
Searching for Osmosensing Determinants in Poplar Histidine-Aspartate Kinases


 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
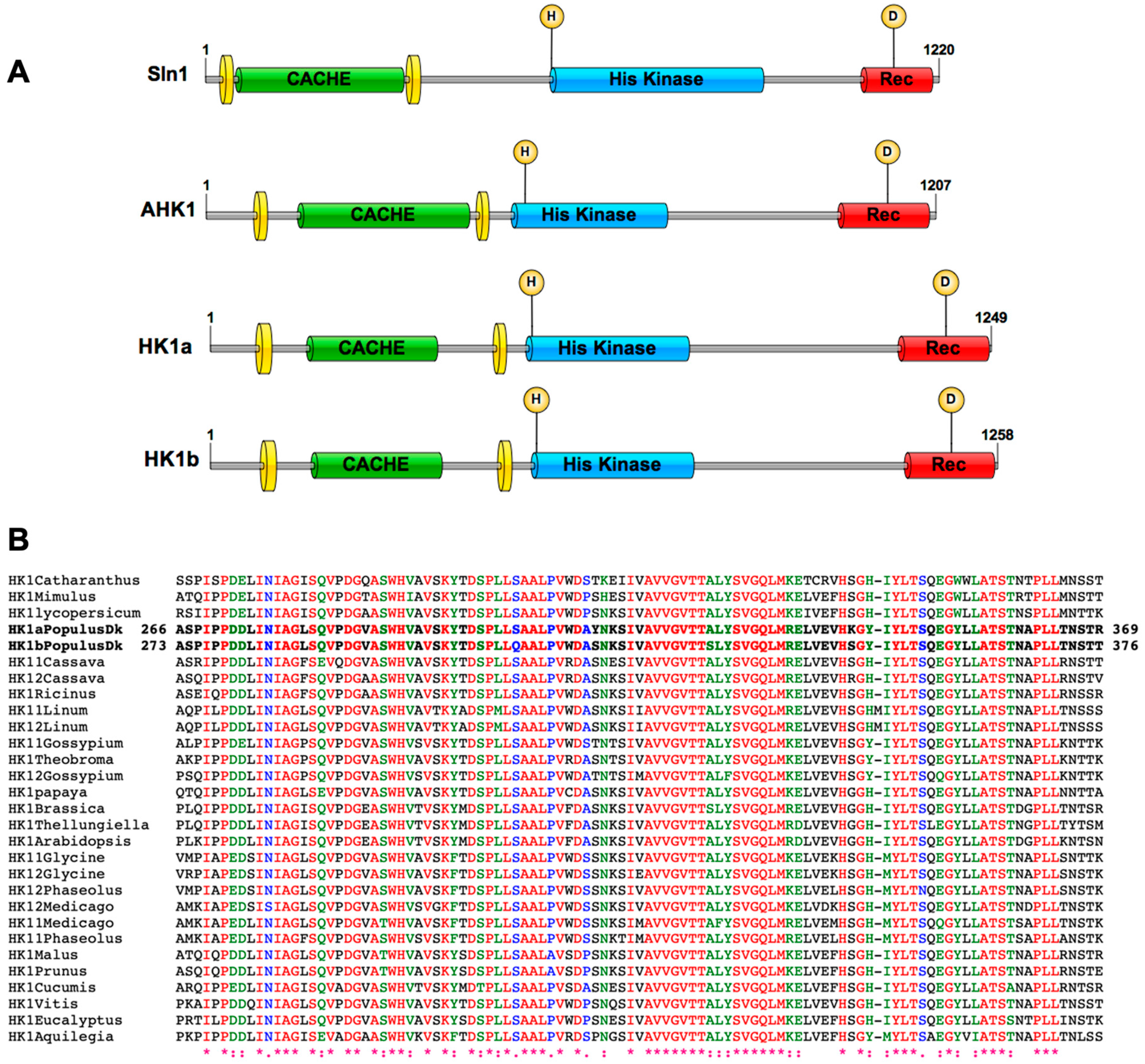
2.1. HK1a/b Contain a Conserved Cache Domain in Their ECD
2.2. Modelling-Assisted Rational Design of Cache Domain Mutants
2.3. Functional Test of Cache Domain Mutants
2.4. Functional Test of TM Mutants
3. Discussion
4. Materials and Methods
4.1. Identification of Cache Domain in Plant AHK1-like Proteins
4.2. Modelling-Assisted Rational Design of Mutants
4.3. Construction of HK1a/b Mutants
4.4. Complementation Analysis of the sln1∆ sho1∆ Deletion Mutant MH179
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Haswell, E.S.; Verslues, P.E. The ongoing search for the molecular basis of plant osmosensing. J. Gen. Physiol. 2015, 145, 389–394. [Google Scholar] [CrossRef]
- Nongpiur, R.C.; Singla-Pareek, S.L.; Pareek, A. The quest for osmosensors in plants. J. Exp. Bot. 2020, 71, 595–607. [Google Scholar] [CrossRef] [PubMed]
- Miermont, A.; Uhlendorf, J.; McClean, M.; Hersen, P. The Dynamical Systems Properties of the HOG Signaling Cascade. J. Signal. Transduct. 2011, 2011, 930940. [Google Scholar] [CrossRef] [Green Version]
- Maeda, T.; Wurgler-Murphy, S.M.; Saito, H. A two-component system that regulates an osmosensing MAP kinase cascade in yeast. Nature 1994, 369, 242–245. [Google Scholar] [CrossRef]
- Posas, F.; Wurgler-Murphy, S.M.; Maeda, T.; Witten, E.A.; Thai, T.C.; Saito, H. Yeast HOG1 MAP kinase cascade is regulated by a multistep phosphorelay mechanism in the SLN1-YPD1-SSK1 “two-component” osmosensor. Cell 1996, 86, 865–875. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, J.M.Y.; Deschenes, R.J.; Fassler, J.S. Saccharomyces cerevisiae histidine phosphotransferase Ypd1p shuttles between the nucleus and cytoplasm for SLN1-dependent phosphorylation of Ssk1p and Skn7p. Eukaryot. Cell 2003, 2, 1304–1314. [Google Scholar] [CrossRef] [Green Version]
- Reiser, V.; Raitt, D.C.; Saito, H. Yeast osmosensor Sln1 and plant cytokinin receptor Cre1 respond to changes in turgor pressure. J. Cell Biol. 2003, 161, 1035–1040. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Urao, T.; Yakubov, B.; Satoh, R.; Yamaguchi-Shinozaki, K.; Seki, M.; Hirayama, T.; Shinozaki, K. A transmembrane hybrid-type histidine kinase in Arabidopsis functions as an osmosensor. Plant Cell 1999, 11, 1743–1754. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tran, L.-S.P.; Urao, T.; Qin, F.; Maruyama, K.; Kakimoto, T.; Shinozaki, K.; Yamaguchi-Shinozaki, K. Functional analysis of AHK1/ATHK1 and cytokinin receptor histidine kinases in response to abscisic acid, drought, and salt stress in Arabidopsis. Proc. Natl. Acad. Sci. USA 2007, 104, 20623–20628. [Google Scholar] [CrossRef] [Green Version]
- Caesar, K.; Thamm, A.M.K.; Witthöft, J.; Elgass, K.; Huppenberger, P.; Grefen, C.; Horak, J.; Harter, K. Evidence for the localization of the Arabidopsis cytokinin receptors AHK3 and AHK4 in the endoplasmic reticulum. J. Exp. Bot. 2011, 62, 5571–5580. [Google Scholar] [CrossRef] [Green Version]
- Wohlbach, D.J.; Quirino, B.F.; Sussman, M.R. Analysis of the Arabidopsis histidine kinase ATHK1 reveals a connection between vegetative osmotic stress sensing and seed maturation. Plant Cell 2008, 20, 1101–1117. [Google Scholar] [CrossRef] [Green Version]
- Kumar, M.N.; Jane, W.N.; Verslues, P.E. Role of the putative osmosensor Arabidopsis histidine kinase1 in zdehydration avoidance and low-water-potential response. Plant Physiol. 2013, 161, 942–953. [Google Scholar] [CrossRef] [Green Version]
- Chefdor, F.; Bénédetti, H.; Depierreux, C.; Delmotte, F.; Morabito, D.; Carpin, S. Osmotic stress sensing in Populus: Components identification of a phosphorelay system. FEBS Lett. 2006, 580, 77–81. [Google Scholar] [CrossRef] [PubMed]
- Héricourt, F.; Chefdor, F.; Djeghdir, I.; Larcher, M.; Lafontaine, F.; Courdavault, V.; Auguin, D.; Coste, F.; Depierreux, C.; Tanigawa, M.; et al. Functional Divergence of Poplar Histidine-Aspartate Kinase HK1 Paralogs in Response to Osmotic Stress. Int. J. Mol. Sci. 2016, 17, 2061. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Héricourt, F.; Chefdor, F.; Bertheau, L.; Tanigawa, M.; Maeda, T.; Guirimand, G.; Courdavault, V.; Larcher, M.; Depierreux, C.; Bénédetti, H. Characterization of histidine-aspartate kinase HK1 and identification of histidine phosphotransfer proteins as potential partners in a Populus multistep phosphorelay. Physiol. Plant. 2013, 149, 188–199. [Google Scholar] [CrossRef] [PubMed]
- Bertheau, L.; Chefdor, F.; Guirimand, G.; Courdavault, V.; Depierreux, C.; Morabito, D.; Brignolas, F.; Héricourt, F.; Carpin, S. Identification of five B-type response regulators as members of a multistep phosphorelay system interacting with histidine-containing phosphotransfer partners of Populus osmosensor. BMC Plant Biol. 2012, 12, 241. [Google Scholar] [CrossRef] [Green Version]
- Bertheau, L.; Miranda, M.; Foureau, E.; Rojas Hoyos, L.F.; Chefdor, F.; Héricourt, F.; Depierreux, C.; Morabito, D.; Papon, N.; Clastre, M.; et al. In planta validation of HK1 homodimerization and recruitment of preferential HPt downstream partners involved in poplar multistep phosphorelay systems. Plant Biosyst.—Int. J. Deal. Asp. Plant Biol. 2013, 147, 991–995. [Google Scholar]
- Bertheau, L.; Djeghdir, I.; Foureau, E.; Chefdor, F.; Glevarec, G.; Oudin, A.; Depierreux, C.; Morabito, D.; Brignolas, F.; Courdavolt, V.; et al. Insights into B-type RR members as signaling partners acting downstream of HPt partners of HK1 in the osmotic stress response in Populus. Plant. Physiol. Biochem. 2015, 94, 244–252. [Google Scholar] [CrossRef] [PubMed]
- Chefdor, F.; Héricourt, F.; Koudounas, K.; Carqueijeiro, I.; Courdavault, V.; Mascagni, F.; Bertheau, L.; Larcher, M.; Depierreux, C.; Lamblin, F.; et al. Highlighting type A RRs as potential regulators of the dkHK1 multi-step phosphorelay pathway in Populus. Plant Sci. 2018, 277, 68–78. [Google Scholar] [CrossRef]
- Djeghdir, I.; Chefdor, F.; Bertheau, L.; Koudounas, K.; Carqueijeiro, I.; Lemos Cruz, P.; Courdavault, V.; Depierreux, C.; Larcher, M.; Lamblin, F.; et al. Evaluation of type-B RR dimerization in poplar: A mechanism to preserve signaling specificity? Plant Sci. 2021, 313, 111068. [Google Scholar] [CrossRef]
- Héricourt, F.; Larcher, M.; Chefdor, F.; Koudounas, K.; Carqueijeiro, I.; Lemos Cruz, P.; Courdavault, V.; Tanigawa, M.; Maeda, T.; Depierreux, C.; et al. New Insight into HPts as Hubs in Poplar Cytokinin and Osmosensing Multistep Phosphorelays: Cytokinin Pathway Uses Specific HPts. Plants 2019, 8, 591. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Upadhyay, A.A.; Fleetwood, A.D.; Adebali, O.; Finn, R.D.; Zhulin, I.B. Cache Domains That are Homologous to, but Different from PAS Domains Comprise the Largest Superfamily of Extracellular Sensors in Prokaryotes. PLoS Comput. Biol. 2016, 12, e1004862. [Google Scholar] [CrossRef] [Green Version]
- Anantharaman, V.; Aravind, L. Cache—A signaling domain common to animal Ca2+-channel subunits and a class of prokaryotic chemotaxis receptors. Trends Biochem. Sci. 2000, 25, 535–537. [Google Scholar] [CrossRef] [PubMed]
- Miwa, K.; Ishikawa, K.; Terada, K.; Yamada, H.; Suzuki, T.; Yamashino, T.; Mizuno, T. Identification of amino acid substitutions that render the Arabidopsis cytokinin receptor histidine kinase AHK4 constitutively active. Plant Cell Physiol. 2007, 48, 1809–1814. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bartrina, I.; Jensen, H.; Novák, O.; Strnad, M.; Werner, T.; Schmülling, T. Gain-of-Function Mutants of the Cytokinin Receptors AHK2 and AHK3 Regulate Plant Organ Size, Flowering Time and Plant Longevity. Plant. Physiol. 2017, 173, 1783–1797. [Google Scholar] [CrossRef] [Green Version]
- Baek, M.; DiMaio, F.; Anishchenko, I.; Dauparas, J.; Ovchinnikov, S.; Lee, G.R.; Wang, J.; Cong, Q.; Kinch, L.N.; Schaeffer, R.D.; et al. Accurate prediction of protein structures and interactions using a three-track neural network. Science 2021, 373, 871–876. [Google Scholar] [CrossRef] [PubMed]
- Holm, L. Dali server: Structural unification of protein families. Nucleic Acids Res. 2022, 50, W2015–W2215. [Google Scholar] [CrossRef]
- Corral-Lugo, A.; Matilla, M.A.; Martín-Mora, D.; Silva Jiménez, H.; Mesa Torres, N.; Kato, J.; Hida, A.; Oku, S.; Conejero-Muriel, M.; Gavira, J.A.; et al. High-affinity chemotaxis to histamine mediated by the TlpQ chemoreceptor of the human pathogen. Pseudomonas aeruginosa. mBio 2018, 9, e01894-18. [Google Scholar] [CrossRef] [Green Version]
- Kooyers, N.J.; Greenlee, A.B.; Colicchio, J.M.; Oh, M.; Blackman, B.K. Replicate altitudinal clines reveal that evolutionary flexibility underlies adaptation to drought stress in annual Mimulus guttatus. New Phytol. 2015, 206, 152–165. [Google Scholar] [CrossRef]
- Khan, A.; Sovero, V.; Gemenet, D. Genome-assisted Breeding For Drought Resistance. Curr. Genom. 2016, 17, 330–342. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.C.; Machuca, M.A.; Beckham, S.A.; Gunzburg, M.; Roujeinikova, A. Structural basis for amino-acid recognition and transmembrane signalling by tandem Per–Arnt–Sim (tandem PAS) chemoreceptor sensory domains. Acta Crystallogr. Sect. D Biol. Crystallogr. 2015, 71, 2127–2136. [Google Scholar] [CrossRef]
- Shankarnarayan, S.; Malone, C.L.; Deschenes, R.J.; Fassler, J.S. Modulation of yeast Sln1 kinase activity by the CCW12 cell wall protein. J. Biol. Chem. 2008, 283, 1962–1973. [Google Scholar] [PubMed] [Green Version]
- Inoue, T.; Higuchi, M.; Hashimoto, Y.; Seki, M.; Kobayashi, M.; Kato, T.; Tabata, S.; Shinozaki, K.; Kakimoto, T. Identification of CRE1 as a cytokinin receptor from Arabidopsis. Nature 2001, 409, 1060–1063. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, T.; Miwa, K.; Ishikawa, K.; Yamada, H.; Aiba, H.; Mizuno, T. The Arabidopsis Sensor His-kinase, AHK4, Can Respond to Cytokinins. Plant Cell Physiol. 2001, 42, 107–113. [Google Scholar] [CrossRef] [PubMed]
- Wulfetange, K.; Lomin, S.N.; Romanov, G.A.; Stolz, A.; Heyl, A.; Schmülling, T. The cytokinin receptors of Arabidopsis are located mainly to the endoplasmic reticulum. Plant. Physiol. 2011, 156, 1808–1818. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lomin, S.N.; Myakushina, Y.A.; Arkhipov, D.V.; Leonova, O.G.; Popenko, V.I.; Schmülling, T.; Romanov, G.A. Studies of cytokinin receptor-phosphotransmitter interaction provide evidences for the initiation of cytokinin signalling in the endoplasmic reticulum. Funct. Plant Biol. FPB 2018, 45, 192–202. [Google Scholar] [CrossRef]
- Arkhipov, D.V.; Lomin, S.N.; Myakushina, Y.A.; Savelieva, E.M.; Osolodkin, D.I.; Romanov, G.A. Modeling of Protein–Protein Interactions in Cytokinin Signal Transduction. Int. J. Mol. Sci. 2019, 20, 2096. [Google Scholar] [CrossRef] [Green Version]
- Li, W.; Herrera-Estrella, L.; Tran, L.-S.P. The Yin-Yang of cytokinin homeostasis and drought acclimation/adaptation. Trends Plant Sci. 2016, 21, 548–550. [Google Scholar] [CrossRef]
- Gietz, R.D.; Schiestl, R.H. Transforming yeast with DNA. Method Mol. Cell Biol. 1995, 5, 255–269. [Google Scholar]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Makhokh, H.; Lafite, P.; Larcher, M.; Lamblin, F.; Chefdor, F.; Depierreux, C.; Tanigawa, M.; Maeda, T.; Carpin, S.; Héricourt, F. Searching for Osmosensing Determinants in Poplar Histidine-Aspartate Kinases. Int. J. Mol. Sci. 2023, 24, 6318. https://doi.org/10.3390/ijms24076318
Makhokh H, Lafite P, Larcher M, Lamblin F, Chefdor F, Depierreux C, Tanigawa M, Maeda T, Carpin S, Héricourt F. Searching for Osmosensing Determinants in Poplar Histidine-Aspartate Kinases. International Journal of Molecular Sciences. 2023; 24(7):6318. https://doi.org/10.3390/ijms24076318
Chicago/Turabian StyleMakhokh, Hanae, Pierre Lafite, Mélanie Larcher, Frédéric Lamblin, Françoise Chefdor, Christiane Depierreux, Mirai Tanigawa, Tatsuya Maeda, Sabine Carpin, and François Héricourt. 2023. "Searching for Osmosensing Determinants in Poplar Histidine-Aspartate Kinases" International Journal of Molecular Sciences 24, no. 7: 6318. https://doi.org/10.3390/ijms24076318