
Current Approach to the Pathogenesis of Diabetic Cataracts

{kind=link}
{kind=link}
Abstract
:1. Introduction
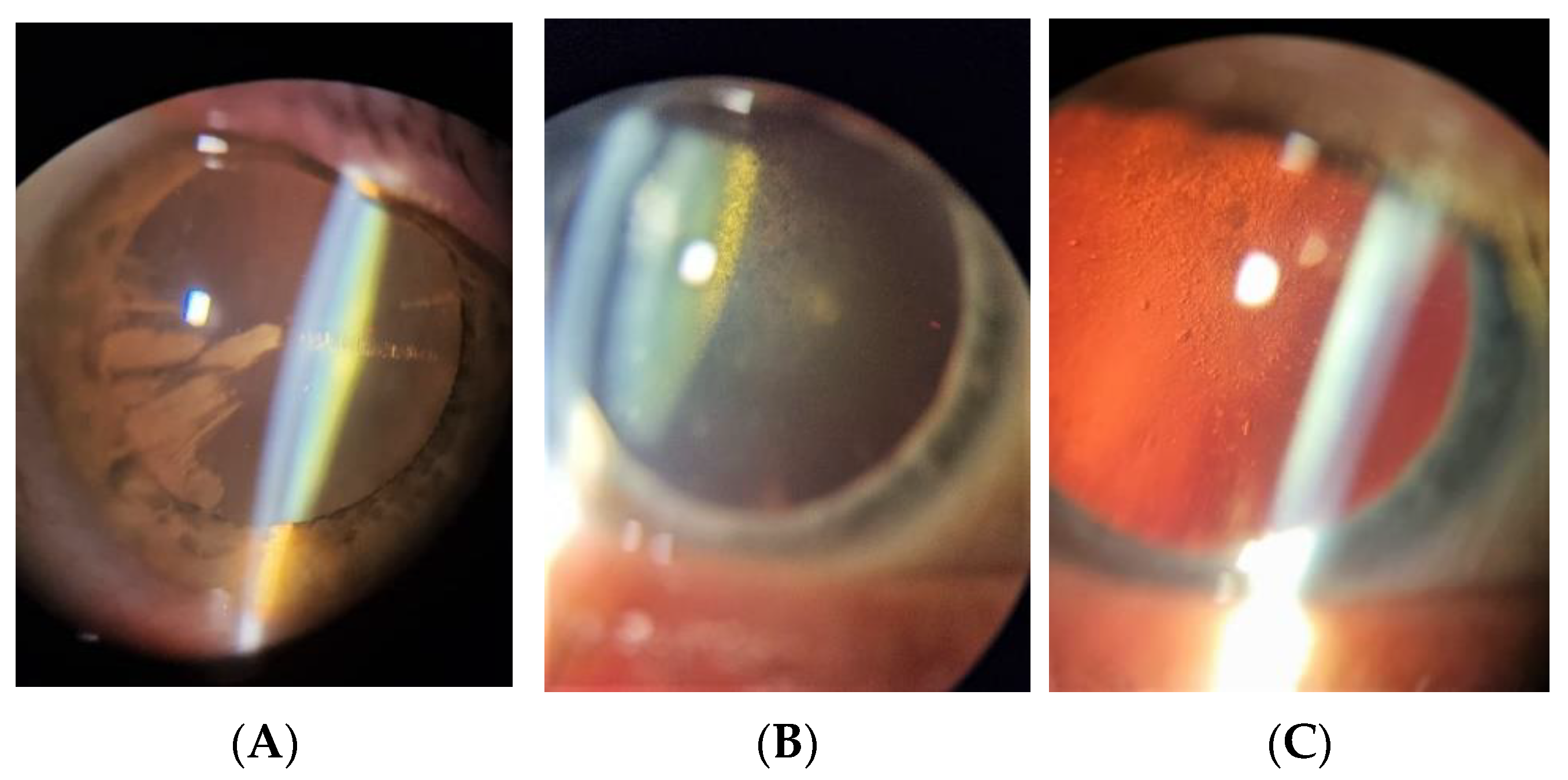
1.1. Diabetic Cataract Pathogenesis
1.2. The Sorbitol Pathway
1.3. Oxidative Stress
1.4. Autophagy Lysosomal Pathway
1.5. Changes in the Metabolic Compositions in the Aqueous Humor
1.6. The Influence of Diet on the Development of Cataracts
2. Challenges and Future Directions
3. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- IDF–International Diabetes Federation. Available online: www.idf.org (accessed on 5 August 2022).
- Chiang, P.P.; Lamoureux, E.L.; Zheng, Y.; Tay, W.T.; Mitchell, P.; Wang, J.J.; Wong, T.Y. Frequency and risk factors of non-retinopathy ocular conditions in people with diabetes: The Singapore Malay Eye Study. Diabet. Med. 2013, 30, e32–e40. [Google Scholar] [CrossRef] [PubMed]
- Memon, A.F.; Mahar, P.S.; Memon, M.S.; Mumtaz, S.N.; Shaikh, S.A.; Fahim, M.F. Age-related cataract and its types in patients with and without type 2 diabetes mellitus: A Hospital-based comparative study. J. Pak. Med. Assoc. 2016, 66, 1272–1276. [Google Scholar]
- Becker, C.; Schneider, C.; Aballéa, S.; Bailey, C.; Bourne, R.; Jick, S.; Meier, C. Cataract in patients with diabetes mellitus-incidence rates in the UK and risk factors. Eye 2018, 32, 1028–1035. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Olafsdottir, E.; Andersson, D.K.; Stefansson, E. The prevalence of cataract in a population wirh and without type 2 diabetes mellitus. Acta Ophthalmol. 2012, 90, 334–340. [Google Scholar] [CrossRef] [PubMed]
- Alabdulwahhab, K.M. Senile Cataract in Patients with Diabetes with and Without Diabetic Retinopathy: A Community-Based Comparative Study. J. Epidemiol. Glob. Health 2022, 12, 56–63. [Google Scholar] [CrossRef]
- Pollreisz, A.; Schmidt-Erfurth, U. Diabetic cataract-pathogenesis, epidemiology and treatment. J. Ophthalmol. 2010, 2010, 608751. [Google Scholar] [CrossRef] [Green Version]
- Li, L.; Wan, X.H.; Zhao, G.H. Meta-analysis of the risk of cataract in type 2 diabetes. BMC Ophthalmol. 2014, 14, 94. [Google Scholar] [CrossRef]
- Machan, C.M.; Hrynchak, P.K.; Irving, E.L. Age-related cataract is associated with type 2 diabetes and statin use. Optom. Vis. Sci. 2012, 89, 1165–1171. [Google Scholar] [CrossRef]
- Schäfer, C.; Lautenschläger, C.; Struck, H.G. Kataraktformen bei Diabetikern und Nichtdiabetikern: Eine densitometrische Studie mit der Topcon-Scheimpflug-Kamera [Cataract types in diabetics and non-diabetics: A densitometric study with the Topcon-Scheimpflug camera]. Klin. Mon. Augenheilkd. 2006, 223, 589–592. (In German) [Google Scholar] [CrossRef]
- Srinivasan, S.; Raman, R.; Swaminathan, G.; Ganesan, S.; Kulothungan, V.; Sharma, T. Incidence, Progression, and Risk Factors for Cataract in Type 2 Diabetes. Investig. Ophthalmol. Vis. Sci. 2017, 58, 5921–5929. [Google Scholar] [CrossRef]
- Tan, J.S.; Wang, J.J.; Mitchell, P. Influence of diabetes and cardiovascular disease on the long-term incidence of cataract: The Blue Mountains Eye Study. Ophthalmic Epidemiol. 2008, 15, 317–327. [Google Scholar] [CrossRef]
- Richter, G.M.; Torres, M.; Choudhury, F.; Azen, S.P.; Varma, R.; Los Angeles Latino Eye Study Group. Risk factors for cortical, nuclear, posterior subcapsular, and mixed lens opacities: The Los Angeles Latino Eye Study. Ophthalmology 2012, 119, 547–554. [Google Scholar] [CrossRef] [Green Version]
- Esteves, J.F.; Dal Pizzol, M.M.; Sccoco, C.A.; Roggia, M.F.; Milano, S.B.; Guarienti, J.A.; Rodrigues, T.C.; Canani, L.H. Cataract and type 1 diabetes mellitus. Diabetes Res. Clin. Pract. 2008, 82, 324–328. [Google Scholar] [CrossRef]
- Geloneck, M.M.; Forbes, B.J.; Shaffer, J.; Ying, G.; Binenbaum, G. Ocular complications in children with diabetes mellitus. Ophthalmology 2015, 122, 2457–2464. [Google Scholar] [CrossRef] [Green Version]
- Iafusco, D.; Prisco, F.; Romano, M.R.; Dell’Omo, R.; Libondi, T.; Costagliola, C. Acute juvenile cataract in newly diagnosed type 1 diabetic patients: A description of six cases. Pediatr. Diabetes 2011, 12, 642–648. [Google Scholar] [CrossRef] [PubMed]
- Wilson, M.E., Jr.; Levin, A.V.; Trivedi, R.H.; Kruger, S.J.; Elliott, L.A.; Ainsworth, J.R.; Awner, S.; Cruz, O.A.; Kivlin, J.; Vroman, D.T.; et al. Cataract associated with type-1 diabetes mellitus in the pediatric population. J. Am. Assoc. Pediatr. Ophthalmol. Strabismus 2007, 11, 162–165. [Google Scholar] [CrossRef] [PubMed]
- Skrabic, V.; Ivanisevic, M.; Stanic, R.; Unic, I.; Bucan, K.; Galetovic, D. Acute bilateral cataract with phacomorphic glaucoma in a girl with newly diagnosed type 1 diabetes mellitus. J. Pediatr. Ophthalmol. Strabismus 2010, 47, e1–e3. [Google Scholar] [CrossRef] [PubMed]
- Jin, Y.Y.; Huang, K.; Zou, C.C.; Liang, L.; Wang, X.M.; Jin, J. Reversible cataract as the presenting sign of diabetes mellitus: Report of two cases and literature review. Iran. J. Pediatr. 2012, 22, 125–128. [Google Scholar]
- Song, E.; Qian, D.J.; Wang, S.; Xu, C.; Pan, C.W. Refractive error in Chinese with type 2 diabetes and its association with glycaemic control. Clin. Exp. Optom. 2018, 101, 213–219. [Google Scholar] [CrossRef]
- Wang, Y.H.; Huang, C.; Tseng, Y.L.; Zhong, J.; Li, X.M. Refractive Error and Eye Health: An Umbrella Review of Meta-Analyses. Front. Med. 2021, 8, 759767. [Google Scholar] [CrossRef] [PubMed]
- Kelkar, A.; Kelkar, J.; Mehta, H.; Amoaku, W. Cataract surgery in diabetes mellitus: A systematic review. Indian J. Ophthalmol. 2018, 66, 1401–1410. [Google Scholar] [CrossRef]
- Lin, S.F.; Lin, P.K.; Chang, F.L.; Tsai, R.K. Transient hyperopia after intensive treatment of hyperglycemia in newly diagnosed diabetes. Ophthalmologica 2009, 223, 68–71. [Google Scholar] [CrossRef]
- Mehdizadeh, M.; Nowroozzadeh, M.H. Transient hyperopia and diabetes. Ophthalmologica 2010, 224, 63. [Google Scholar] [CrossRef]
- Sonmez, B.; Bozkurt, B.; Atmaca, A.; Irkec, M.; Orhan, M.; Aslan, U. Effect of glycemic control on refractive changes in diabetic patients with hyperglycemia. Cornea 2005, 24, 531–537. [Google Scholar] [CrossRef] [PubMed]
- García-Sáenz, M.C.; Arias-Puente, A.; Rodríguez-Caravaca, G.; Andrés Alba, Y.; Bañuelos Bañuelos, J. Endophthalmitis after Cataract Surgery: Epidemiology, Clinical Features and Antibiotic Prophylaxis. Arch. Soc. Española Oftalmol. 2010, 85, 263–267. [Google Scholar] [CrossRef] [PubMed]
- Lin, Z.; Wen, L.; Li, D.; Moonasar, N.; Zhai, G.; Wang, Y.; Wang, F.H.; Liang, Y.B. Refractive Error in a Chinese Population with Type 2 Diabetes: A Report from the Fushun Diabetic Retinopathy Cohort Study. Ophthalmic Epidemiol. 2022, 30, 38–45. [Google Scholar] [CrossRef] [PubMed]
- Squirrell, D.; Bhola, R.; Bush, J.; Winder, S.; Talbot, J. A prospective, case controlled study of the natural history of diabetic retinopathy and maculopathy after uncomplicated phacoemulsification cataract surgery in patients with type 2 diabetes. Br. J. Ophthalmol. 2002, 86, 565–571. [Google Scholar] [CrossRef] [Green Version]
- Haddad, N.M.; Sun, J.K.; Abujaber, S.; Schlossman, D.K.; Silva, P.S. Cataract surgery and its complications in diabetic patients. Semin. Ophthalmol. 2014, 29, 329–337. [Google Scholar] [CrossRef]
- Heesterman, B.L.; Hogewind, B.F. Phacoemulsification and Intraoperative Complications in 452 Patients with Diabetic Retinopathy. Semin. Ophthalmol. 2017, 32, 395–396. [Google Scholar] [CrossRef] [PubMed]
- Gawęcki, M.; Prądzyńska, N.; Karska-Basta, I. Long-Term Variations in Retinal Parameters after Uncomplicated Cataract Surgery. J. Clin. Med. 2022, 11, 3426. [Google Scholar] [CrossRef]
- Okamoto, F. Refractive Changes in Diabetic Patients during Intensive Glycaemic Control. Br. J. Ophthalmol. 2000, 84, 1097–1102. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dong, N.; Xu, B.; Wang, B.; Chu, L.; Tang, X. Aqueous cytokines as predictors of macular edema in patients with diabetes following uncomplicated phacoemulsification cataract surgery. BioMed Res. Int. 2015, 2015, 126984. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hua, J.; Guerin, K.I.; Chen, J.; Michán, S.; Stahl, A.; Krah, N.M.; Seaward, M.R.; Dennison, R.J.; Juan, A.M.; Hatton, C.J.; et al. Resveratrol inhibits pathologic retinal neovascularization in VLDLR−/− mice. Investig. Ophthalmol. Vis. Sci. 2011, 52, 2809–2816. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tabin, G.; Chen, M.; Espandar, L. Cataract surgery for the developing world. Curr. Opin. Ophthalmol. 2008, 19, 55–59. [Google Scholar] [CrossRef]
- Hejtmancik, J.F.; Riazuddin, S.A.; McGreal, R.; Liu, W.; Cvekl, A.; Shiels, A. Lens Biology and Biochemistry. Prog. Mol. Biol. Transl. Sci. 2015, 134, 169–201. [Google Scholar]
- Obrosova, I.G.; Chung SS, M.; Kador, P.F. Diabetic cataracts: Mechanisms and management. Diabetes/Metab. Res. Rev. 2010, 26, 172–180. [Google Scholar] [CrossRef]
- Snow, A.; Shieh, B.; Chang, K.C.; Pal, A.; Lenhart, P.; Ammar, D.; Ruzycki, P.; Palla, S.; Reddy, G.B.; Petrash, J.M. Aldose reductase expression as a risk factor for cataract. Chem. Biol. Interact. 2015, 234, 247–253. [Google Scholar] [CrossRef] [Green Version]
- Rosenfeld, S.I.; Blecher, M.H.; Bobrow, J.C.; Bradford, C.A.; Glasser, D.; Berestka, J.S. Basic and Clinical Science Course. Section 11. Lens and Cataract. In Chapter 3: Biochemistry and Physiology; Edra Urban & Partner: Wroclaw, Poland, 2021; pp. 17–24. [Google Scholar]
- Abdul Nasir, N.A.; Agarwal, R.; Sheikh Abdul Kadir, S.H.; Vasudevan, S.; Tripathy, M.; Iezhitsa, I.; Mohammad Daher, A.; Ibrahim, M.I.; Mohd Ismail, N. Reduction of oxidative-nitrosative stress underlies anticataract effect of topically applied tocotrienol in streptozotocin-induced diabetic rats. PLoS ONE 2017, 12, e0174542. [Google Scholar] [CrossRef] [Green Version]
- Kumar, C.U.; Suryavanshi, U.; Sontake, V.; Reddy, P.Y.; Sankhala, R.S.; Swamy, M.J.; Reddy, G.B. Effect of Sorbitol on Alpha-Crystallin Structure and Function. Biochemistry 2022, 87, 131–140. [Google Scholar] [CrossRef] [PubMed]
- Haroon, H.B.; Perumalsamy, V.; Nair, G.; Anand, D.K.; Kolli, R.; Monichen, J.; Prabha, K. Repression of Polyol Pathway Activity by Hemidesmus indicus var. pubescens R.Br. Linn Root Extract, an Aldose Reductase Inhibitor: An In Silico and Ex Vivo Study. Nat. Prod. Bioprospect. 2021, 11, 315–324. [Google Scholar] [CrossRef]
- Lu, W.L.; Shen, P.C.; Lee, C.H.; Su, Y.T.; Chen, L.M. High Risk of Early Cataracts in Young Type 1 Diabetes Group: A Nationwide Cohort Study. Int. J. Endocrinol. 2020, 9, 8160256. [Google Scholar] [CrossRef]
- Papadimitriou, D.T.; Bothou, C.; Skarmoutsos, F.; Papaevangelou, V.; Papadimitriou, A. Acute Bilateral Cataract in Type 1 Diabetes Mellitus. Ann. Pediatr. Child Health 2015, 3, 1080. [Google Scholar]
- Sayin, N.; Kara, N.; Pekel, G. Ocular complications of diabetes mellitus. World J. Diabetes 2015, 15, 92–108. [Google Scholar] [CrossRef]
- Oishi, N.; Morikubo, S.; Takamura, Y.; Kubo, E.; Tsuzuki, S.; Tanimoto, T.; Akagi, Y. Correlation between adult diabetic cataracts and red blood cell aldose reductase levels. Investig. Ophthalmol. Vis. Sci. 2006, 47, 2061–2064. [Google Scholar] [CrossRef]
- Kumamoto, Y.; Takamura, Y.; Kubo, E.; Tsuzuki, S.; Akagi, Y. Epithelial cell density in cataractous lenses of patients with diabetes: Association with erythrocyte aldose reductase. Exp. Eye Res. 2007, 85, 393–399. [Google Scholar] [CrossRef] [PubMed]
- Laspias, G.A.; Thomopoulou, G.H.; Lazaris, A.C.; Kavantzas, N.; Koutselini, H.; Pagonis, N.; Tsapeli, E.; Politi, E. Cytomorphometric study of epithelial cells in normal and cataractous human lenses in relation with hyperglycemia. Int. Ophthalmol. 2016, 36, 147–158. [Google Scholar] [CrossRef]
- Kador, P.F.; Wyman, M.; Oates, P.J. Aldose reductase, ocular diabetic complications and the development of topical Kinostat®. Prog. Retin. Eye Res. 2016, 54, 1–29. [Google Scholar] [CrossRef] [Green Version]
- Braakhuis, A.J.; Donaldson, C.I.; Lim, J.C.; Donaldson, P.J. Nutritional Strategies to Prevent Lens Cataract: Current Status and Future Strategies. Nutrients 2019, 11, 1186. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.Y.; Wu, T.T.; Ho, C.Y.; Yeh, T.C.; Sun, G.C.; Tseng, C.J.; Cheng, P.W. Blocking of SGLT2 to Eliminate NADPH-Induced Oxidative Stress in Lenses of Animals with Fructose-Induced Diabetes Mellitus. Int. J. Mol. Sci. 2022, 23, 7142. [Google Scholar] [CrossRef]
- Kiziltoprak, H.; Tekin, K.; Inanc, M.; Goker, Y.S. Cataract in diabetes mellitus. World J. Diabetes 2019, 10, 140–153. [Google Scholar] [CrossRef]
- Zych, M.; Wojnar, W.; Kielanowska, M.; Folwarczna, J.; Kaczmarczyk-Sedlak, I. Effect of Berberine on Glycation, Aldose Reductase Activity, and Oxidative Stress in the Lenses of Streptozotocin-Induced Diabetic Rats In Vivo-A Preliminary Study. Int. J. Mol. Sci. 2020, 21, 4278. [Google Scholar] [CrossRef] [PubMed]
- Bejarano, E.; Taylor, A. Too sweet: Problems of protein glycation in the eye. Exp. Eye Res. 2019, 178, 255–262. [Google Scholar] [CrossRef] [PubMed]
- Kumar, M.S.; Reddy, P.Y.; Kumar, P.A.; Surolia, I.; Reddy, G.B. Effect of dicarbonyl-induced browning on α-crystallin chaperone-like activity: Physiological significance and caveats of in vitro aggregation assays. Biochem. J. 2004, 379, 273–282. [Google Scholar] [CrossRef] [Green Version]
- Kumar, M.S.; Mrudula, T.; Mitra, N.; Reddy, G.B. Enhanced proteolytic degradation and altered ATP binding of methylglyoxal modified α-crystallin. Exp. Eye Res. 2004, 79, 577–583. [Google Scholar]
- Argirova, M.; Breipohl, W. Comparison between modifications of lens proteins resulted from glycation with methylglyoxal, glyoxal, ascorbic acid, and fructose. J. Biochem. Mol. Toxicol. 2002, 16, 140–145. [Google Scholar] [CrossRef]
- Obrenovich, M.E.; Monnier, V.M. Glycation stimulates amyloid formation. Sci. Aging Knowl. Environ. 2004, 2004, pe3. [Google Scholar] [CrossRef] [PubMed]
- Saremi, L.; Taghvaei, S.; Feizy, F.; Ghaffari, M.E.; Babaniamansour, S.; Saltanatpour, Z. Association study between superoxide Dismutases gene polymorphisms and development of diabetic retinopathy and cataract in Iranian patients with type two diabetes mellitus. J. Diabetes Metab. Disord. 2021, 20, 627–634. [Google Scholar] [CrossRef]
- Yim, M.B.; Yim, H.S.; Lee, C.; Kang, S.O.; Chock, P.B. Protein glycation: Creation of catalytic sites for free radical generation. Ann. N. Y. Acad. Sci. 2001, 928, 48–53. [Google Scholar] [CrossRef]
- Chaudhuri, J.; Bains, Y.; Guha, S.; Kahn, A.; Hall, D.; Bose, N.; Gugliucci, A.; Kapahi, P. The role of advanced glycation end products in aging and metabolic diseases: Bridging association and causality. Cell Metab. 2018, 28, 337–352. [Google Scholar] [CrossRef] [Green Version]
- Wierusz-Wysocka, B.; Araszkiewicz, A.; Schlaffke, J. Advanced glycation end products. A new biomarker of diabetes and late complications of disease? Clin. Diabetol. 2013, 2, 96–103. [Google Scholar]
- Bheda, P. Metabolic transcriptional memory. Mol. Metab. 2020, 38, 100955. [Google Scholar] [CrossRef] [PubMed]
- Hong, S.B.; Lee, K.W.; Handa, J.T.; Joo, C.K. Effect of advanced glycation end products on lens epithelial cells in vitro. Biochem. Biophys. Res. Commun. 2000, 18, 53–59. [Google Scholar] [CrossRef] [PubMed]
- Ranjan, M.; Nayak, S.; Rao, B.S. Immunochemical detection of glycated beta- and gamma-crystallins in lens and their circulating autoantibodies (IgG) in streptozocin induced diabetic rat. Mol. Vis. 2006, 12, 1077–1085. [Google Scholar] [PubMed]
- Rankenberg, J.; Rakete, S.; Wagner, B.D.; Patnaik, J.L.; Henning, C.; Lynch, A.; Glomb, M.A.; Nagaraj, R.H. Advanced glycation end products in human diabetic lens capsules. Exp. Eye Res. 2021, 210, 108704. [Google Scholar] [CrossRef]
- Pokupec, R.; Kalauz, M.; Turk, N.; Turk, Z. Advanced glycation endproducts in human diabetic and non-diabetic cataractous lenses. Graefe’s Arch. Clin. Exp. Ophthalmol. 2003, 241, 378–384. [Google Scholar] [CrossRef]
- Kalousova, M.; Skrha, J.; Zima, T. Advanced glycation end products and advanced oxidation protein products in patients with diabetes mellitus. Physiol. Res. 2002, 51, 597–604. [Google Scholar]
- Rowan, S.; Bejarano, E.; Taylor, A. Mechanistic targeting of advanced glycation end-products in age-related diseases. Biochim. Biophys. Acta (BBA)-Mol. Basis Dis. 2018, 1864, 3631–3643. [Google Scholar] [CrossRef]
- Swamy-Mruthinti, S.; Coral Miriam, K.; Krishna Kumar, S.; Biswas, J.; Ramakrishnan, S.; HNagaraj, R.; NSulochana, K. Immunolocalization and quantification of advanced glycation end products in retinal neovascular membranes and serum: A possible role in ocular neovascularization. Curr. Eye Res. 2002, 25, 139–145. [Google Scholar] [CrossRef]
- Sady, C.; Jiang, C.L.; Chellan, P.; Madhun, Z.; Duve, Y.; A Glomb, M.; Nagaraj, R.H. Maillard reactions by alpha-oxoaldehydes: Detection of glyoxalmodified proteins. Biochim. Biophys. Acta (BBA)-Protein Struct. Mol. Enzym. 2000, 1481, 255–264. [Google Scholar] [CrossRef]
- Derham, B.K.; Harding, J.J. Alpha-crystallin as a molecular chaperone. Prog. Retin. Eye Res. 1999, 18, 463–509. [Google Scholar] [CrossRef]
- Horwitz, J. Alpha-crystallin. Exp. Eye Res. 2003, 76, 145–153. [Google Scholar] [CrossRef]
- Reddy, G.B.; Kumar, P.A.; Kumar, M.S. Critical review: Chaperone-like activity and hydrophobicity of α-crystallin. IUBMB Life 2006, 58, 632–641. [Google Scholar] [CrossRef]
- Kumar, P.A.; Reddy, P.Y.; Srinivas, P.N.; Reddy, G.B. Delay of diabetic cataract in rats by the antiglycating potential of cumin through modulation of alpha-crystallin chaperone activity. J. Nutr. Biochem. 2009, 20, 553–562. [Google Scholar] [CrossRef]
- Thampi, P.; Zarina, S.; Abraham, E.C. alpha-Crystallin chaperone function in diabetic rat and human lenses. Mol. Cell. Biochem. 2002, 229, 113–118. [Google Scholar] [CrossRef]
- Kumar, P.A.; Suryanarayana, P.; Reddy, P.Y.; Reddy, G.B. Modulation of alpha-crystallin chaperone activity in diabetic rat lens by curcumin. Mol. Vis. 2005, 11, 561–568. [Google Scholar]
- Reddy, V.S.; Reddy, G.B. Role of crystallins in diabetic complications. Biochim. Biophys. Acta 2016, 860, 269–277. [Google Scholar] [CrossRef]
- Babizhayev, M.A. Structural and functional properties, chaperone activity and posttranslational modifications of alpha-crystallin and its related subunits in the crystalline lens: N-acetylcarnosine, carnosine and carcinine act as alpha- crystallin/small heat shock protein enhancers in prevention and dissolution of cataract in ocular drug delivery formulations of novel therapeutic agents. Recent Pat. Drug Deliv. Formul. 2012, 6, 107–148. [Google Scholar]
- Rajkumar, S.; Vasavada, A.R.; Praveen, M.R.; Ananthan, R.; Reddy, G.B.; Tripathi, H.; Ganatra, D.A.; Arora, A.I.; Patel, A.R. Exploration of molecular factors impairing superoxide dismutase isoforms activity in human senile cataractous lenses. Investig. Ophthalmol. Vis. Sci. 2013, 54, 6224–6233. [Google Scholar] [CrossRef] [Green Version]
- Behndig, A.; Karlsson, K.; Reaume, A.G.; Sentman, M.-L.; Marklund, S.L. In vitro photochemical cataract in mice lacking copper-zinc superoxide dismutase. Free Radic. Biol. Med. 2001, 31, 738–744. [Google Scholar] [CrossRef]
- Olofsson, E.M.; Marklund, S.L.; Karlsson, K.; Brännström, T.; Behndig, A. In vitro glucose-induced cataract in copper-zinc superoxide dismutase null mice. Exp. Eye Res. 2005, 81, 639–646. [Google Scholar] [CrossRef]
- Olofsson, E.M.; Marklund, S.L.; Behndig, A. Enhanced diabetes-induced cataract in copper-zinc superoxide dismutase-null mice. Investig. Ophthalmol. Vis. Sci. 2009, 50, 2913–2918. [Google Scholar] [CrossRef] [Green Version]
- Chung, S.S.M.; Ho, E.C.M.; Lam, K.S.L.; Chung, S.K. Contribution of polyol pathway to diabetes-induced oxidative stress. J. Am. Soc. Nephrol. 2003, 14, S233–S236. [Google Scholar] [CrossRef] [Green Version]
- Sevin, G.; Kerry, Z.; Sozer, N.; Ozsarlak-Sozer, G. Taurine supplementation protects lens against glutathione depletion. Eur. Rev. Med. Pharmacol. Sci. 2021, 25, 4520–4526. [Google Scholar]
- Morre, D.M.; Lenaz, G.; Morre, D.J. Surface oxidase and oxidative stress propagation in aging. J. Exp. Biol. 2000, 203, 1513–1521. [Google Scholar] [CrossRef]
- Funatsu, H.; Yamashita, H.; Noma, H.; Shimizu, E.; Mimura, T.; Hori, S. Prediction of macular edema exacerbation after phacoemulsification in patients with nonproliferative diabetic retinopathy. J. Cataract. Refract. Surg. 2002, 28, 1355–1363. [Google Scholar] [CrossRef]
- Lim, S.A.; Joo, C.K.; Kim, M.S.; Chung, S.K. Expression of p53 and caspase-8 in lens epithelial cells of diabetic cataract. J. Cataract. Refract. Surg. 2002, 28, 1102–1108. [Google Scholar] [CrossRef]
- Xie, Q.; Xue, L.; Cao, X.; Huang, L.; Song, Y. Apoptosis of Lens Epithelial Cells and Expression of NLRP3-related Proteins in Patients with Diabetes and Cataract. Ocul. Immunol. Inflamm. 2022, 16, 1–8. [Google Scholar] [CrossRef]
- Du, S.; Shao, J.; Xie, D.; Zhang, F. Decorin inhibits glucose-induced lens epithelial cell apoptosis via suppressing p22phox-p38 MAPK signaling pathway. PLoS ONE 2020, 15, e0224251. [Google Scholar] [CrossRef]
- Kojima, M.; Sun, L.; Hata, I.; Sakamoto, Y.; Sasaki, H.; Sasaki, K. Efficacy of α-lipoic acid against diabetic cataract in rat. Jpn. J. Ophthalmol. 2007, 51, 10–13. [Google Scholar] [CrossRef]
- Yoshida, M.; Kimura, H.; Kyuki, K.; Ito, M. Combined effect of vitamin E and insulin on cataracts of diabetic rats fed a high cholesterol diet. Biol. Pharm. Bull. 2004, 27, 338–344. [Google Scholar] [CrossRef] [Green Version]
- Zhao, W.; Devamanoharan, P.S.; Henein, M.; Ali, A.H.; Varma, S.D. Diabetes-induced biochemical changes in rat lens: Attenuation of cataractogenesis by pyruvate. Diabetes Obes. Metab. 2000, 2, 165–174. [Google Scholar] [CrossRef] [PubMed]
- Meyer, C.H.; Sekundo, W. Nutritional supplementation to prevent cataract formation. Dev. Ophthalmol. 2005, 38, 103–119. [Google Scholar]
- Yu, L.; Chen, Y.; Tooze, S.A. Autophagy pathway: Cellular and molecular mechanisms. Autophagy 2017, 14, 207–215. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, J.; Ye, W.; Xu, W.; Chang, T.; Zhang, L.; Ma, J.; Per, R.; He, M.; Zhou, J. Activation of autophagy inhibits epithelial to mesenchymal transition process of human lens epithelial cells induced by high glucose conditions. Cell. Signal. 2020, 75, 109768. [Google Scholar] [CrossRef]
- Sun, Y.; Wang, X.; Chen, B.; Huang, M.; Ma, P.; Xiong, L.; Huang, J.; Chen, J.; Huang, S.; Liu, Y. TFEB-Mediated Lysosomal Restoration Alleviates High Glucose-Induced Cataracts Via Attenuating Oxidative Stress. Investig. Ophthalmol. Vis. Sci. 2022, 63, 26. [Google Scholar] [CrossRef]
- Suryanarayana, P.; Saraswat, M.; Mrudula, T.; Krishna, T.P.; Krishnaswamy, K.; Reddy, G.B. Curcumin and Turmeric Delay Streptozotocin-Induced Diabetic Cataract in Rats. Investig. Ophthalmol. Vis. Sci. 2005, 46, 2092. [Google Scholar] [CrossRef]
- Lu, Q.; Yang, T.; Zhang, M.; Du, L.; Liu, L.; Zhang, N.; Guo, H.; Zhang, F.; Hu, G.; Yin, X. Preventative effects of Ginkgo biloba extract (EGb761) on high glucose-cultured opacity of rat lens. Phytother. Res. 2014, 28, 767–773. [Google Scholar] [CrossRef] [PubMed]
- Shu, D.Y.; Lovicu, F.J. Myofibroblast transdifferentiation: The dark force in ocular wound healing and fibrosis. Prog. Retin. Eye Res. 2017, 60, 44–65. [Google Scholar] [CrossRef] [PubMed]
- Saika, S.; Yamanaka, O.; Flanders, K.C.; Okada, Y.; Miyamoto, T.; Sumioka, T.; Shirai, K.; Kitano, A.; Miyazaki, K.; Tanaka, S.-I. Epithelial-mesenchymal transition as a therapeutic target for prevention of ocular tissue fibrosis. Endocr. Metab. Immune Disord.-Drug Targets 2008, 8, 69–76. [Google Scholar] [CrossRef]
- Kalluri, R.; Neilson, E. Epithelial-mesenchymal transition and its implications for fibrosis. J. Clin. Investig. 2003, 112, 1776–1784. [Google Scholar] [CrossRef]
- De Iongh, R.U.; Wederell, E.; Lovicu, F.J.; McAvoy, J.W. Transforming growth factor-beta-induced epithelial-mesenchymal transition in the lens: A model for cataract formation. Cells Tissues Organs 2005, 179, 43–55. [Google Scholar] [CrossRef]
- Shirai, K.; Tanaka, S.I.; Lovicu, F.J.; Saika, S. The murine lens: A model to investigate in vivo epithelial-mesenchymal transition. Dev. Dyn. 2018, 247, 340–345. [Google Scholar] [CrossRef] [PubMed]
- Feng, H.; Zhao, X.; Guo, Q.; Feng, Y.; Ma, M.; Guo, W.; Dong, X.; Deng, C.; Li, C.; Song, X.; et al. Autophagy resists EMT process to maintain retinal pigment epithelium homeostasis. Int. J. Biol. Sci. 2019, 15, 507–521. [Google Scholar] [CrossRef] [Green Version]
- Gugnoni, M.; Sancisi, V.; Manzotti, G.; Gandolfi, G.; Ciarrocchi, A. Autophagy and epithelial-mesenchymal transition: An intricate interplay in cancer. Cell Death Dis. 2016, 7, e2520. [Google Scholar] [CrossRef] [PubMed]
- Cabrerizo, J.; Urcola, J.A.; Vecino, E. Changes in the lipidomic profile of aqueous humor in open-angle glaucoma. Eur. J. Gastroenterol. Hepatol. 2017, 26, 349–355. [Google Scholar] [CrossRef] [PubMed]
- Edwards, G.; Aribindi, K.; Guerra, Y.; Lee, R.K.; Bhattacharya, S.K. Phospholipid profiles of control and glaucomatous human aqueous humor. Biochimie 2014, 101, 232–247. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cabrerizo, J.; Urcola, J.A.; Vecino, E.; Melles, G. Changes in lipidomic profile of aqueous humour in Fuchs endothelial dystrophy. Acta Ophthalmol. 2017, 95, 727–732. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, B.; Cong, C.; Li, Z.; Hao, L.; Yuan, X.; Wang, W.; Shi, Y.; Liu, T. Analysis of the aqueous humor lipid profile in patients with polypoidal choroidal vasculopathy. Exp. Eye Res. 2022, 222, 109160. [Google Scholar] [CrossRef]
- Wang, J.; Zhang, Y.; Li, W.; Zhou, F.; Li, J. Changes in the Lipid Profile of Aqueous Humor From Diabetic Cataract Patients. Transl. Vis. Sci. Technol. 2022, 11, 5. [Google Scholar] [CrossRef]
- Pietrowska, K.; Dmuchowska, D.A.; Krasnicki, P.; Bujalska, A.; Samczuk, P.; Parfieniuk, E.; Kowalczyk, T.; Wojnar, M.; Mariak, Z.; Kretowski, A.; et al. An exploratory LC-MS-based metabolomics study reveals differences in aqueous humor composition between diabetic and non-diabetic patients with cataract. Electrophoresis 2018, 39, 1233–1240. [Google Scholar] [CrossRef]
- Gutierrez, L.; Lim, J.S.; Foo, L.L.; Ng, W.Y.; Yip, M.; Lim, G.Y.S.; Wong, M.H.Y.; Fong, A.; Rosman, M.; Mehta, J.S.; et al. Application of artificial intelligence in cataract management: Current and future directions. Eye Vis. 2022, 9, 3. [Google Scholar] [CrossRef]
- Bathaie, S.Z.; Bahmani, F.; Farajzadeh, A. Handbook of Nutrition, Diet and the Eye; Academic Press: San Diego, CA, USA, 2014; pp. 141–155. [Google Scholar]
- Andrzejewska-Buczko, J.; Pawlak, D.; Tankiewicz, A.; Matys, T.; Buczko, W. Possible involvement of kynurenamines in the pathogenesis of cataract in diabetic patients. Med. Sci. Monit. 2001, 7, 742–745. [Google Scholar]
- Cheng, X.; Song, Z.; Wang, X.; Xu, S.; Dong, L.; Bai, J.; Li, G.; Zhang, C. A Network Pharmacology Study on the Molecular Mechanism of Protocatechualdehyde in the Treatment of Diabetic Cataract. Drug Des. Dev. Ther. 2021, ume 15, 4011–4023. [Google Scholar] [CrossRef]
- Anwar, S.; Khan, S.; Almatroudi, A.; Khan, A.A.; Alsahli, M.A.; Almatroodi, S.A.; Rahmani, A.H. A review on mechanism of inhibition of advanced glycation end products formation by plant derived polyphenolic compounds. Mol. Biol. Rep. 2021, 48, 787–805. [Google Scholar] [CrossRef]
- Wang, J.; Xu, J.; Gong, X.; Yang, M.; Zhang, C.; Li, M. Biosynthesis, Chemistry, and Pharmacology of Polyphenols from Chinese Salvia Species: A Review. Molecules 2019, 24, 155. [Google Scholar] [CrossRef] [Green Version]
- Kim, Y.S.; Kim, N.H.; Lee, S.W.; Lee, Y.M.; Jang, D.S.; Kim, J.S. Effect of protocatechualdehyde on receptor for advanced glycation end products and TGF-beta1 expression in human lens epithelial cells cultured under diabetic conditions and on lens opacity in streptozotocin-diabetic rats. Eur. J. Pharmacol. 2007, 569, 171–179. [Google Scholar] [CrossRef]
- Du, L.; Hao, M.; Li, C.; Wu, W.; Wang, W.; Ma, Z.; Yang, T.; Zhang, N.; Isaac, A.T.; Zhu, X.; et al. Quercetin inhibited epithelial mesenchymal transition in diabetic rats, high-glucose-cultured lens, and SRA01/04 cells through transforming growth factor-β2/phosphoinositide 3-kinase/Akt pathway. Mol. Cell. Endocrinol. 2017, 452, 44–56. [Google Scholar] [CrossRef]
- Qi, M.; Elion, E.A. MAP kinase pathways. J. Cell Sci. 2005, 118 Pt 16, 3569–3572. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Li, F.; Zhang, G.; Kang, L.; Guan, H. Ultraviolet-B induces ERCC6 repression in lens epithelium cells of age-related nuclear cataract through coordinated DNA hypermethylation and histone deacetylation. Clin. Epigenetics 2016, 8, 62. [Google Scholar] [CrossRef] [Green Version]
- Xie, L.; Santhoshkumar, P.; Reneker, L.W.; Sharma, K.K. Histone deacetylase inhibitors trichostatin A and vorinostat inhibit TGFβ2-induced lens epithelial-to-mesenchymal cell transition. Investig. Ophthalmol. Vis. Sci. 2014, 55, 4731–4740. [Google Scholar] [CrossRef] [Green Version]
- Chen, X.; Xiao, W.; Chen, W.; Luo, L.; Ye, S.; Liu, Y. The epigenetic modifier trichostatin A, a histone deacetylase inhibitor, suppresses proliferation and epithelial-mesenchymal transition of lens epithelial cells. Cell Death Dis. 2013, 4, e884. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kanada, F.; Takamura, Y.; Miyake, S.; Kamata, K.; Inami, M.; Inatani, M.; Oki, M. Histone acetyltransferase and Polo-like kinase 3 inhibitors prevent rat galactose-induced cataract. Sci. Rep. 2019, 9, 20085. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Galiniak, S.; Aebisher, D.; Bartusik-Aebisher, D. Health benefits of resveratrol administration. Acta Biochim. Pol. 2019, 66, 13–21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smith, A.J.O.; Eldred, J.A.; Wormstone, I.M. Resveratrol Inhibits Wound Healing and Lens Fibrosis: A Putative Candidate for Posterior Capsule Opacification Prevention. Investig. Ophthalmol. Vis. Sci. 2019, 60, 3863–3877. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Doganay, S.; Borazan, M.; Iraz, M.; Cigremis, Y. The effect of resveratrol in experimental cataract model formed by sodium selenite. Curr. Eye Res. 2006, 31, 147–153. [Google Scholar] [CrossRef]
- Zheng, Y.; Liu, Y.; Ge, J.; Wang, X.; Liu, L.; Bu, Z.; Liu, P. Resveratrol protects human lens epithelial cells against H2O2-induced oxidative stress by increasing catalase, SOD-1, and HO-1 expression. Mol. Vis. 2010, 16, 1467–1474. [Google Scholar]
- Chen, P.; Yao, Z.; He, Z. Resveratrol protects against high glucose-induced oxidative damage in human lens epithelial cells by activating autophagy. Exp. Ther. Med. 2021, 21, 440. [Google Scholar] [CrossRef]
- Chen, M.; Rong, R.; Xia, X. Spotlight on pyroptosis: Role in pathogenesis and therapeutic potential of ocular diseases. J. Neuroinflammation 2022, 19, 183. [Google Scholar] [CrossRef]
- Giugliano, D.; Ceriello, A.; Paolisso, G. Oxidative stress and diabetic vascular complications. Diabetes Care 1996, 19, 257–267. [Google Scholar] [CrossRef]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mrugacz, M.; Pony-Uram, M.; Bryl, A.; Zorena, K. Current Approach to the Pathogenesis of Diabetic Cataracts. Int. J. Mol. Sci. 2023, 24, 6317. https://doi.org/10.3390/ijms24076317
Mrugacz M, Pony-Uram M, Bryl A, Zorena K. Current Approach to the Pathogenesis of Diabetic Cataracts. International Journal of Molecular Sciences. 2023; 24(7):6317. https://doi.org/10.3390/ijms24076317
Chicago/Turabian StyleMrugacz, Małgorzata, Magdalena Pony-Uram, Anna Bryl, and Katarzyna Zorena. 2023. "Current Approach to the Pathogenesis of Diabetic Cataracts" International Journal of Molecular Sciences 24, no. 7: 6317. https://doi.org/10.3390/ijms24076317