
IL-10 Promotes CXCL13 Expression in Macrophages Following Foot-and-Mouth Disease Virus Infection

 , and
, and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. CXCL13 Was Expressed in Macrophages and T Cells from the Spleen of FMDV-Infected Mice
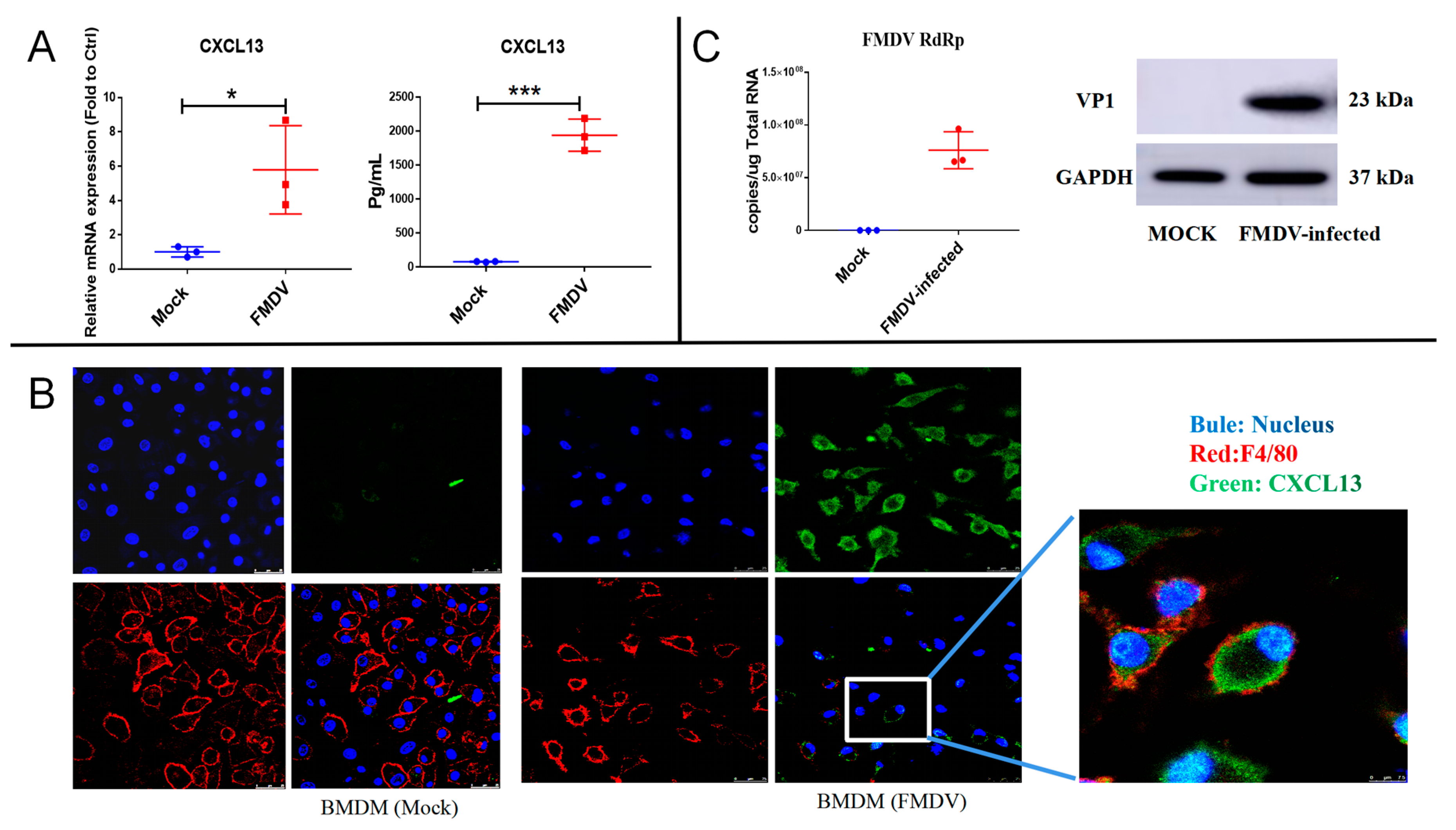
2.2. CXCL13 Was Expressed in Mice BMDMs following FMDV Infection
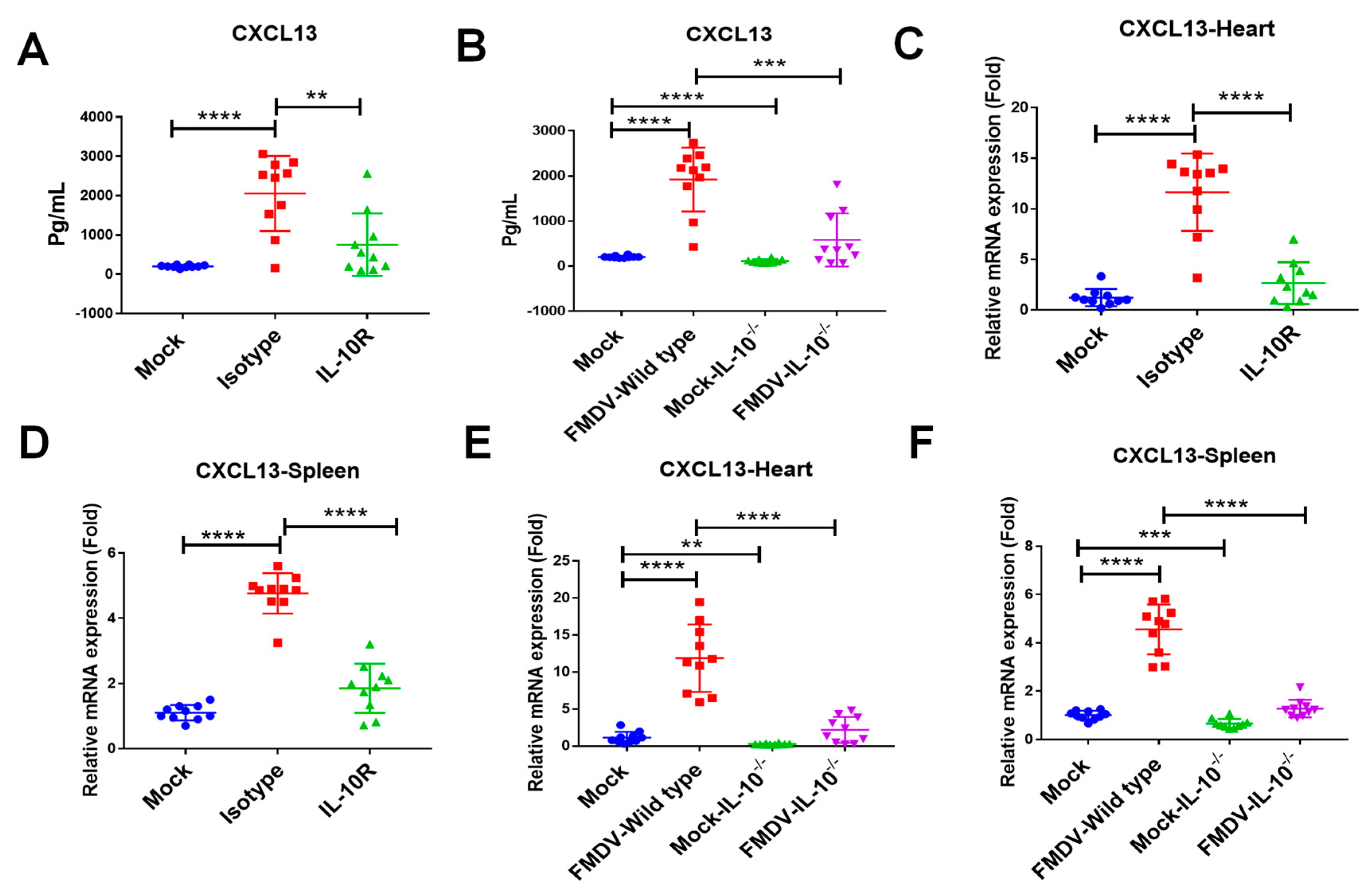
2.3. IL-10 Mediated FMDV-Induced CXCL13 Production in Mice
2.4. IL-10 Enhanced CXCL13 Expression in FMDV-Infected BMDMs
2.5. FMDV Induced CXCL13 Expression in BMDMs by Activating NF-κB and JAK/STAT Signaling Pathways
2.6. IL-10 Induced CXCL13 Expression in BMDMs by Activating JAK/STAT Signaling Pathway following FMDV Infection
3. Discussion
4. Materials and Methods
4.1. Mice, Cells and Virus
4.2. Antibodies and Chemicals
4.3. Infection and Preparation of Samples
4.4. RNA Extraction and cDNA Synthesis
4.5. RT-qPCR
4.6. Cell Sorting and RNA Analysis
4.7. Immunofluorescence
4.8. Western Blotting
4.9. In Vivo Antibody Treatment
4.10. ELISA
4.11. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Kazanietz, M.G.; Durando, M.; Cooke, M. CXCL13 and Its Receptor CXCR5 in Cancer: Inflammation, Immune Response, and Beyond. Front. Endocrinol. 2019, 10, 471. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bekele Feyissa, Y.; Chiodi, F.; Sui, Y.; Berzofsky, J.A. The Role of CXCL13 in Antibody Responses to HIV-1 Infection and Vaccination. Front. Immunol. 2021, 12, 638872. [Google Scholar] [CrossRef] [PubMed]
- Tsai, C.H.; Chen, C.J.; Gong, C.L.; Liu, S.C.; Chen, P.C.; Huang, C.C.; Hu, S.L.; Wang, S.W.; Tang, C.H. CXCL13/CXCR5 axis facilitates endothelial progenitor cell homing and angiogenesis during rheumatoid arthritis progression. Cell Death Dis. 2021, 12, 846. [Google Scholar] [CrossRef]
- Litsiou, E.; Semitekolou, M.; Galani, I.E.; Morianos, I.; Tsoutsa, A.; Kara, P.; Rontogianni, D.; Bellenis, I.; Konstantinou, M.; Potaris, K.; et al. CXCL13 production in B cells via Toll-like receptor/lymphotoxin receptor signaling is involved in lymphoid neogenesis in chronic obstructive pulmonary disease. Am. J. Respir. Crit. Care Med. 2013, 187, 1194–1202. [Google Scholar] [CrossRef] [PubMed]
- Wang, B.; Wang, M.; Ao, D.; Wei, X. CXCL13-CXCR5 axis: Regulation in inflammatory diseases and cancer. Biochim. Biophys. Acta. Rev. Cancer 2022, 1877, 188799. [Google Scholar] [CrossRef]
- Suto, H.; Katakai, T.; Sugai, M.; Kinashi, T.; Shimizu, A. CXCL13 production by an established lymph node stromal cell line via lymphotoxin-beta receptor engagement involves the cooperation of multiple signaling pathways. Int. Immunol. 2009, 21, 467–476. [Google Scholar] [CrossRef] [Green Version]
- Salabè, G.B. Pathogenesis of thyroid nodules: Histological classification? Biomed. Pharmacother. 2001, 55, 39–53. [Google Scholar] [CrossRef]
- Jiang, J.; Cao, Z.; Qu, J.; Liu, H.; Han, H.; Cheng, X. PD-1-expressing MAIT cells from patients with tuberculosis exhibit elevated production of CXCL13. Scand. J. Immunol. 2020, 91, e12858. [Google Scholar] [CrossRef]
- Gu-Trantien, C.; Migliori, E.; Buisseret, L.; de Wind, A.; Brohée, S.; Garaud, S.; Noël, G.; Dang Chi, V.L.; Lodewyckx, J.N.; Naveaux, C.; et al. CXCL13-producing TFH cells link immune suppression and adaptive memory in human breast cancer. JCI Insight 2017, 2, e91487. [Google Scholar] [CrossRef] [Green Version]
- Carlsen, H.S.; Baekkevold, E.S.; Morton, H.C.; Haraldsen, G.; Brandtzaeg, P. Monocyte-like and mature macrophages produce CXCL13 (B cell-attracting chemokine 1) in inflammatory lesions with lymphoid neogenesis. Blood 2004, 104, 3021–3027. [Google Scholar] [CrossRef]
- Denton, A.E.; Innocentin, S.; Carr, E.J.; Bradford, B.M.; Lafouresse, F.; Mabbott, N.A.; Mörbe, U.; Ludewig, B.; Groom, J.R.; Good-Jacobson, K.L.; et al. Type I interferon induces CXCL13 to support ectopic germinal center formation. J. Exp. Med. 2019, 216, 621–637. [Google Scholar] [CrossRef] [Green Version]
- Bellamri, N.; Viel, R.; Morzadec, C.; Lecureur, V.; Joannes, A.; de Latour, B.; Llamas-Gutierrez, F.; Wollin, L.; Jouneau, S.; Vernhet, L. TNF-α and IL-10 Control CXCL13 Expression in Human Macrophages. J. Immunol. (Baltim. Md. 1950) 2020, 204, 2492–2502. [Google Scholar] [CrossRef]
- Horspool, A.M.; Kieffer, T.; Russ, B.P.; DeJong, M.A.; Wolf, M.A.; Karakiozis, J.M.; Hickey, B.J.; Fagone, P.; Tacker, D.H.; Bevere, J.R.; et al. Interplay of Antibody and Cytokine Production Reveals CXCL13 as a Potential Novel Biomarker of Lethal SARS-CoV-2 Infection. mSphere 2021, 6, e01324-20. [Google Scholar] [CrossRef]
- Mehraj, V.; Ramendra, R.; Isnard, S.; Dupuy, F.P.; Lebouché, B.; Costiniuk, C.; Thomas, R.; Szabo, J.; Baril, J.G.; Trottier, B.; et al. CXCL13 as a Biomarker of Immune Activation During Early and Chronic HIV Infection. Front. Immunol. 2019, 10, 289. [Google Scholar] [CrossRef]
- Li, Y.; Tang, L.; Guo, L.; Chen, C.; Gu, S.; Zhou, Y.; Ye, G.; Li, X.; Wang, W.; Liao, X.; et al. CXCL13-mediated recruitment of intrahepatic CXCR5(+)CD8(+) T cells favors viral control in chronic HBV infection. J. Hepatol. 2020, 72, 420–430. [Google Scholar] [CrossRef] [Green Version]
- Perreau, M.; Suffiotti, M.; Marques-Vidal, P.; Wiedemann, A.; Levy, Y.; Laouénan, C.; Ghosn, J.; Fenwick, C.; Comte, D.; Roger, T.; et al. The cytokines HGF and CXCL13 predict the severity and the mortality in COVID-19 patients. Nat. Commun. 2021, 12, 4888. [Google Scholar] [CrossRef]
- Belsham, G.J.; Kristensen, T.; Jackson, T. Foot-and-mouth disease virus: Prospects for using knowledge of virus biology to improve control of this continuing global threat. Virus Res. 2020, 281, 197909. [Google Scholar] [CrossRef]
- Guo, Z.; Zhao, Y.; Zhang, Z.; Li, Y. Interleukin-10-Mediated Lymphopenia Caused by Acute Infection with Foot-and-Mouth Disease Virus in Mice. Viruses 2021, 13, 2358. [Google Scholar] [CrossRef]
- Dos Anjos Cassado, A. F4/80 as a Major Macrophage Marker: The Case of the Peritoneum and Spleen. Results Probl. Cell Differ. 2017, 62, 161–179. [Google Scholar] [CrossRef]
- Bautista, E.M.; Ferman, G.S.; Golde, W.T. Induction of lymphopenia and inhibition of T cell function during acute infection of swine with foot and mouth disease virus (FMDV). Vet. Immunol. Immunopathol. 2003, 92, 61–73. [Google Scholar] [CrossRef]
- Nfon, C.K.; Toka, F.N.; Kenney, M.; Pacheco, J.M.; Golde, W.T. Loss of plasmacytoid dendritic cell function coincides with lymphopenia and viremia during foot-and-mouth disease virus infection. Viral Immunol. 2010, 23, 29–41. [Google Scholar] [CrossRef] [PubMed]
- Yim, L.Y.; Lau, C.S.; Chan, V.S. Heightened TLR7/9-Induced IL-10 and CXCL13 Production with Dysregulated NF-ĸB Activation in CD11c(hi)CD11b(+) Dendritic Cells in NZB/W F1 Mice. Int. J. Mol. Sci. 2019, 20, 4639. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mori, N.; Yamada, Y.; Ikeda, S.; Yamasaki, Y.; Tsukasaki, K.; Tanaka, Y.; Tomonaga, M.; Yamamoto, N.; Fujii, M. Bay 11-7082 inhibits transcription factor NF-kappaB and induces apoptosis of HTLV-I-infected T-cell lines and primary adult T-cell leukemia cells. Blood 2002, 100, 1828–1834. [Google Scholar] [CrossRef]
- Hu, H.; Piotrowska, Z.; Hare, P.J.; Chen, H.; Mulvey, H.E.; Mayfield, A.; Noeen, S.; Kattermann, K.; Greenberg, M.; Williams, A.; et al. Three subtypes of lung cancer fibroblasts define distinct therapeutic paradigms. Cancer Cell 2021, 39, 1531–1547.e1510. [Google Scholar] [CrossRef]
- Stenfeldt, C.; Diaz-San Segundo, F.; de Los Santos, T.; Rodriguez, L.L.; Arzt, J. The Pathogenesis of Foot-and-Mouth Disease in Pigs. Front. Vet. Sci. 2016, 3, 41. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singleton, H.; Graham, S.P.; Frossard, J.P.; Bodman-Smith, K.B.; Steinbach, F. Infection of monocytes with European porcine reproductive and respiratory syndrome virus (PRRSV-1) strain Lena is significantly enhanced by dexamethasone and IL-10. Virology 2018, 517, 199–207. [Google Scholar] [CrossRef]
- Huang, C.; Zhu, J.; Wang, L.; Chu, A.; Yin, Y.; Vali, K.; Garmendia, A.; Tang, Y. Cryptotanshinone protects porcine alveolar macrophages from infection with porcine reproductive and respiratory syndrome virus. Antivir. Res. 2020, 183, 104937. [Google Scholar] [CrossRef]
- Cohen, K.W.; Dugast, A.S.; Alter, G.; McElrath, M.J.; Stamatatos, L. HIV-1 single-stranded RNA induces CXCL13 secretion in human monocytes via TLR7 activation and plasmacytoid dendritic cell-derived type I IFN. J. Immunol. (Baltim. Md. 1950) 2015, 194, 2769–2775. [Google Scholar] [CrossRef] [Green Version]
- Neyt, K.; GeurtsvanKessel, C.H.; Deswarte, K.; Hammad, H.; Lambrecht, B.N. Early IL-1 Signaling Promotes iBALT Induction after Influenza Virus Infection. Front. Immunol. 2016, 7, 312. [Google Scholar] [CrossRef] [Green Version]
- Barone, F.; Nayar, S.; Campos, J.; Cloake, T.; Withers, D.R.; Toellner, K.M.; Zhang, Y.; Fouser, L.; Fisher, B.; Bowman, S.; et al. IL-22 regulates lymphoid chemokine production and assembly of tertiary lymphoid organs. Proc. Natl. Acad. Sci. USA 2015, 112, 11024–11029. [Google Scholar] [CrossRef] [Green Version]
- Bénézech, C.; Luu, N.T.; Walker, J.A.; Kruglov, A.A.; Loo, Y.; Nakamura, K.; Zhang, Y.; Nayar, S.; Jones, L.H.; Flores-Langarica, A.; et al. Inflammation-induced formation of fat-associated lymphoid clusters. Nat. Immunol. 2015, 16, 819–828. [Google Scholar] [CrossRef]
- Ouyang, W.; O’Garra, A. IL-10 Family Cytokines IL-10 and IL-22: From Basic Science to Clinical Translation. Immunity 2019, 50, 871–891. [Google Scholar] [CrossRef]
- Schmetterer, K.G.; Pickl, W.F. The IL-10/STAT3 axis: Contributions to immune tolerance by thymus and peripherally derived regulatory T-cells. Eur. J. Immunol. 2017, 47, 1256–1265. [Google Scholar] [CrossRef] [Green Version]
- Manzanero, S. Generation of mouse bone marrow-derived macrophages. Methods Mol. Biol. 2012, 844, 177–181. [Google Scholar] [CrossRef]
- Sun, P.; Zhang, S.; Qin, X.; Chang, X.; Cui, X.; Li, H.; Zhang, S.; Gao, H.; Wang, P.; Zhang, Z.; et al. Foot-and-mouth disease virus capsid protein VP2 activates the cellular EIF2S1-ATF4 pathway and induces autophagy via HSPB1. Autophagy 2018, 14, 336–346. [Google Scholar] [CrossRef]
- Han, F.L.; Liang, F.; Jiang, T.C.; Liu, M. Increased expression of CXCR5 and CXCL13 in mice with experimental autoimmune myocarditis. Eur. Rev. Med. Pharmacol. Sci. 2017, 21, 1860–1867. [Google Scholar]
- Sebastian, R.; Sravanthi, M.; Umapathi, V.; Krishnaswamy, N.; Priyanka, M.; Dechamma, H.J.; Ganesh, K.; Basagoudanavar, S.H.; Sanyal, A.; Reddy, G.R. Foot and mouth disease virus undergoes non-progressive replication in mice peritoneal macrophages and induces M1 polarization. Virus Res. 2020, 281, 197906. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Bustin, S.A.; Benes, V.; Garson, J.A.; Hellemans, J.; Huggett, J.; Kubista, M.; Mueller, R.; Nolan, T.; Pfaffl, M.W.; Shipley, G.L.; et al. The MIQE guidelines: Minimum information for publication of quantitative real-time PCR experiments. Clin. Chem. 2009, 55, 611–622. [Google Scholar] [CrossRef] [Green Version]
- Burrack, K.S.; Huggins, M.A.; Taras, E.; Dougherty, P.; Henzler, C.M.; Yang, R.; Alter, S.; Jeng, E.K.; Wong, H.C.; Felices, M.; et al. Interleukin-15 Complex Treatment Protects Mice from Cerebral Malaria by Inducing Interleukin-10-Producing Natural Killer Cells. Immunity 2018, 48, 760–772.e4. [Google Scholar] [CrossRef] [Green Version]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Guo, Z.; Chen, F.; Zhao, S.; Zhang, Z.; Zhang, H.; Bai, L.; Zhang, Z.; Li, Y. IL-10 Promotes CXCL13 Expression in Macrophages Following Foot-and-Mouth Disease Virus Infection. Int. J. Mol. Sci. 2023, 24, 6322. https://doi.org/10.3390/ijms24076322
Guo Z, Chen F, Zhao S, Zhang Z, Zhang H, Bai L, Zhang Z, Li Y. IL-10 Promotes CXCL13 Expression in Macrophages Following Foot-and-Mouth Disease Virus Infection. International Journal of Molecular Sciences. 2023; 24(7):6322. https://doi.org/10.3390/ijms24076322
Chicago/Turabian StyleGuo, Zijing, Fei Chen, Shuaiyang Zhao, Zhixiong Zhang, Huijun Zhang, Ling Bai, Zhidong Zhang, and Yanmin Li. 2023. "IL-10 Promotes CXCL13 Expression in Macrophages Following Foot-and-Mouth Disease Virus Infection" International Journal of Molecular Sciences 24, no. 7: 6322. https://doi.org/10.3390/ijms24076322