
How Single Amino Acid Substitutions Can Disrupt a Protein Hetero-Dimer Interface: Computational and Experimental Studies of the LigAB Dioxygenase from Sphingobium sp. Strain SYK-6

Abstract
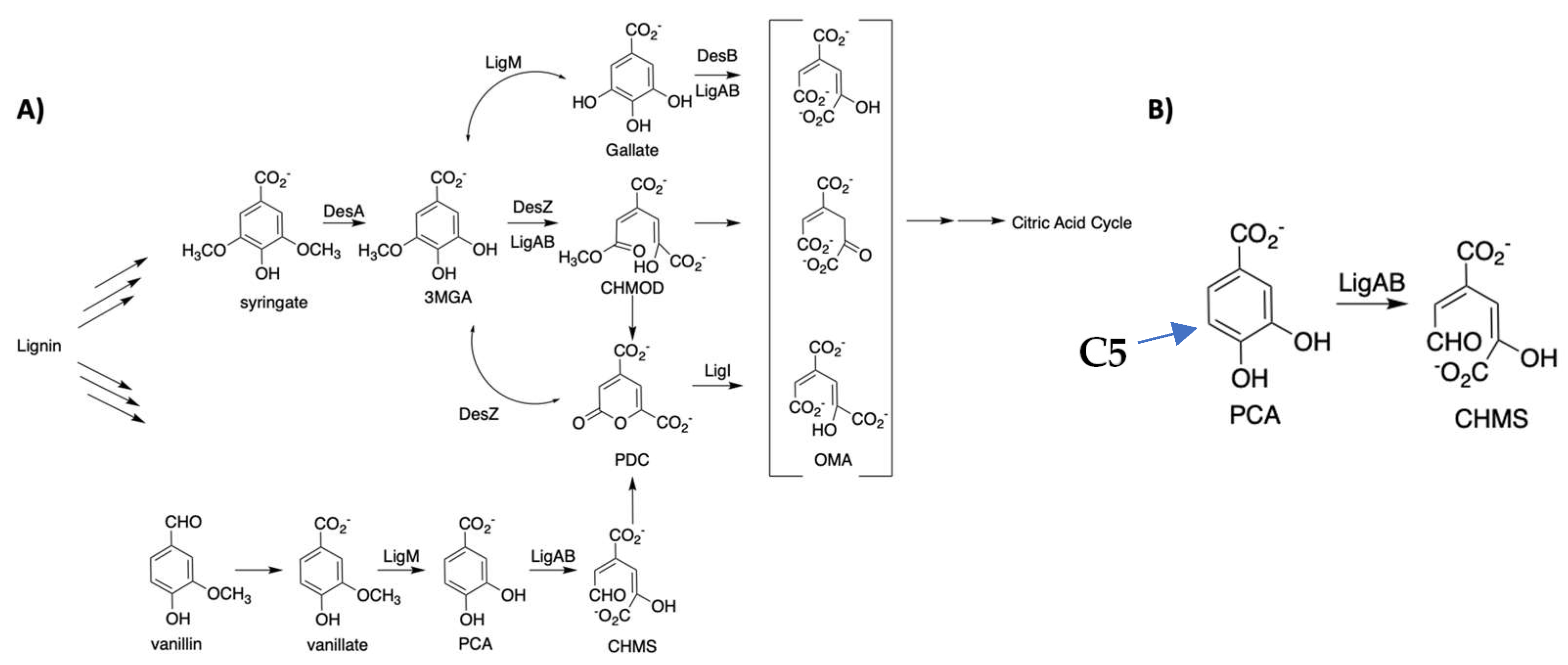
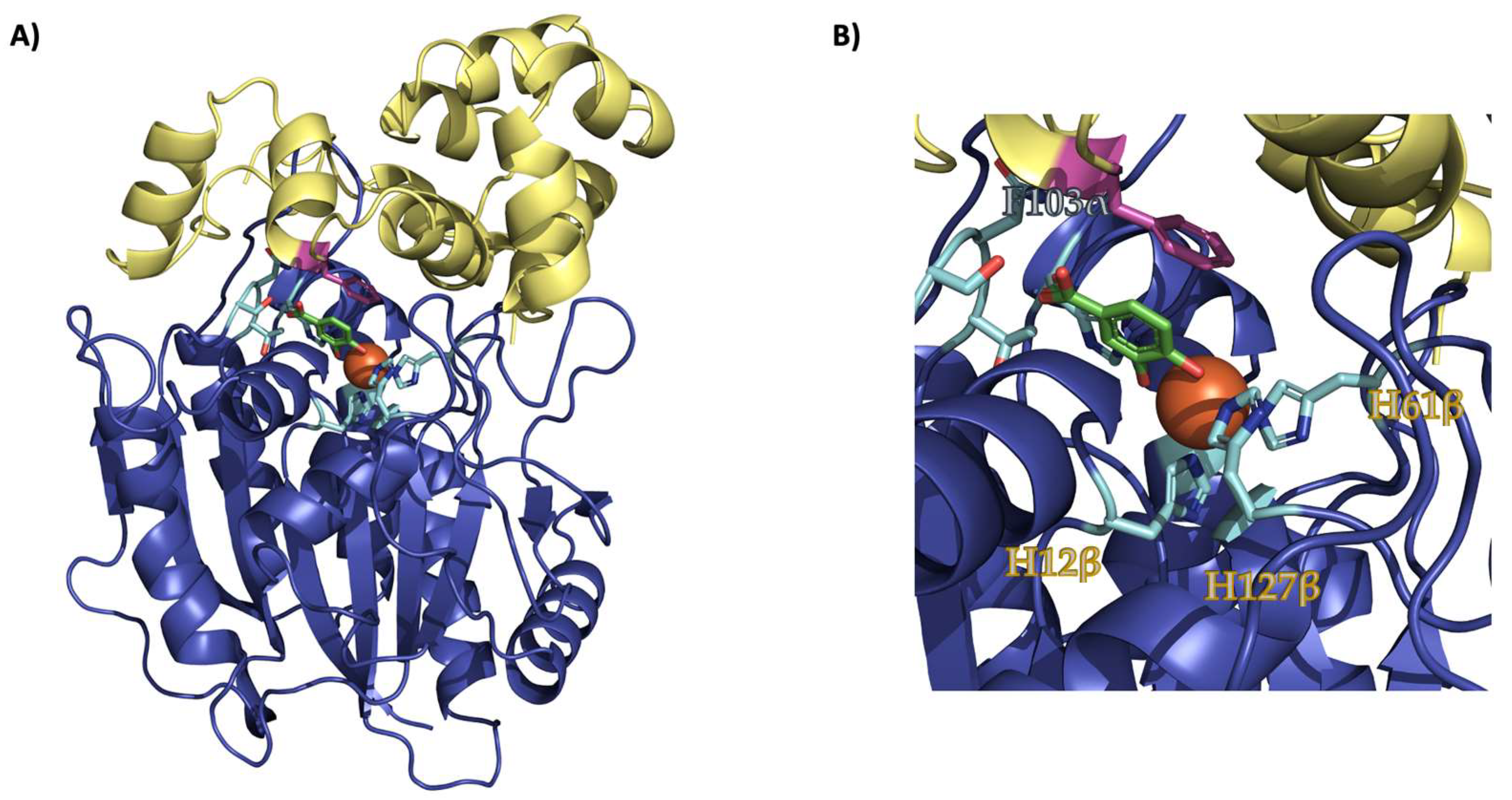
:1. Introduction
2. Results
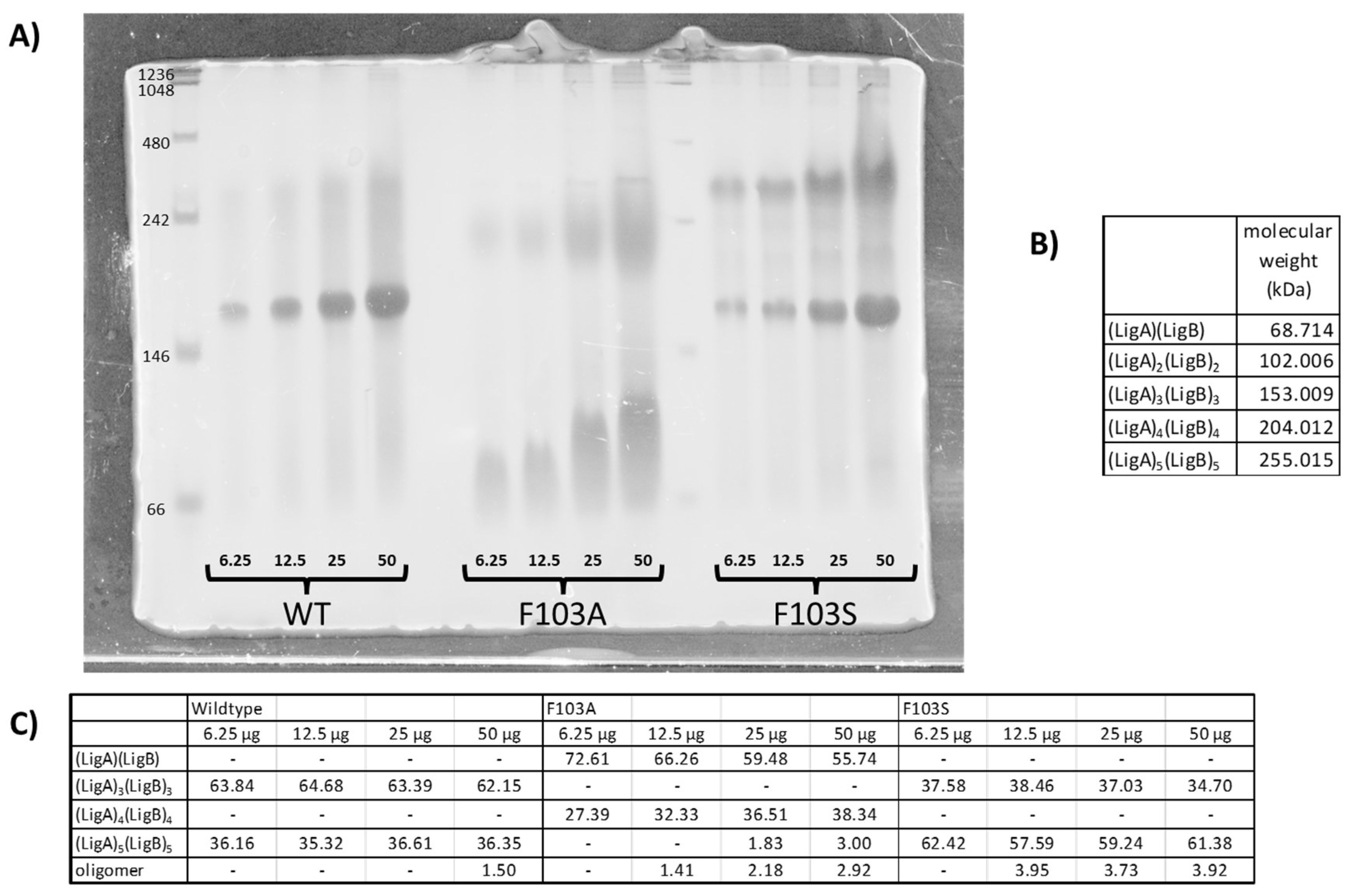
2.1. Protein Dimer Purification and SDS-PAGE Analysis
2.2. Native Gel Analysis
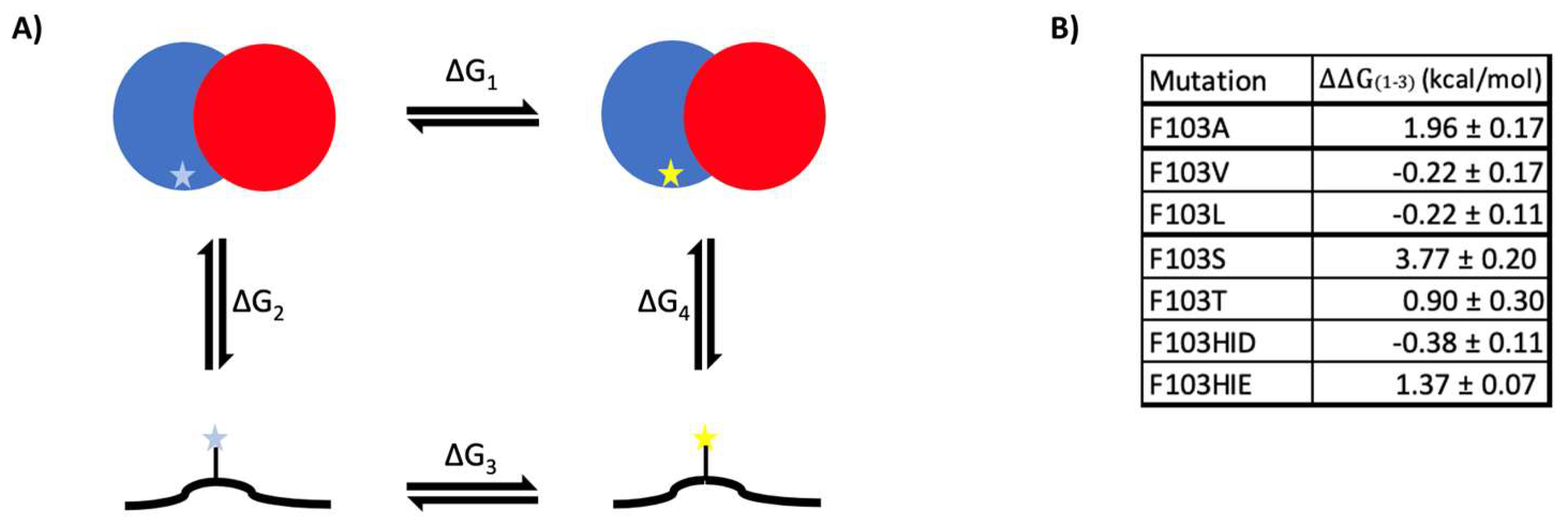
2.3. Heterodimer Stability Calculation
3. Discussion
4. Materials and Methods
4.1. Molecular Dynamic Simulations and Relative Binding Free Energy Calculation
4.2. Protein Expression and Purification
4.3. SDS-PAGE and Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Beauchet, R.; Monteil-Rivera, F.; Lavoie, J.M. Conversion of Lignin to Aromatic-Based Chemicals (L-Chems) and Biofuels (L-Fuels). Bioresour. Technol. 2012, 121, 328–334. [Google Scholar] [CrossRef] [PubMed]
- Bruijnincx, P.C.A.; Rinaldi, R.; Weckhuysen, B.M. Unlocking the Potential of a Sleeping Giant: Lignins as Sustainable Raw Materials for Renewable Fuels, Chemicals and Materials. Green Chem. 2015, 17, 4860–4861. [Google Scholar] [CrossRef]
- Pandey, M.P.; Kim, C.S. Lignin Depolymerization and Conversion: A Review of Thermochemical Methods. Chem. Eng. Technol. 2011, 34, 29–41. [Google Scholar] [CrossRef]
- Bugg, T.D.H.; Ahmad, M.; Hardiman, E.M.; Rahmanpour, R. Pathways for Degradation of Lignin in Bacteria and Fungi. Nat. Prod. Rep. 2011, 28, 1883–1896. [Google Scholar] [CrossRef] [PubMed]
- Bugg, T.D.; Ahmad, M.; Hardiman, E.M.; Singh, R. The Emerging Role for Bacteria in Lignin Degradation and Bio-Product Formation. Curr. Opin. Biotechnol. 2011, 22, 394–400. [Google Scholar] [CrossRef] [PubMed]
- Hammel, K.E.; Cullen, D. Role of Fungal Peroxidases in Biological Ligninolysis. Curr. Opin. Plant Biol. 2008, 11, 349–355. [Google Scholar] [CrossRef] [PubMed]
- Xue, P.; Takashi, E.; Kaoru, H.; Eiji, M.; Seiji, N.; Yoshihiro, K.; Kazuhide, K.; Masao, F. Cloning of a Sphingomonas Paucimobilis SYK-6 Gene Encoding a Novel Oxygenase That Cleaves Lignin-Related Biphenyl and Characterization of the Enzyme. Appl. Environ. Microbiol. 1998, 64, 2520–2527. [Google Scholar] [CrossRef] [Green Version]
- Masai, E.; Katayama, Y.; Nishikawa, S.; Yamasaki, M.; Morohoshi, N.; Haraguchi, T. Detection and Localization of a New Enzyme Catalyzing the β-Aryl Ether Cleavage in the Soil Bacterium (Pseudomonas Paucimobilis SYK-6). FEBS Lett. 1989, 249, 348–352. [Google Scholar] [CrossRef] [Green Version]
- Masai, E.; Katayama, Y.; Nishikawa, S.; Fukuda, M. Characterization of Sphingomonas Paucimobilis SYK-6 Genes Involved in Degradation of Lignin-Related Compounds. J. Ind. Microbiol. Biotechnol. 1999, 23, 364–373. [Google Scholar] [CrossRef]
- Sugimoto, K.; Senda, T.; Aoshima, H.; Masai, E.; Fukuda, M.; Mitsui, Y. Crystal Structure of an Aromatic Ring Opening Dioxygenase LigAB, a Protocatechuate 4,5-Dioxygenase, under Aerobic Conditions. Structure 1999, 7, 953–965. [Google Scholar] [CrossRef] [Green Version]
- Burroughs, A.M.; Glasner, M.E.; Barry, K.P.; Taylor, E.A.; Aravind, L. Oxidative Opening of the Aromatic Ring: Tracing the Natural History of a Large Superfamily of Dioxygenase Domains and Their Relatives. J. Biol. Chem. 2019, 294, 10211–10235. [Google Scholar] [CrossRef]
- MASAI, E.; KATAYAMA, Y.; FUKUDA, M. Genetic and Biochemical Investigations on Bacterial Catabolic Pathways for Lignin-Derived Aromatic Compounds. Biosci. Biotechnol. Biochem. 2007, 71, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Barry, K.P.; Taylor, E.A. Characterizing the Promiscuity of LigAB, a Lignin Catabolite Degrading Extradiol Dioxygenase from Sphingomonas Paucimobilis SYK-6. Biochemistry 2013, 52, 6724–6736. [Google Scholar] [CrossRef] [Green Version]
- Tsagogiannis, E.; Vandera, E.; Primikyri, A.; Asimakoula, S.; Tzakos, A.G.; Gerothanassis, I.P.; Koukkou, A.-I. Characterization of Protocatechuate 4,5-Dioxygenase from Pseudarthrobacter Phenanthrenivorans Sphe3 and In Situ Reaction Monitoring in the NMR Tube. Int. J. Mol. Sci. 2021, 22, 9647. [Google Scholar] [CrossRef] [PubMed]
- Daisuke, K.; Eiji, M.; Keisuke, M.; Yoshihiro, K.; Masao, F. Characterization of the Gallate Dioxygenase Gene: Three Distinct Ring Cleavage Dioxygenases Are Involved in Syringate Degradation by Sphingomonas Paucimobilis SYK-6. J. Bacteriol. 2005, 187, 5067–5074. [Google Scholar] [CrossRef] [Green Version]
- Daisuke, K.; Eiji, M.; Keisuke, M.; Yoshihiro, K.; Masao, F. Characterization of the 3-O-Methylgallate Dioxygenase Gene and Evidence of Multiple 3-O-Methylgallate Catabolic Pathways in Sphingomonas Paucimobilis SYK-6. J. Bacteriol. 2004, 186, 4951–4959. [Google Scholar] [CrossRef] [Green Version]
- Barry, K.P.; Ngu, A.; Cohn, E.F.; Cote, J.M.; Burroughs, A.M.; Gerbino, J.P.; Taylor, E.A. Exploring Allosteric Activation of LigAB from Sphingobium sp. Strain SYK-6 through Kinetics, Mutagenesis and Computational Studies. Arch. Biochem. Biophys. 2015, 567, 35–45. [Google Scholar] [CrossRef] [Green Version]
- Barry, K.P.; Cohn, E.F.; Ngu, A.; Taylor, E.A. Improving Alternate Lignin Catabolite Utilization of LigAB from Sphingobium sp. Strain SYK-6 through Site Directed Mutagenesis. Process Biochem. 2015, 50, 1634–1639. [Google Scholar] [CrossRef] [Green Version]
- Fahrenkamp, D.; Li, J.; Ernst, S.; Schmitz-Van de Leur, H.; Chatain, N.; Küster, A.; Koschmieder, S.; Lüscher, B.; Rossetti, G.; Müller-Newen, G. Intramolecular Hydrophobic Interactions Are Critical Mediators of STAT5 Dimerization. Sci. Rep. 2016, 6, 35454. [Google Scholar] [CrossRef] [Green Version]
- Porter, C.J.; Wilce, M.C.J.; Mackay, J.P.; Leedman, P.; Wilce, J.A. Grb7-SH2 Domain Dimerisation Is Affected by a Single Point Mutation. Eur. Biophys. J. 2005, 34, 454–460. [Google Scholar] [CrossRef]
- Yan, C.; Wu, F.; Jernigan, R.L.; Dobbs, D.; Honavar, V. Characterization of Protein–Protein Interfaces. Protein J. 2008, 27, 59–70. [Google Scholar] [CrossRef] [Green Version]
- Prakash, A.; Janosi, L.; Doxastakis, M. GxxxG Motifs, Phenylalanine, and Cholesterol Guide the Self-Association of Transmembrane Domains of ErbB2 Receptors. Biophys. J. 2011, 101, 1949–1958. [Google Scholar] [CrossRef] [Green Version]
- Unterreitmeier, S.; Fuchs, A.; Schäffler, T.; Heym, R.G.; Frishman, D.; Langosch, D. Phenylalanine Promotes Interaction of Transmembrane Domains via GxxxG Motifs. J. Mol. Biol. 2007, 374, 705–718. [Google Scholar] [CrossRef] [PubMed]
- Vaillancourt, F.H.; Bolin, J.T.; Eltis, L.D. The Ins and Outs of Ring-Cleaving Dioxygenases. Crit. Rev. Biochem. Mol. Biol. 2006, 41, 241–267. [Google Scholar] [CrossRef] [PubMed]
- Abraham, M.J.; Murtola, T.; Schulz, R.; Páll, S.; Smith, J.C.; Hess, B.; Lindahl, E. GROMACS: High Performance Molecular Simulations through Multi-Level Parallelism from Laptops to Supercomputers. SoftwareX 2015, 1, 19–25. [Google Scholar] [CrossRef] [Green Version]
- Hornak, V.; Abel, R.; Okur, A.; Strockbine, B.; Roitberg, A.; Simmerling, C. Comparison of Multiple Amber Force Fields and Development of Improved Protein Backbone Parameters. Proteins Struct. Funct. Bioinform. 2006, 65, 712–725. [Google Scholar] [CrossRef] [Green Version]
- Jacobson, M.P.; Pincus, D.L.; Rapp, C.S.; Day, T.J.F.; Honig, B.; Shaw, D.E.; Friesner, R.A. A Hierarchical Approach to All-Atom Protein Loop Prediction. Proteins Struct. Funct. Bioinform. 2004, 55, 351–367. [Google Scholar] [CrossRef] [Green Version]
- Jacobson, M.P.; Friesner, R.A.; Xiang, Z.; Honig, B. On the Role of the Crystal Environment in Determining Protein Side-Chain Conformations. J. Mol. Biol. 2002, 320, 597–608. [Google Scholar] [CrossRef]
- Søndergaard, C.R.; Olsson, M.H.M.; Rostkowski, M.; Jensen, J.H. Improved Treatment of Ligands and Coupling Effects in Empirical Calculation and Rationalization of PKa Values. J. Chem. Theory Comput. 2011, 7, 2284–2295. [Google Scholar] [CrossRef]
- Olsson, M.H.M.; Søndergaard, C.R.; Rostkowski, M.; Jensen, J.H. PROPKA3: Consistent Treatment of Internal and Surface Residues in Empirical PKa Predictions. J. Chem. Theory Comput. 2011, 7, 525–537. [Google Scholar] [CrossRef]
- Gapsys, V.; Michielssens, S.; Seeliger, D.; de Groot, B.L. Pmx: Automated Protein Structure and Topology Generation for Alchemical Perturbations. J. Comput. Chem. 2015, 36, 348–354. [Google Scholar] [CrossRef] [Green Version]
- Seeliger, D.; de Groot, B.L. Protein Thermostability Calculations Using Alchemical Free Energy Simulations. Biophys J 2010, 98, 2309–2316. [Google Scholar] [CrossRef] [Green Version]
- Gapsys, V.; Seeliger, D.; de Groot, B.L. New Soft-Core Potential Function for Molecular Dynamics Based Alchemical Free Energy Calculations. J. Chem. Theory Comput. 2012, 8, 2373–2382. [Google Scholar] [CrossRef] [PubMed]
- Shirts, M.R.; Bair, E.; Hooker, G.; Pande, V.S. Equilibrium Free Energies from Nonequilibrium Measurements Using Maximum-Likelihood Methods. Phys. Rev. Lett. 2003, 91, 140601. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Crooks, G.E. Nonequilibrium Measurements of Free Energy Differences for Microscopically Reversible Markovian Systems. J. Stat. Phys. 1998, 90, 1481–1487. [Google Scholar] [CrossRef]
- Hess, B.; Bekker, H.; Berendsen, H.J.C.; Fraaije, J.G.E.M. LINCS: A Linear Constraint Solver for Molecular Simulations. J. Comput. Chem. 1997, 18, 1463–1472. [Google Scholar] [CrossRef]
- Eisenhaber, F.; Lijnzaad, P.; Argos, P.; Sander, C.; Scharf, M. The Double Cubic Lattice Method: Efficient Approaches to Numerical Integration of Surface Area and Volume and to Dot Surface Contouring of Molecular Assemblies. J. Comput. Chem. 1995, 16, 273–284. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| α | β | α:β | β:α | |
|---|---|---|---|---|
| Wild-type | 81.1 | 75.3 | 1.077 | 0.928 |
| F103A | 46.8 | 57.3 | 0.817 | 1.224 |
| F103V | 56.7 | 71.8 | 0.790 | 1.266 |
| F103L | 51.7 | 58.8 | 0.879 | 1.137 |
| F103S | 69.0 | 66.2 | 1.042 | 0.959 |
| F103T | 80.7 | 49.1 | 1.644 | 0.608 |
| F103H | 71.1 | 32.4 | 2.194 | 0.456 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rafalowski, A.; Hassan, B.A.; Lou, K.; Nguyen, M.C.; Taylor, E.A. How Single Amino Acid Substitutions Can Disrupt a Protein Hetero-Dimer Interface: Computational and Experimental Studies of the LigAB Dioxygenase from Sphingobium sp. Strain SYK-6. Int. J. Mol. Sci. 2023, 24, 6319. https://doi.org/10.3390/ijms24076319
Rafalowski A, Hassan BA, Lou K, Nguyen MC, Taylor EA. How Single Amino Acid Substitutions Can Disrupt a Protein Hetero-Dimer Interface: Computational and Experimental Studies of the LigAB Dioxygenase from Sphingobium sp. Strain SYK-6. International Journal of Molecular Sciences. 2023; 24(7):6319. https://doi.org/10.3390/ijms24076319
Chicago/Turabian StyleRafalowski, Angelika, Bakar A. Hassan, Kate Lou, Minh Chau Nguyen, and Erika A. Taylor. 2023. "How Single Amino Acid Substitutions Can Disrupt a Protein Hetero-Dimer Interface: Computational and Experimental Studies of the LigAB Dioxygenase from Sphingobium sp. Strain SYK-6" International Journal of Molecular Sciences 24, no. 7: 6319. https://doi.org/10.3390/ijms24076319