Scholar] [CrossRef] [PubMed]
- Artimo, P.; Jonnalagedda, M.; Arnold, K.; Baratin, D.; Csardi, G.; de Castro, E.; Duvaud, S.; Flegel, V.; Fortier, A.; Gasteiger, E.; et al. ExPASy: SIB Bioinformatics Resource Portal. Nucleic Acids Res. 2012, 40, W597–W603. [Google Scholar] [CrossRef]
- Jones, D.T.; Taylor, W.R.; Thornton, J.M. The Rapid Generation of Mutation Data Matrices from Protein Sequences. Comput. Appl. Biosci. 1992, 8, 275–282. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis Version 7.0 for Bigger Datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [Green Version]
- Chen, C.; Chen, H.; Zhang, Y.; Thomas, H.R.; Frank, M.H.; He, Y.; Xia, R. TBtools: An Integrative Toolkit Developed for Interactive Analyses of Big Biological Data. Mol. Plant 2020, 13, 1194–1202. [Google Scholar] [CrossRef]
- Bailey, T.L.; Elkan, C. Fitting a Mixture Model by Expectation Maximization to Discover Motifs in Biopolymers. Proc. Int. Conf. Intell. Syst. Mol. Biol. 1994, 2, 28–36. [Google Scholar] [PubMed]
- Hu, B.; Jin, J.; Guo, A.-Y.; Zhang, H.; Luo, J.; Gao, G. GSDS 2.0: An Upgraded Gene Feature Visualization Server. Bioinformatics 2015, 31, 1296–1297. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Tang, H.; DeBarry, J.D.; Tan, X.; Li, J.; Wang, X.; Lee, T.-h.; Jin, H.; Marler, B.; Guo, H.; et al. MCScanX: A Toolkit for Detection and Evolutionary Analysis of Gene Synteny and Collinearity. Nucleic Acids Res. 2012, 40, e49. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lescot, M.; Déhais, P.; Thijs, G.; Marchal, K.; Moreau, Y.; Van de Peer, Y.; Rouzé, P.; Rombauts, S. PlantCARE, a Database of Plant Cis-Acting Regulatory Elements and a Portal to Tools for in Silico Analysis of Promoter Sequences. Nucleic Acids Res. 2002, 30, 325–327. [Google Scholar] [CrossRef]
- Fang, L.; Wei, X.-Y.; Liu, L.-Z.; Zhou, L.-X.; Tian, Y.-P.; Geng, C.; Li, X.-D. A Tobacco Ringspot Virus-Based Vector System for Gene and MicroRNA Function Studies in Cucurbits. Plant Physiol. 2021, 186, 853–864. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Zhou, Y.; Ding, L.; Wu, Z.; Liu, R.; Meyerowitz, E.M. Transcription Repressor HANABA TARANU Controls Flower Development by Integrating the Actions of Multiple Hormones, Floral Organ Specification Genes, and GATA3 Family Genes in Arabidopsis. Plant Cell 2013, 25, 83–101. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | Gene ID | Gene Position | CDS (bp) | Size (aa) | MW (kDa) | pI | Arabidopsis Homology | |
|---|---|---|---|---|---|---|---|---|
| Start | End (+/−) | |||||||
| CsMYC1 | Csa6G003480 | 351773 | 356445 (+) | 1680 | 559 | 62,880.54 | 5.79 | AT5G41315 |
| CsMYC2 | Csa3G902270 | 38986873 | 38988912 (+) | 2040 | 679 | 74,590.96 | 5.11 | AT1G32640 |
| CsMYC3 | Csa3G011620 | 1162977 | 1164962 (+) | 1986 | 661 | 72,140.65 | 6.03 | AT1G32640 |
| CsMYC4 | Csa3G002860 | 522560 | 524726 (−) | 1344 | 447 | 49,382.41 | 8.66 | AT4G00870 |
| CsMYC5 | Csa6G107910 | 7137128 | 7138598 (−) | 1296 | 431 | 48,394.52 | 5.42 | AT4G00870 |
| CsMYC6 | Csa2G080170 | 6627028 | 6628241 (+) | 969 | 322 | 36,381.2 | 6.02 | AT1G32640 |
| CsMYC7 | Csa3G119500 | 6736736 | 6739380 (+) | 1770 | 589 | 65,256.14 | 5.73 | AT1G01260 |
| CsMYC8 | Csa3G391380 | 19088549 | 19090777 (+) | 1560 | 519 | 55,301.14 | 5.70 | AT4G16430 |
| CsMYC9 | Csa5G601530 | 22011206 | 22013944 (+) | 1422 | 473 | 53,390.76 | 5.70 | AT2G16910 |
| CsMYC10 | Csa7G378380 | 13826607 | 13834438 (−) | 2076 | 691 | 76,190.07 | 5.66 | AT1G06150 |
| CsMYC11 | Csa3G733950 | 27943647 | 27948020 (+) | 2085 | 694 | 78,486.44 | 5.24 | AT2G27230 |
| CsMYC12 | Csa3G002970 | 600151 | 604757 (−) | 2319 | 772 | 85,076.51 | 5.11 | AT2G27230 |
| CsMYC13 | Csa1G632370 | 25233574 | 25240151 (−) | 2880 | 959 | 104,477.47 | 6.40 | AT2G27230 |
| Gene Name | Gene ID | Species | Function | References |
|---|---|---|---|---|
| GoPGF | Gh_A12G2172 | Gossypium hirsutum | GT formation | [42] |
| GhCGF1 | Gh_A11G0909 | Gossypium hirsutum | GT formation | [43] |
| SlMYC1 | Solyc08g005050 | Solanum lycopersicum | Glandular cell division and expansion | [41] |
| AtTT8 | AT4G09820 | Arabidopsis thaliana | Marginal NGT development | [36] |
| AtGL3 | AT5G41315 | Arabidopsis thaliana | NGT formation | [37] |
| AtEGL3 | AT1G63650 | Arabidopsis thaliana | NGT formation | [38] |
| AtMYC1 | AT4G00480 | Arabidopsis thaliana | NGT formation | [35] |
| GhDEL61 | LOC107904486 | Gossypium hirsutum | NGT formation | [47] |
| GhDEL65 | LOC107936704 | Gossypium hirsutum | NGT formation | [48] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Feng, Z.; Sun, L.; Dong, M.; Fan, S.; Shi, K.; Qu, Y.; Zhu, L.; Shi, J.; Wang, W.; Liu, Y.; et al. Identification and Functional Characterization of CsMYCs in Cucumber Glandular Trichome Development. Int. J. Mol. Sci. 2023, 24, 6435. https://doi.org/10.3390/ijms24076435
Feng Z, Sun L, Dong M, Fan S, Shi K, Qu Y, Zhu L, Shi J, Wang W, Liu Y, et al. Identification and Functional Characterization of CsMYCs in Cucumber Glandular Trichome Development. International Journal of Molecular Sciences. 2023; 24(7):6435. https://doi.org/10.3390/ijms24076435
Chicago/Turabian StyleFeng, Zhongxuan, Lei Sun, Mingming Dong, Shanshan Fan, Kexin Shi, Yixin Qu, Liyan Zhu, Jinfeng Shi, Wujun Wang, Yihan Liu, and et al. 2023. "Identification and Functional Characterization of CsMYCs in Cucumber Glandular Trichome Development" International Journal of Molecular Sciences 24, no. 7: 6435. https://doi.org/10.3390/ijms24076435